1. Introduction
Cellular disease modeling is a powerful tool used to study the underlying mechanisms of various diseases and to develop new treatments. These models can be used to study both genetic and acquired diseases, including cancer, neurodegeneration, and infectious diseases [
1,
2,
3,
4].
One of the key benefits of cellular disease modeling is that it allows the study of diseases in a controlled environment in ways that would not be possible in a living organism, such as introducing specific genetic mutations or environmental factors to study their effects on the cells. Additionally, cellular disease modeling can also be used to study complex infectious diseases that involve multiple pathways and interactions between different cell types. By studying the effects of different viruses or bacteria on the cells, it is possible uncover the underlying mechanisms of the disease and to test the effectiveness of new treatments, providing insights into the disease process [
5,
6,
7].
The ability to obtain an embryonic-like pluripotent state from differentiated murine fibroblasts using transcription factors changed the paradigm of the stem cell biology field [
8]. One of the most commonly used models for studying cellular diseases involves the use of induced pluripotent stem cells (iPSCs). Human iPSCs can be obtained from different somatic tissue cell types and can be differentiated into any specialized cell type in the human body through specific factors that control cell fate [
9,
10]. Additionally, iPSCs can also be used to study genetic diseases, particularly those with unfathomable genetic backgrounds, which can provide insights into the underlying mechanisms of the disease [
11] or when the translation of animal models is limited [
12,
13,
14,
15,
16]. Furthermore, they can serve as a relevant model for studying host species and genotype-specific parasite interactions [
17,
18,
19,
20].
Organoids technology can recapitulate the complex cellular interactions and microenvironment of native tissues, enabling the study of disease-specific mechanisms in a more physiologically relevant context than traditional 2D cell culture or animal models. For instance, organoids derived from intestinal stem cells can be used to investigate inflammatory bowel disease, while those derived from neural stem cells can be used to study neurodegenerative disorders. Furthermore, organoids can be utilized to study the effects of genetic variations or environmental factors on organ development and function, providing insights into the mechanisms that underlie human diseases [
21,
22,
23].
2. iPSCs
The discovery of iPSCs from mouse ESC was first reported in 2006 by a team of scientists led by Shinya Yamanaka. They demonstrated that by introducing four specific transcription factors, octamer-binding transcription factor 3 or 4 (Oct3/4), sex-determining region Y-box 2 (SOX2), myc pronto-oncogene (c-Myc), and Kruppel-like factor 4 (Klf4), through retrovirus-mediated transfection, they could reprogram adult cells into a pluripotent state. In 2007, the same team generated and cultured iPSCs from adult human fibroblasts using the same retroviral transduction factors, which exhibited similar characteristics and differentiation potential in vitro and teratomas to hESC [
9]. Regarding the reprogramming process, iPSCs exhibit similar results in morphology, proliferation, gene expression, and teratomas compared to embryonic stem cells, avoiding the ethical concerns related with the use of embryonic stem cells [
8,
24,
25,
26]. This breakthrough was a significant step forward in the field of stem cell research, as it provided a way to generate patient-specific pluripotent cells without the need for embryos, which had been a major ethical concern in the field [
26,
27].
Since the discovery of iPSCs, there have been many advancements in the technology used to generate and manipulate these cells. The use of viral vectors to introduce the reprogramming factors was replaced by non-viral methods, such as the use of plasmids or proteins, which eliminated the risk of insertional mutagenesis. Additionally, the number of reprogramming factors required were reduced, making the process more efficient and cost-effective [
28,
29]. Some of the advantages of using iPSCs include:
-
Ethical concerns: iPSCs can be derived from adult cells, avoiding the ethical concerns associated with embryonic stem cell research [
30];
-
Patient-specific cells: iPSCs can be generated from a patient’s own cells, allowing for the creation of patient-specific cells for use in therapy. This can help to avoid immune rejection of the transplanted cells [
31];
-
Disease modeling: iPSCs can be used to create models of specific diseases, which can aid in the understanding of the disease and the development of new treatments [
18];
-
Drug development: iPSCs can be used to test the safety and efficacy of new drugs on a variety of cell types, including those that are difficult to obtain from living donors [
32,
33];
-
Tissue repair and regeneration: iPSCs have the potential to be used to repair or regenerate damaged or diseased tissue, such as in the treatment of heart disease, diabetes, and neurodegenerative disorders [
15];
-
Cost-effective: iPSCs can be created from a small sample of adult cells; however, can be very cost-effective compare to other methods [
34,
35,
36].
