Chromatin remodelling is a major mechanism by which cells control fundamental processes including gene expression, the DNA damage response (DDR) and ensuring the genomic plasticity required by stem cells to enable differentiation. The post-translational modification of histone H2B resulting in addition of a single ubiquitin, in humans at lysine 120 (K120; H2Bub1) and in yeast at K123, has key roles in transcriptional elongation associated with the RNA polymerase II-associated factor 1 complex (PAF1C) and in the DDR. H2Bub1 itself has been described as having tumour suppressive roles and a number of cancer-related proteins and/or complexes are recognised as part of the H2Bub1 interactome. These include the RING finger E3 ubiquitin ligases RNF20, RNF40 and BRCA1, the guardian of the genome p53, the PAF1C member CDC73, subunits of the switch/sucrose non-fermenting (SWI/SNF) chromatin remodelling complex and histone methyltransferase complexes DOT1L and COMPASS, as well as multiple deubiquitinases including USP22 and USP44. While globally depleted in many primary human malignancies, including breast, lung and colorectal cancer, H2Bub1 is selectively enriched at the coding region of certain highly expressed genes, including at p53 target genes in response to DNA damage, functioning to exercise transcriptional control of these loci.
- histone monoubiquitination
- E3 ubiquitin ligases
- RNF20
- p53
- BRCA1
- CDC73
- SWI/SNF
- transcriptional elongation
- DNA damage
1. Introduction
Post-translational modifications (PTMs) of core histone proteins H2A, H2B, H3 and H4 constituting the nucleosome have driving roles in modulating the chromatin landscape in order to regulate fundamental processes such as transcription and the DNA damage response (DDR). Examples of histone PTMs include methylation, acetylation, phosphorylation, SUMOylation, proline isomerisation and ubiquitination [1,2]. The addition of a single 76 amino acid (8.5 kDa) ubiquitin, monoubiquitination, is one of the bulkier histone modifications [2,3]. A number of histone lysines (K) are monoubiquitinated in the mammalian genome, including K13, K15, K119, K127 and K129 of histone H2A; K34, K120 and K125 of histone H2B; and K31 on histone H4 (reviewed in [4]). The two most well-known mammalian histone monoubiquitination events occur on K119 of histone H2A (H2AK119ub1) linked to the Polycomb Repressor Complex 1 (PRC1) [5] and K120 of histone H2B (H2Bub1), the latter being the subject of this review. Interestingly, these events have opposing functions, with H2AK119ub1 associated with gene silencing and H2Bub1 frequently associated with active transcription [4–6].
A cascade of enzymatic events is responsible for monoubiquitination, involving an activating ATP-dependent ubiquitin enzyme E1 of which humans have a few, a ubiquitin conjugating enzyme (E2) of which humans have around 40, and a ubiquitin protein ligase (E3) of which humans have as many as 600–1000 [4,7,8]. The most prevalent of the E3 ubiquitin ligases are RING (really interesting new gene) finger domain ligases of which the E3 complex RNF20/RNF40 is the main writer enzyme complex responsible for catalysing H2Bub1 [1,9]. The H2Bub1 chromatin mark is erased by deubiquitinases (DUBs) from the ubiquitin-specific protease (USP) sub-family of DUBs, including USP44 and USP22 (reviewed in [1,10–12]) (Figure 1).
