Bacterial pathogens of concern within the context of the worldwide emergence and spread of antimicrobial resistance (AMR) are classified under the group ESKAPEE, comprising
Enterococcus faecium,
Staphylococcus aureus,
Acinetobacter baumannii,
Klebsiella pneumoniae,
Pseudomonas aeruginosa,
Enterobacter species, and
Escherichia coli [
1]. These bacterial species represent some of the extremely drug-resistant strains that threaten the relevance of existing antimicrobial therapy due to their current ability to resist all available antimicrobials [
2]. ESKAPEE pathogens are the major cause of healthcare-associated infections (nosocomial), and it is estimated that more than 80% of global deaths are due to ESKAPEE pathogens [
3].
2. Antimicrobial Resistance Mechanisms
Antimicrobial agents are purportedly meant to eliminate microorganisms, particularly those deleterious to the health of plants, animals, and humans. Besides these, antimicrobials are also useful in preserving foods from spoilage. Antimicrobial agents could be of chemical or natural origin. The antibiotic penicillin is recognized as the first microbe-derived antimicrobial compound employed clinically to treat infectious diseases. The three-decade period following the discovery of penicillin is termed the “golden period of antimicrobial therapy”, as numerous antibiotics were discovered and inducted into clinical use [
23]. However, along with the discovery of antimicrobials, bacteria that survived and grew in antibiotics were also discovered, suggesting that such bacteria can be present in the environment or possibly evolve during antibiotic exposure. Penicillin-resistant staphylococci were isolated from clinical samples in the early 1940s, and with the widespread use of antibiotics in the following two decades, the emergence and spread of antibiotic-resistant strains increased dramatically in clinical settings [
24]. The development of resistance followed the discovery of new antimicrobials, and this phenomenon is often aided by the development of cross-resistance to closely related antibiotics or antibiotics belonging to the same group.
The bacterial mechanisms of antibiotic resistance are diverse and are broadly classified into (i) enzymatic inactivation of antibiotics, (ii) modification of drug target, (iii) deceased drug permeability, and (iv) active efflux of antibiotics [
25]. The enzymatic inactivation is achieved by hydrolysis of antibiotics by bacterial enzymes such as the β-lactamases that degrade β-lactam antibiotics such as the penicillins, cephalosporins, and carbapenems, macrolide esterases, and the fosfomycin-inactivating enzymes. The second mechanism of enzymatic inactivation of antibiotics involves the structural modification of drugs by group transfer activities from enzymes such as acetyltransferases, phosphotransferases, glycosyltransferases, and nucleotidyltransferases [
26,
27]. The group transfer mechanism results in the irreversible modification of the antibiotic structure, which can no longer bind to its target. The antibiotics susceptible to this resistance mechanism include chloramphenicol, aminoglycosides, macrolides, and fosfomycin [
23].
Alternatively, the antibiotic targets may be modified to circumvent binding by the antibiotics. This resistance may be achieved by mutations in the genes encoding the proteins which act as antibiotic targets, such as the penicillin-binding proteins (PBPs), the modification of which results in the inability of β-lactams to bind to PBPs resulting in resistance development [
28]. Mutations in QRDRs (quinolone resistance determinant regions) result in the modification of ribosomal targets leading to the development of resistance to quinolone–fluoroquinolone antibiotics [
29].
The outer membrane porins often regulate the entry of antimicrobials into the bacterial cell, the structural modification of which can result in reduced permeability of drugs through the cellular membrane. Such porin modifications result from mutations in porin-encoding genes under antibiotic pressure, and bacteria expressing defective porin proteins resist the entry of antibiotics such as the aminoglycosides, chloramphenicol, tetracyclines, β-lactams, and fluoroquinolones [
30].
The efflux-mediated antimicrobial resistance mechanism involves the transmembrane proteins, which transport structurally diverse substrates across the membrane and primarily function to extrude toxic metabolites, Kreb’s cycle intermediates such as salts, sugars, vitamins, fatty acids, and amino acids, among others [
9,
31]. Based on the source of energy that drives this efflux process, the transporter proteins are broadly grouped into primary active transporters, which use ATP, and secondary active transporters, which utilize the concentration gradient established across the membrane by the primary active transport and respiration [
32,
33].
3. Antimicrobial Transporter Superfamilies
Many of the transport systems used by microorganisms can fall into one of a variety of protein superfamilies, the vast majority of which have been incorporated into a massive transporter classification database (TCDB) [
34]. These and other biological transport systems have been systematically and taxonomically organized and are readily accessible in the TCDB, a continually updated resource [
35]. The transporter superfamilies are established according to similarities in evolutionary origin, sequence, protein structure, and energization modes [
36,
37].