The potential applications of iPSCs are vast, and ongoing research is being conducted in several areas. One of the most promising fields is regenerative medicine, where iPSCs can be used to generate specific cell types for tissue engineering and cell replacement therapy. For example, iPSCs can be differentiated into heart muscle cells, which can then be used to repair damaged heart tissue in patients with heart disease [
37,
38,
39]. Similarly, iPSCs can be differentiated into insulin-producing cells, which can be used to treat patients with diabetes [
40,
41].
iPSCs differentiated into hepatocyte-like cells can be a good substitute for primary human hepatocytes because they can be maintained in long-term culture. Moreover, generating cells from patients with specific genetic backgrounds to study genotype-phenotype relationships may in the future avoid the need for organ donation [
42,
43].
Proof-of-concept studies revealed the efficacy of these cells in drug development, high-throughput drug screening, and modeling liver diseases and viral and parasitic infections, such as malaria [
44,
45].
Another area of research for iPSCs is drug discovery and development. Through the generation of iPSCs from patients with specific diseases, in vitro models of these diseases can be created and used to study the underlying causes of the disease and to test the efficacy of new drugs. This can greatly accelerate the drug development process and ultimately lead to new treatments for patients [
46,
47].
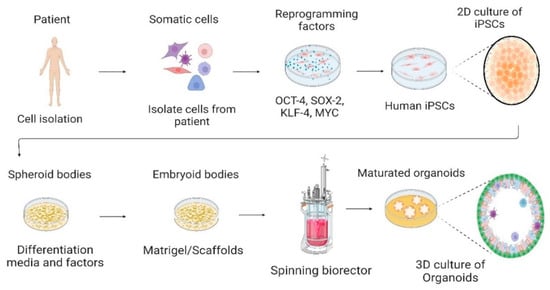
iPSCs also have potential applications in personalized medicine, with iPSCs generated from a patient’s own cells; researchers can create patient-specific cells that can be used in cell-based therapies. Additionally, iPSCs can be used to generate patient-specific organoids, which are miniaturized versions of organs that can be used to study the development and progression of diseases [
48,
49] (
Figure 1).
Figure 1. Culture of human-induced pluripotent stem cells in the 2D culture system and differentiation into 3D organoids. Somatic cells are isolated from a human patient, cultured in vitro, and transduced by pluripotency transcription factors, such as OCT-4, SOX-2, KLF-4, and c-MYC. Through reprogramming that induces a pluripotent state in somatic cells, induced pluripotent stem cells (iPSCs) are generated. Cells are cultured and differentiated under specific factors and matrices to generate organoids. Created with BioRender.com, accessed on 23 July 2022.
iPSCs technology was extensively studied for neurological disorders in recent years due to the limitations of studying the human brain. iPSCs can be directly generated from patients with neurological diseases, allowing the study of nervous system diseases in vitro using 2D, 3D, and BBB models [
50,
51,
52,
53]. The iPSCs differentiation and their use as in vitro model of neurodegenerative disorders, such as Parkinson’s disease (PD), enables a better understanding of neuronal cell death and contributes to the development of drug discovery [
47,
54]. iPSC derived from PD and differentiated into dopaminergic neurons revealed disease-related cell phenotypes, such as impaired mitochondrial function, increased oxidative stress, increased apoptosis, and an accumulation of α-synuclein, providing more insights into PD pathophysiology [
51,
55].