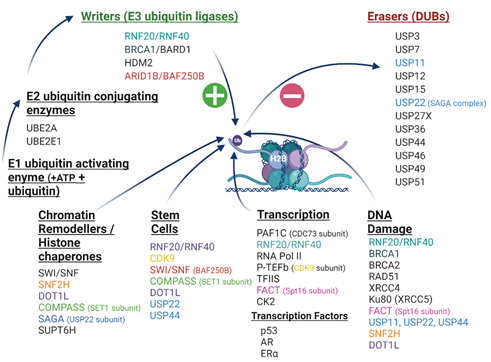
Figure 1. The H2Bub1 interactome. H2Bub1 can be written (green plus symbol) by multiple writer enzymes (E3 ubiquitin ligases) following an enzymatic ubiquitin cascade commencing with an E1 ubiquitin activating enzyme followed by an E2 ubiquitin conjugating enzyme. It can be removed (red minus symbol) by numerous erasing deubiquitinases (DUBs) from the Ubiquitin-Specific Protease (USP) sub-family. H2Bub1 interacts with a number of chromatin remodelling complexes and histone chaperones. Numerous components of the H2Bub1 interactome function in more than one cellular process, such as cyclin-dependent kinase 9 (CDK9) in stem cell plasticity and transcription, identified in this schematic by matching colours. The H2Bub1 interactome continues to be elucidated and is growing as new discoveries are made. Androgen receptor (AR); AT-Rich Interaction Domain 1B (ARID1B); BRG1/BRM-associated factor (BAF250B); BRCA1-Associated RING Domain 1 (BARD1); Breast Cancer Type 1 (BRCA1); Breast Cancer Type 2 (BRCA2); cyclin-dependent kinase 9 (CDK9); estrogen receptor alpha (ERα); complex of proteins associated with SET1 (COMPASS); disrupter of telomere silencing 1-like (DOT1L); facilitates chromatin transcription (FACT); Human Double Minute 2 (HDM2); X-Ray Repair Cross Complementing 5 (Ku80, also known as XRCC5); RNA polymerase II-associated factor 1 complex (PAF1C); Positive Transcription Elongation Factor-b (P-TEFb); tumour protein 53 (p53); RNA polymerase II (RNA Pol II); RAD51 Recombinase (RAD51); RING Finger Protein 20 (RNF20); RING Finger Protein 40 (RNF40); Spt-Ada-Gcn5-acetyltransferase (SAGA); Su(var) 3–9 (SET1, suppressor of position effect variegation), enhancer of zeste (E(z)), and trithorax (Trx); SNF2 homologue, (SNF2H, also known as SMARCA5); Suppressor of Ty Homologue-6 (SUPT6H); Switch/sucrose non-fermenting (SWI/SNF); transcription elongation factor II S (TFIIS); ubiquitin conjugating enzyme E2 A (UBE2A); ubiquitin-conjugating enzyme E2 E (UBE2E1); Ubiquitin-Specific Protease (USP); X-Ray Repair Cross Complementing 4 (XRCC4). Created with BioRender.com.
The enzymatic cascade that writes H2Bub1 is invoked to enable association of the human RNA polymerase II-associated factor 1 (PAF1) transcriptional complex with RNA polymerase II (RNA pol II) to facilitate transcriptional elongation [13]. It is also required to facilitate the accumulation of H2Bub1 at sites of double strand breaks (DSBs) where it functions as a scaffold or docking platform for the recruitment of DNA repair factors [14]. H2Bub1 has also been linked to DNA replication in yeast [15] and in both yeast and human cells, genomic stability through the maintenance of centromeric chromatin [16]. While H2Bub1 itself has been described as having tumour suppressive properties, a number of tumour suppressor proteins are also associated with H2Bub1-related processes including RNF20, CDC73, BRCA1, p53 and members of the switch/sucrose non-fermenting (SWI/SNF) chromatin remodelling complexes (also known as the BAF complexes) [17–20]
Global levels of H2Bub1 and/or its E3 ligase complex members RNF20 and RNF40, have been investigated in numerous tumours using methods including immunohistochemistry and for RNF40 and/or RNF20, assessment of transcript levels and promoter hypermethylation. Dependent upon tumour type, depleted global H2Bub1 levels have been shown to correlate with inflammatory processes that may function as precursor events to tumorigenesis, in tumour progression and in some tumour types have been correlated with a worse prognosis [21,22].
On the background of global loss of H2Bub1 in cancer cells, H2Bub1 is enriched at the coding regions of highly expressed genes in response to DNA damage, including p53 gene targets and genes involved in resistance to therapeutic drugs [23,24]. H2Bub1 therefore has a role in determining the fate of cancer cells that may be amenable to therapeutic intervention. Further evidence for a key role in determining cell fate is the role of H2Bub1 in the differentiation of stem cells, whether it be through engaging with other histone PTMs via histone cross-talk, or the enzymes involved in writing and erasing H2Bub1 [10,12]. H2Bub1 and its associated factors have also been linked to regulation of key development pathways in plants, including seed dormancy/germination and flowering time, as well as resistance to pathogen invasion [25–28]. It is clear that H2Bub1 has a significant role across species to regulate important functions through the modulation of gene expression.
In this review we discuss the role of H2Bub1 in fundamental cellular processes including gene transcription, the DDR and stem cell differentiation. We bring together the current literature on the involvement of H2Bub1 across multiple different tumour types, including links recently established between H2Bub1 and inflammation. Lastly, we investigate the current state-of-play between H2Bub1 and members of its interactome complicit in the establishment and/or progression of malignancy.