Solute transporters that involve antimicrobial agents and bacterial pathogens can be grouped into a handful of superfamilies. The drug/metabolite transporter (DMT) superfamily consists of many protein families, such as the small multidrug resistance (SMR) family [
38]. The multidrug/oligosaccharidyl-lipid/polysaccharide (MOP) superfamily harbors [
39] several distantly related protein families, one of which is the multidrug and toxic compound extrusion transporters (MATE) family consisting of numerous cation-based multidrug antiporters [
40]. The resistance–nodulation–cell division (RND) superfamily contains many solute transporters, many of which are found in bacterial pathogens conferring resistance to multiple antimicrobial agents [
41]. The proteobacterial antimicrobial compound efflux (PACE) family encompasses members that are multidrug efflux pump systems in bacteria [
42]. One of the largest groups is the major facilitator superfamily (MFS), consisting of secondary active transporters and passive facilitators [
43].
3.1. Major Facilitator Superfamily (MFS)
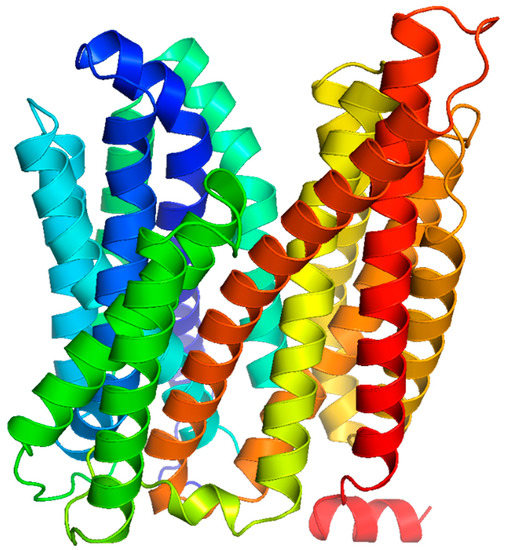
The MFS solute transporters consist of passive and secondary active transporters such as uniporters, symporters, and antiporters [
45]. The protein members of the MFS typically have between 300 and 600 amino acids with 12 or 14 transmembrane domains composed of α-helices [
46]. For instance, the LmrS multidrug efflux pump from
S. aureus is predicted to harbor 14 membrane-spanning helices, with the N- and C-termini facing the cytoplasmic side of the cell membrane,
Figure 2.
Figure 2. Predicted structure of LmrS from S. aureus. The LmrS multidrug efflux pump of the MFS has 14 predicted transmembrane domains that are α-helical in nature.
MFS Antimicrobial Efflux Pump Structure and Function
The first transporter of the MFS to be characterized physiologically and at the molecular level was the
E. coli tetracycline efflux pump, TetA, discovered in the laboratory of Levy [
49,
50]. Shortly afterward, related tetracycline transporters were found in Gram-negative and -positive microorganisms, collectively called TetA, with sub-classes including A-D and others [
51]. Interestingly, it was discovered that these so-called single-substrate TetA transporters were homologous to bacterial multidrug efflux pumps, such as NorA [
52], MdfA [
53], and QacA [
54], which are among the now most intensively studied antimicrobial transporters of the MFS.
High-resolution protein structures have been elucidated for several multidrug efflux pumps of the MFS. Multidrug efflux pumps of the MFS that have been crystalized and structures determined include EmrD [
55], YajR [
56,
57], MdfA [
53,
58], and SotB [
59], all from
E. coli, and more recently, NorC from
S. aureus [
60] and LmrP from
Lactococcus lactis [
61]. These MFS drug transport proteins generally have two global domains, the N-terminus and the C-terminus domains, each harboring six-helical bundles [
62] related by a so-called twofold pseudosymmetrical axis that runs perpendicular to the membrane bilayer [
63]. This structural motif, called the MFS fold, appears in many multidrug efflux pump proteins and symporters of the superfamily [
64,
65].
Thus far, the MFS antimicrobial and multidrug efflux pumps share a highly conserved signature sequence called motif C [
68] and the antiporter motif [
69,
70] in the fifth transmembrane helix. The antiporter motif C has the consensus amino acid sequence of “G (X)
8 G (X)
3 G P (X)
2 G G” [
71] consisting largely of a highly hydrophobic region with a proline manifested as a so-called GP dipeptide, where X is any amino acid [
69,
70]. The residues of the antiporter motif appear to play major mechanistic roles during drug and multidrug transport [
48]. The first evidence to demonstrate the functional importance of the antiporter motif was reported on TetA(C), in which the highly conserved residue Gly-147, part of the GP dipeptide, was necessary for tetracycline resistance [
70].