The iPSC lines derived from Alzheimer’s disease (AD) patients, a neurodegenerative disease characterized by β-amyloid (Aβ) plaques and neurofibrillary tangles with progressive cognitive decline, showed neurons differentiated with disease characteristics. A study demonstrated an increased Aβ42:40 percentage of generated cortical neurons from AD patients-iPSC lines compared with control neurons of non-AD iPSCs and different signatures for Aβ43, Aβ38, Aβ fragments in 2D and 3D cultures [
56]. Hence, iPSCs from patient-derived neural cells can reflect the human genetics and physiology of AD and other dementia diseases, being an excellent cellular model to explore Alzheimer’s pathogenesis and lead to neuronal drug discovery [
57,
58].
However, there are also some ethical concerns surrounding the use of iPSCs. One concern is that the use of iPSCs in research and therapy could perpetuate inequalities in access to healthcare, as the technology to generate and manipulate these cells is currently expensive and not widely available [
59]. Moreover, there are concerns about the limitation of genetic diversity, which jeopardizes equity and can impede the acceleration of biological discoveries [
60]. There is also potential for the use of iPSCs in controversial areas, such as human cloning or the creation of genetically modified organisms [
61,
62].
The iPSCs technology also has several technical and scientific drawbacks that must be overcome for its full potential to be realized. One major technical weakness of iPSC technology is the variability in the quality and characteristics of iPSCs generated from different sources. iPSCs can vary in their differentiation potential, genomic stability, epigenetic modifications, and gene expression patterns. These variations can be influenced by the methods of reprogramming, the quality of the starting cells, and the culture conditions used to maintain the iPSCs. This variability can make it difficult to compare results between studies and to predict the behavior of iPSC-derived cells in a clinical setting [
63,
64,
65,
66].
The efficiency of the iPSCs reprogramming process can be low, especially for certain cell types or patient populations. Additionally, the reprogramming process can be time-consuming and expensive, requiring the use of multiple factors and genetic modifications. Scientifically, one of the major drawbacks of iPSC technology is the potential for genetic abnormalities to be introduced during the reprogramming process. Reprogramming can lead to changes in gene expression, DNA methylation patterns, and chromosomal stability, which can alter the properties and behavior of iPSCs and their differentiated derivatives. These changes can also increase the risk of tumor formation or other adverse effects when iPSC-derived cells are used in a clinical context [
67,
68,
69].
Furthermore, iPSC technology faces challenges in differentiating into specific cell types with high efficiency and fidelity. The protocols for differentiation can be complex and require optimization for each cell type, and there can be significant variability in the quality and purity of the differentiated cells produced. Additionally, there may be functional differences between iPSC-derived cells and their natural counterparts, which may limit their therapeutic potential [
70,
71,
72,
73,
74].
In conclusion, due to the plasticity of iPSCs in differentiation into several cell types and their self-renewal capacity, they constitute a vital tool for research in regenerative medicine, neurological disease modeling, cell therapies, and drug screening [
12,
47,
54,
57,
75,
76,
77,
78,
79,
80,
81,
82]. However, iPSC technology holds technical and scientific hurdles that must be overcome to fully realize its potential. Continued research and development in iPSC technology will be critical to address these challenges and to enable the safe and effective use of iPSC-derived cells in clinical applications, as well as to address the ethical concerns surrounding their use [
30].
3. Cell Culture System—From 2D to 3D
Cell culture is the process of growing cells in a controlled environment outside of their natural setting; the two main types of cell culture systems are 2D and 3D [
83].
The 2D cell culture system refers to the growth of cells on a flat surface, such as a Petri dish, microscope slide, or a culture plate. This is the most common and well-established type of cell culture, and it is widely used in basic research, drug discovery, and biotechnology [
84]. In a 2D cell culture system, cells are grown in monolayers and are typically maintained in a liquid medium, such as nutrient broth or serum-containing medium, which provides the necessary nutrients and growth factors for cell growth and proliferation [
84,
85,
86,
87].
An important application of 2D culture is the production of biological products. Many of these products, such as monoclonal antibodies and recombinant proteins, are produced through cells that were grown in 2D culture. These cells can be genetically engineered to produce specific proteins, which are then harvested and purified to create the final product [
88,
89].
The usage of iPSCs in 2D culture can help better understand neurotransmission, the central nervous system, and the differentiation of neurons, astrocytes, and microglia [
90], giving more insights into the genetic and molecular conditions of neurological disorders [
76].