2. H2Bub1 and Transcriptional Elongation
Monoubiquitination isn’t the only histone PTM that occurs at K120 of histone H2B. Acetylation at this site (H2BK120ac) written by the lysine acetyl transferase KAT3 is thought to act as a mark of chromatin poised to enter the active state, highlighting the temporal acetylation/ubiquitination switch working to achieve transcription [29]. H2Bub1 physically disrupts chromatin compaction, creating a more open conformation accessible to transcription factors and other proteins and/or protein complexes involved in activities such as DNA repair [30]. This is more than purely physical disruption, as replacement of ubiquitin with the even bulkier molecule small ubiquitin-like modifier (SUMO), does not result in the same functional effects [31]. K120 of histone H2B is physically located at the interface of adjacent nucleosomes, making it possible that H2Bub1 has an impact on nucleosome stacking that may affect nucleosome stability [30]. Chandrasekharan and colleagues demonstrated that nucleosome stability increases in the presence of H2Bub1 during transcription and that this negatively impacts upon cell growth [31], with a later study suggesting that this may be a modest effect [32].
H2Bub1 is enriched at the coding regions of highly expressed genes [24]. In response to stimuli such as a hormone and/or a developmental signal or DNA damaging agent, cyclin-dependent kinase 9 (CDK9), part of the Positive Transcription Elongation Factor-b (P-TEFb) [33] complex, phosphorylates both the H2Bub1 E2 enzyme UBE2A and Ser2 in the carboxy-terminal domain (CTD) of RNA Pol II [12,34]. This creates a binding domain for WAC (WW Domain-Containing Adapter Protein With Coiled-Coil) that directly links the main H2Bub1 E3 ligase complex RNF20/RNF40 to RNA pol II [12,35]. In this co-transcriptional role with H2Bub1, CDK9 has been shown in yeast to be responsible for releasing RNA pol II from promoter-proximal pausing, making it an important regulator of gene expression [1,36,37]. Notably, RNA pol II promoter-proximal pausing has been implicated in the suppression of transcription of antisense genes [38].
Both RNF20 and RNF40 have been shown to physically interact with subunits of the human PAF1C (including the PAF1 subunit itself, CDC73 and CTR9) that associates with RNA Pol II, setting the stage for interaction of all these factors that enable transcriptional elongation to proceed [39] (reviewed in [1,10,12]) (Figure 1, Figure 2A). Enrichment of H2Bub1 occupancy at highly expressed genes has been shown to correlate with the recruitment of Pol II, prior to the increase of mRNA expression and to decrease once RNA Pol II has dissociated [24]. Examples of genes with enriched H2Bub1 include those that are transcriptionally driven by hormone receptors, specifically estrogen receptor alpha (ERα) [22,40,41] important in some breast and ovarian cancers, and the androgen receptor (AR), important in prostate cancer [42], as well as p53 target genes that function in the cellular response to DNA damage [10,24] (Figure 2B).
To enable transcriptional elongation, H2Bub1 recruits the histone chaperone that facilitates chromatin transcription (FACT) that works to remove the H2A-H2B dimer from the nucleosome, therefore removing the physical block to RNA pol II (Figure 2A) [43]. A FACT subunit, Spt16, functions with H2Bub1 to then enable nucleosome reassembly following RNA pol II transcriptional elongation [44]. These apparently opposing functions of H2Bub1 highlight its key roles in the kinetics of nucleosome assembly and disassembly required during and post transcription.
H2Bub1 is also central in trans-histone crosstalk, recruiting histone methyltransferase complexes that further direct chromatin configuration and gene expression [45–48]. The catalytic activity of the disrupter of telomere silencing 1-like (DOT1L) histone methyltransferase that facilitates methylation of histone H3 at K79 (H3K79me) at the proximal region of actively transcribed genes is stimulated by H2Bub1 [49–54]. Loss of H3K79me2 has been correlated with genomic instability [55]. Another active chromatin mark is histone H3K4 methylation that is catalysed by SET1, the methyltransferase subunit of the complex of proteins associated with SET1 (COMPASS) after first being recruited by H2Bub1 [39,56,57]. Furthermore, Basnet and colleagues report an association between the phosphorylation of tyrosine 57 on histone H2A by casein kinase 2 (CK2) and H2Bub1 in yeast and mammalian cells. Loss of this histone H2A phosphorylation event, either by mutation at the histone H2A tyrosine site or inhibition of CK2, leads to loss of H2Bub1, H3K4me3 and H3K79me3, impacting on transcriptional elongation and likely involving the Spt-AdaGcn5 acetyltransferase (SAGA) chromatin modifying complex [58]. This is another example of how H2Bub1 functions in histone crosstalk. These patterns of trans-histone cross-talk cement the importance of H2Bub1 as a central regulator of transcription (Figure 1).