4. Efflux Pump Inhibition
Multidrug efflux pumps of the MFS are known to confound treatment efforts toward clinical bacterial infections owing to their abilities to extrude many clinically relevant antibiotics [
23]. Thus, understanding the modulation of antimicrobial agent efflux by MFS drug pumps in pathogenic microorganisms is of considerable interest to restore the clinical efficacy of treatment of infections caused by multidrug-resistant pathogens [
44,
94]. Naturally occurring plant-derived agents as resistance modulators in such cases represent a promising avenue [
31].
The NorA multidrug efflux pump system of
S. aureus has been a good target for transport inhibition studies reviewed elsewhere [
98]. In
S. aureus, plant-derived chalcones showed synergistic activity with antimicrobial agents ciprofloxacin, norfloxacin, and ethidium bromide involving the MFS proteins NorA and MepA [
99]. Molecular docking analysis of these chalcones showed close interactions with NorA residues Ser-337, Met-338, Gly-339, and Asn-340, implicating these residues as good molecular targets for transport studies and resistance modulation [
99].
Interestingly, evaluation of the 1,8-naphthyridine sulfonamides showed that they could inhibit both β-lactamases and the QacC and QacA/B efflux pumps in
S. aureus [
101]. Thus, these agents and their derivatives show promise in addressing infection by pathogens by targeting more than one resistance mechanism, as in the case of resistances conferred by enzymatic inactivation and active drug efflux. More recently, berberine, a naturally occurring plant compound and known efflux pump inhibitor of Mdr1p, an MFS multidrug efflux pump from
Candida albicans [
102], reduced the resistance to multiple antimicrobial agents conferred by MdfA of
E. coli [
103]. Using a combination of molecular simulation dynamics and physiological analysis of transport by MdfA across the membrane, berberine was reported to affect the formation of salt bridges and alter the hydrophobic interactions of MdfA with water in the membrane during the transport cycle to inhibit transport [
103].
5. Antimicrobial Resistance and Biofilms
Dynamics between Biofilm Formation and Antibiotic Resistance
Bacteria form biofilms in most scenarios where the dispersal and planktonic stages are considered intermediate or transitional. Biofilms are reported from the sea, rivers, food processing surfaces, medical implants, and the International Space Station [
141]. Biofilms are medically important since almost 65–80% of human chronic infections are attributed to pathogenic biofilms [
142]. The biofilm has become a menace in the food processing industry due to its persistence on food contact surfaces [
143]. Biofilms are a survival strategy for bacteria to escape environmental stress, including predation by the bacteriophage [
144]. Environmental stress triggers the transition of free-swimming planktonic forms to sessile forms that attach to a biotic or abiotic surface [
145].
There are various aspects in the relationship between antibiotic resistance and biofilm formation. One aspect is that the biofilm acts as the antibiotic resistance gene pool, thus facilitating the emergence of antibiotic resistance bacteria. Another is that the biofilms enhance the survivability of drug-resistant bacteria in a harsh environment, thus helping them to sustain in various environments for a long time.
A statistically negative correlation was observed between biofilm formation and MDR in
Acinetobacter baumanii. The MDR and XDR isolates formed weak biofilms compared to non-MDR isolates producing robust biofilms [
148]. Exposure to sub-inhibitory and sub-lethal concentrations of different antimicrobials triggers biofilm formation in different bacteria. A study conducted with
P. aeruginosa found that exposure to aminoglycosides, particularly tobramycin, had the most effect on biofilm formation. Neither polymyxin B, a peptide antibiotic cationic like the aminoglycosides, nor carbenicillin or chloramphenicol had any effect on biofilm formation. Thus,
P. aeruginosa forms biofilm as a specific response to the aminoglycoside antibiotics [
149].
In contrast, tolerance to an antimicrobial agent is the ability of a microorganism to survive but neither grow nor die in the presence of a bactericidal antimicrobial agent measured as minimum bactericidal concentration (MBC) [
151]. The reduced susceptibility of biofilm to antibiotics could be due to complex interactions, and hence a clear demarcation as antibiotic resistance or tolerance might not be feasible in all the settings. Hence many authors use the term recalcitrance to denote the reduced susceptibility of the biofilm community to the antibiotics tested [
152,
153,
154]. The mechanisms involved in antibiotic recalcitrance in a biofilm are depicted in
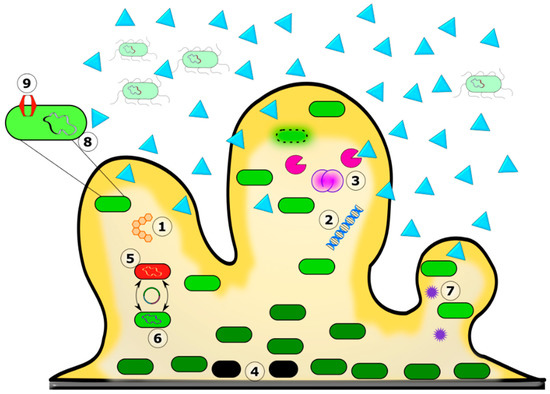
Figure 3.