Advantages of 2D cell culture include its simplicity and ease of use, as well as its accessibility and low cost, due to the easy set up and maintenance, the cells can be easily observed and manipulated. Additionally, 2D cell culture is well-suited for growing a wide range of cell types, including primary cells, stem cells, and cancer cells and allows the differentiation of specific subtypes of cells in a dish [
91,
92,
93].
Although 2D cell culture is widely used and valuable tool in laboratory research, it has a limited role in the disease modeling because it cannot accurately resemble the dynamic complexity of the in vivo environment, cell–cell communications, and tissue- and organ-level structures. This is because 2D cultures can only do so for differentiation of one cell type in a mono-culture system [
76,
90,
94,
95,
96,
97], leading to artificial behavior of cells, such as altered proliferation and differentiation, and reduced cellular interaction compared to in vivo conditions [
98,
99]. For example, drug development has a high-cost when the process goes from target identification stage to clinical use. The drug screening performed in cells cultured in 2D is not representative of cells in a tissue microenvironment, resulting in a high failure rate in drug discovery and low levels of approved drugs in the market [
100].
Despite these limitations, 2D cell culture remains a valuable tool in the laboratory, and recent advances improved the system, such as techniques for analyzing and characterizing cells, such as high-content imaging and transcriptomics.
The use of 3D cell culture systems started more than a decade ago and allow more complexity between cells’ interaction with heterotypic settings, mimicking multiple structures, using scaffold-based or scaffold-free cultures [
76,
101]. This system present similarities with tissue architecture in vivo, respecting the phenotypic and functional characteristics and circumventing a limitation of monolayer cultures [
102,
103].
Despite these limitations, 2D cell culture remains a valuable tool in the laboratory, and many advances were made in recent years to improve the system, such as techniques for analyzing and characterizing cells, such as high-content imaging and transcriptomics. These techniques allow for a more detailed understanding of the behavior and characteristics of cells in culture [
104,
105].
Scaffolds-based techniques are typically made of synthetic or natural materials, such as hydrogel-based supports, which provide a physical structure for cells to attach and grow on. Scaffold-free technique can be used to create small structures, such as microchannels and microwells, that can control cell behavior, such as suspended microplates or spheroids with coated ultra-low attachment microplates. Hydrogels are hydrophilic polymeric materials that can mimic the extracellular matrix (ECM), such as fibrinogen, hyaluronic acid, collagen, matrigel, or gelatin, providing a supportive environment for cells. They allow soluble factors such as cytokines and growth factors to move through the tissue-like gel [
87,
100].
The 3D cell culture system also allows for the study of cell–cell interactions, cell-matrix interactions, and cell-microenvironment interactions that closely mimics the in vivo microenvironment [
51,
106]. Moreover, 3D cell culture enables the formation of complex cellular structures, such as spheroids involved in bone regeneration [
107] and organoids. These features provide insights about the organ’s behavior and bridge the gap between 2D cell culture and animal models [
87].
Organoids and spheroids are 3D structures that are increasingly used in biological and medical research, as they better mimic the in vivo environment compared to traditional 2D cell cultures. Although organoids and spheroids have some similarities, they have distinct differences that distinguish them from one another [
108].
Organoids are 3D structures derived from stem cells or tissue explants that can self-organize into structures resembling specific organs. They exhibit complex cellular organization, spatial orientation, and function similar to that of the in vivo tissue, composed of multiple cell types, and can be used to study organ development, disease modeling, and drug screening. They are typically cultured in a specialized medium containing growth factors that promote differentiation and tissue-specific gene expression [
108,
109,
110,
111]. Spheroids, on the other hand, are aggregates of cells that form a 3D structure, but lack the complex organization and functional specialization of organoids, and often have a homogenous cell population. They can be generated by culturing cells in non-adherent conditions, such as suspension cultures, hanging drops, or microfluidic devices. Spheroids can be composed of a single cell type or multiple cell types and can be used to study cell–cell interactions, drug screening, and tumor biology [
112,
113,
114,
115].