While being a clear enabler of transcriptional elongation, in certain circumstances H2Bub1 can also impede it. At least in yeast, there would appear to be a positional effect determined by gene location given that H2Bub1 at promoters of genes that aren’t expressed appear to inhibit the formation of transcriptional complexes [59]. The H2Bub1 writer enzyme RNF20 has been demonstrated to obstruct binding of the transcription elongation factor transcription elongation factor II S (TFIIS) to the PAF1C, so blocking the ability to relieve stalled RNA pol II on chromatin [60]. In this way, RNF20 works to selectively inhibit the transcription of pro-oncogenic genes located in condensed chromatin, supportive of a tumour suppressor function for this E3 ligase. H2Bub1 occupancy is, therefore, complex and likely tissue, genomic position and contextually specific, as not all genes require H2Bub1 enrichment for their expression [61]. One study combining RNA sequencing and Assay for Transposase Accessible Chromatin (ATAC-seq) in fallopian tube epithelial cell lines suggested an association between loss of H2Bub1 and a more open chromatin configuration [62]. In summary, H2Bub1 is a master regulator of transcription, controlling gene expression in response to acute stimuli and developmental signals in a context specific manner and occupying a central role in histone cross-talk directing gene expression.
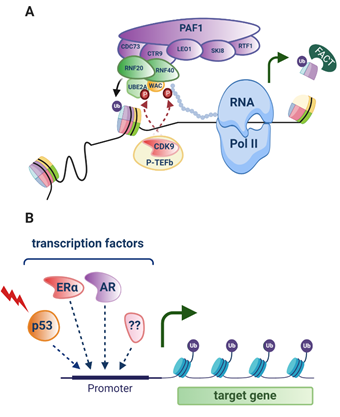
Figure 2. H2Bub1 in transcriptional elongation. (A) Following stimuli such as DNA damage, hormones or developmental signals, cyclin-dependent kinase 9 (CDK9) that is part of the Positive Transcription Elongation Factor-b (P-TEFb) complex phosphorylates Ser2 in the carboxy-terminal domain of RNA polymerase II (RNA pol II) depicted by a chain of circles, as well as the E2 ubiquitin conjugating enzyme ubiquitin conjugating enzyme E2 A (UBE2A). This dual phosphorylation shown by red circles, establishes a binding domain for the WW domain-containing adaptor with coiled-coil (WAC) and the E3 ubiquitin ligase complex RNF20/RNF40 that monoubiquitinates histone H2B at lysine 120 (H2Bub1, ubiquitin shown as a purple circle). RNF20/RNF40 physically interacts with the PAF1 complex consisting of CDC73, CTR9, LEO1, SKI8, RTF1 and PAF1 that associates with RNA pol II to establish transcriptional elongation. The chromatin remodelling factor FACT is recruited by H2Bub1, and removes a H2B-H2A dimer from the core nucleosome that takes away the physical block to RNA pol II, allowing it to move through the nucleosome, facilitating gene expression (green arrow). (B) A number of transcription factors have been associated with enriched H2Bub1 at target genes, including p53 (red lightning bolt depicts DNA damage that activates this tumour suppressor), the estrogen receptor (ERα) and androgen receptor (AR). . indicates as yet unknown transcription factors. Created with BioRender.com.
3. H2Bub1 and the DNA Damage Response (DDR)
H2Bub1 accumulates after DNA damage at sites of DSBs as part of the cellular DDR [14,63,64]. Upon DNA damage, the ATM (ataxia telangiectasia mutated) kinase phosphorylates RNF20 and RNF40, facilitating the recruitment of this E3 ligase complex to DSBs where it acts to catalyse H2Bub1 [14]. DNA damage-associated H2Bub1 then acts as a platform to recruit the chromatin remodelling factor SNF2H and other proteins required for homologous recombination repair (HRR) including BRCA1, BRCA2 and RAD51, as well as non-homologous end joining, including XRCC4 and Ku80 (Figures 1,3) [14,63,65].