Figure 3. Mechanism of antibiotic recalcitrance in the biofilm matrix. The biofilm’s color gradient (yellow) depicts the availability of nutrients and oxygen from high to low and darker to lighter shades. The color differentiation in the bacterial cells depicts their physiological state, with the lighter ones in active phases and the darker green ones in less active stationary phases. Biofilm greatly reduced the diffusion of certain antibiotic molecules (blue triangles). 1. The exopolysaccharide. 2. eDNA. 3. Bacterial autolysis releases antibiotic-binding and degrading molecules into the matrix. 4. Persister cells. 5. Mixed species in the biofilm. 6. Horizontal gene transfer and increased frequency of mutation. 7. Quorum sensing. 8. Biofilm-specific antibiotic resistance genes. 9. Efflux pump.
6. Role of the Biofilm Matrix in Antibiotic Recalcitrance
The biofilm matrix comprises components such as the extracellular polymeric substance (EPS), extracellular DNA (eDNA), proteins, and lipids inside which the bacterial cells are embedded. In most biofilms, the bacterial cells account for less than 10% of the biofilm’s dry mass [
155]. The biofilm composition differs between species and even within the species. The MRSA biofilms are more proteinaceous compared to the polysaccharide-rich MSSA biofilms [
156].
7. MFS Transporters and Biofilms
Efflux pumps play important roles in the formation of biofilms, as well as in the antibiotic resistance of biofilm bacteria. The overexpression of tetracycline resistance efflux pump TetA(C) in
Escherichia coli biofilm contributes to forming mature biofilms, stress tolerance, and antimicrobial resistance [
157]. The inactivation of efflux pumps abolishes biofilm formation, suggesting that efflux pumps as essential for biofilm formation and its persistence [
158]. Pinostrobin, a plant-derived flavonoid compound, has anti-efflux and anti-biofilm activities. This compound supposedly interacts more efficiently with MFS efflux pumps of Gram-positive bacteria, reducing the MIC of ciprofloxacin by 128 times in MRSA [
159].
Since biofilm formation is strongly associated with the quorum-sensing mechanism of bacteria, efflux pumps promote biofilm formation by reducing the impact of antibacterials and extrusion of quorum-sensing signaling molecules [
161,
162]. Efflux pump genes are overexpressed in biofilms, and this corresponds with the overexpression of quorum-sensing genes, suggesting a strong relationship between these two processes [
163,
164,
165]. Biologically derived polyamines such as cadaverine, putrescine, spermidine, and spermine are important in bacteria for oxidative stress tolerance, biofilm formation, and persistence. These molecules are also substrates for efflux pumps, such as the AmvA protein of
Acinetobacter baumannii, suggesting that the biofilm formation and efflux pump activities can be interrelated [
166].
The efflux-pump-mediated biofilm formation is a complex process involving an interplay between multiple pathways, and the net effect of these interactions might vary in different bacterial species. For example, the inactivation of LmrB in
Streptococcus mutans resulted in increased EPS secretion and biofilm formation while upregulating other efflux genes’ expression [
167]. In many Gram-negative and -positive bacteria, efflux pumps contribute positively to biofilm formation [
168]. Efflux pump inhibitors (EPI), in many instances, not only enhance the susceptibility of bacteria to antimicrobials but also reduce or inhibit biofilm formation [
169]
The quorum-sensing system also regulates the expression of virulence genes, suggesting that antibiotic resistance, biofilm formation, virulence, and persistence of pathogenic bacteria stay interlinked and provide opportunities to identify the means of interfering with these processes with the ultimate goal of pathogen control [
171]. Apart from clinical implications, efflux pump inhibition and the consequent quorum-sensing inhibition could positively impact the shelf life of highly perishable products such as fish.
In view of growing number of antibiotics being detected as substrates for MFS efflux pumps and the realization of the importance of these pumps in multidrug resistance in human pathogens, modulation of efflux pump using natural, semisynthetic or synthetic compounds has gained significance. Efflux pump inhibition can restore the efficacy of antibiotics, prevent development of resistance to antibiotics due to mutations in target genes and can also have significant negative effects on the survival and persistence of pathogens.