Other 3D structures that can be generated in vitro include organotypic cultures, tissue-engineered constructs, and scaffolds. Organotypic cultures are 3D cultures that resemble the in vivo tissue architecture but lack the self-organization seen in organoids. Tissue-engineered constructs are artificial structures that are created using cells and biomaterials and can be used for tissue engineering applications. Scaffolds are 3D structures that provide support for cells to grow and differentiate, and can be used for tissue engineering, drug screening, and regenerative medicine applications [
116,
117,
118,
119,
120]. In summary, while organoids, spheroids, and other 3D structures have some similarities, they can be distinguished by their cellular organization, complexity, and functional specialization. Organoids are self-organized structures that resemble specific organs, while spheroids are simple aggregates of cells lacking distinct organization. Other 3D structures, such as organotypic cultures, tissue-engineered constructs, and scaffolds, are useful for a range of applications, including tissue engineering and drug screening.
Additionally, the 3D cell culture has the potential to provide different cell types, giving helpful information about tumor cell biology, signal transduction, cell migration, drug discovery, angiogenesis, metabolic profiling, inflammation, and apoptosis. Moreover, the 3D culture model exhibits cellular behavior that is more similar to in vivo in responses to oxidative stress compared to the 2D model [
121]. For example, the 3D culture allowed us to understand how the formative components of the cerebellar structure interact and how they self-organize and differentiate into Purkinje cells. The addition of the factors lead to a polarized design with a rhombic-lip-like layout and a three-layered cytoarchitecture similar to the human embryonic cerebellum, as in the first trimester [
122]. The 3D model applied in neural cell culture of AD pathogenesis study showed relevant information about amyloid-β (Aβ) aggregation and increased concentration of hyperphosphorylated tau [
123,
124].
The 3D model could be used for pre-clinical studies at a low cost, such as for the gastrointestinal tract–liver system [
125]. It is a potential new model for pre-clinical stage research, providing relevant information about other types of diseases, and could contribute to the use of fewer animal models [
87,
125,
126]. Moreover, it is a model that can be used to study drug efficacy and toxicity because it can provide a more realistic model for drug testing, allowing for a better understanding of how drugs interact with cells. However, there are also some limitations associated with 3D cell culture, such as the complexity of the system and the difficulty of monitoring the cells within it. With the growing interest in 3D cell culture, it is expected that this field will continue to develop and provide new opportunities for research and drug development in the future [
87,
100].
In conclusion, 2D and 3D cell culture systems are both important tools in cell biology research and biotechnology, but they have distinct advantages and limitations. The 2D cell culture system is well-established, simple, and accessible, while 3D cell culture more closely mimics the in vivo environment of cells, making it useful for studying certain types of biological processes. Both 2D and 3D cell culture are important for understanding the biology of cells and for developing new therapies [
99,
127].
4. Organoids Technology
Organoids are 3D structures, miniaturized versions of organs that are grown in a lab using stem cells or other cell types. These structures closely mimic the architecture, function, and behavior of their in vivo counterparts, making them valuable tools for studying a wide range of biological processes and diseases [
111,
128].
Organoids can form endodermal, mesodermal, and ectodermal organs. Organs derived from cells of the endoderm layer are usually associated with complex systems, including the gastrointestinal and respiratory tract, and all their associated organs. Mesoderm derivatives are associated with blood vessels, muscles, kidneys, heart, bone, cartilage, and reproductive organs. Those derived from the ectoderm, from the cells of the epithelium layers, are related to the nervous system, such as the brain [
22]. Additionally, organoids can be established in different ways, through the extracellular matrix, such as collagen or matrigel, and with differentiating factors, in plates, or rotating bioreactors, which are usually used for cerebral organoids as they allow greater medium perfusion. Air-liquid interface (ALI) can also generate organoids, such as microfluidic, in which the top layer of cells is exposed to air, and the basal layer is in contact with the culture medium, usually applied to renal, gastrointestinal, and neural organoids [
22,
129,
130,
131].