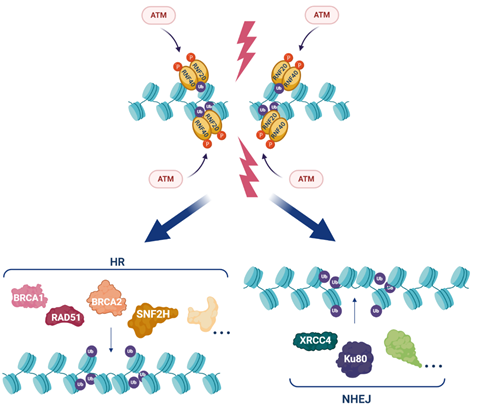
Figure 3. H2Bub1 in the DNA damage response. In the presence of double strand breaks (DSBs), ATM (ataxia telangiectasia mutated) phosphorylates (red circles) RNF20 and RNF40 that localise to DSBs where they function as an E3 ligase complex to write H2Bub1 (purple circles). H2Bub1 acts as a platform to attract proteins that work in both homologous recombination (HRR: BRCA1, BRCA2, RAD51, SNF2H and additional proteins) and non-homologous end joining (NHEJ: XRCC4, Ku80 (XRCC5) and additional proteins) to enable DNA repair. Created with BioRender.com.
Numerous factors that have a role in H2Bub1-related transcription also have a role in H2Bub1-related DNA repair. One of these is the histone chaperone FACT complex subunit Spt16 that binds to RNF20 to enable recruitment of SNF2H as well as the DNA repair proteins BRCA1 and RAD51 at DSBs to initiate HRR [66]. RNF40 was shown to have a similar role and interaction with Spt16 at sites of DSBs [67]. A number of the H2Bub1 DUBs have also been implicated in the DNA damage response through modulation of chromatin, including USP11 [68], USP22 [69,70] and USP44 [71]. Furthermore, the DOT1L histone methyltransferase known to engage in trans-histone cross-talk with H2Bub1 is important for HRR at DSBs [72].
Global loss of H2Bub1 has been observed in cancer cell line models following treatment with DNA damaging agents including doxorubicin, neocarzinostatin and cisplatin [14,23,24]. On the background of this global loss, H2Bub1 is enriched in the coding regions of specific highly expressed genes, including p53 target genes such as CDKN1A and MDM2 [23,24]. It is likely in these cases that many genes that display enriched H2Bub1 following DNA damage are required for decisions regarding cellular fate [23].
4. H2Bub1 and Cellular Proliferation in Cancer
It is well established that cellular proliferation is dependent upon access to chromatin to regulate the expression of oncogenes and tumour suppressors that when aberrantly expressed can lead to a malignant phenotype. Loss of H2Bub1 has been associated with tumour progression consistent with a tumour suppressive function of this histone PTM [1,10,12]. The role of H2Bub1 in cellular proliferation has been studied predominantly through manipulation of its key writer and eraser enzymes. For example, siRNA (short interfering RNA) down-regulation of RNF20 and RNF40 has been reported to inhibit proliferation of prostate cancer cells [42]. Of note, RNF40 has been implicated in the control of key apoptotic genes in colorectal cancer cells [73].
RNF20 has also been identified as necessary for proliferation, both in vitro in mixed-lineage leukemia (MLL)-rearranged human acute myeloid leukemia (AML) cell lines and in vivo, being important for disease progression in a genetically engineered mouse model of AML [74]. RNF20 and USP44 have different effects on the proliferation and migration of breast cancer cells dependent on subtype, i.e., basal-like versus luminal [22]. H2Bub1-associated DUBs USP22, USP51 and USP27X have been shown in vitro to be required for normal growth, with in vivo depletion suppressing tumour growth in a mouse xenograft model of breast cancer cells [75]. Down-regulation of USP36 in lung cancer cells has also been shown to negatively impact on cell proliferation [76].
5. Summary
In summary, H2Bub1 is a key histone modification influencing fundamental cellular activities including transcriptional elongation, the DDR and stem cell plasticity. It is increasingly recognised as a central histone PTM given its influence and/or interactions with other chromatin remodelling complexes such as COMPASS, DOT1L, SAGA and SWI/SNF. Continued expansion and elucidation of the H2Bub1 interactome offers new insights into cellular processes and extends our opportunities for therapeutic targeting of malignancy based on the epigenome and ubiquitin chromatin modelling.
This entry is adapted from the peer-reviewed paper 10.3390/cancers12113462