Organoids have gained particular focus as an appealing model since they are 3D self-organized structures and the microenvironment is preserved, including morphological and biological issues of organs. They can mimic genetic diseases, host-infectious disease interaction, regenerative therapy, or drug screening using patient-specific iPSCs cells and establishing biobanks. Furthermore, through molecular technologies, such as the lentiviral expression system and CRISPR/CAS9, it is possible to manipulate the genome of organoids, allowing for disease replication and targeted gene therapy [
23,
76,
132,
133].
Organoids can be used to study human biology and disease in a way that is not possible with traditional cell culture or animal models. For example, organoids can be derived from patient-specific stem cells, allowing researchers to study the effects of genetic mutations on organ development and function in a way that is not feasible in vivo. Organoids have been used to study a wide range of organs and systems, including the brain, liver, pancreas, and gut. For instance, brain organoids were used to study the development of neural circuits and the effects of genetic mutations on brain function [
134,
135,
136,
137].
Organoids are a potential tool to study organ transplantation and the process of transplant rejection, as well as to test new drugs and other treatments that could be used to prevent rejection. Understanding genetic mutations associated with organ-specific diseases, as well as how genetic mutations affect organ development and function, are important factors that can be explored [
138,
139,
140].
Organoids are a powerful tool for studying human biology and disease, as they allow for the study of the effects of genetic mutations and environmental factors on organ development and function, which is not possible with traditional cell culture or animal models. In addition, organoids can be used to study the efficacy and toxicity of different drug treatments, providing a more physiologically relevant context for drug screening or pathways, providing insights into the mechanisms of drug action. As the technology continues to improve, organoids are likely to play an increasingly important role in scientific research and the development of new treatments for a wide range of diseases. Moreover, organoids have the potential to revolutionize the way of study human diseases and develop new therapeutics, but further research is needed to fully realize their potential. With the continued growth of organoid technology, significant advances are expected in the fields of disease modeling, drug discovery, and personalized medicine [
141] (
Figure 2).
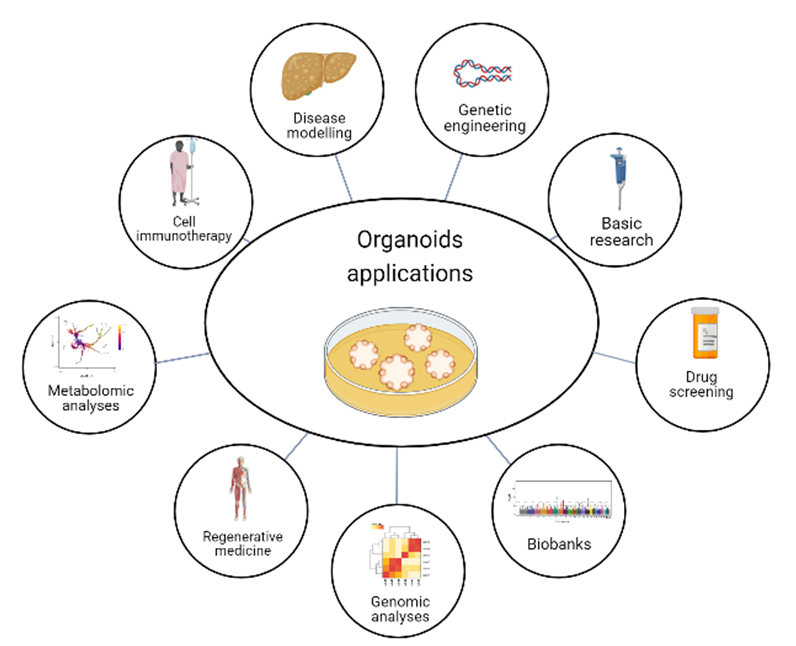
Figure 2. Organoid applications. Organoids have the potential for several purposes, such as basic research, drug screening, genomic and metabolomic analysis, and gene editing to explore disease-linked alleles, called genetic engineering. Moreover, organoids can model diseases, have applications in cell immunotherapy and regenerative medicine, and provide the opportunity to have biobanks. Created with BioRender.com, accessed on 23 July 2022.
This entry is adapted from the peer-reviewed paper 10.3390/cells12060930