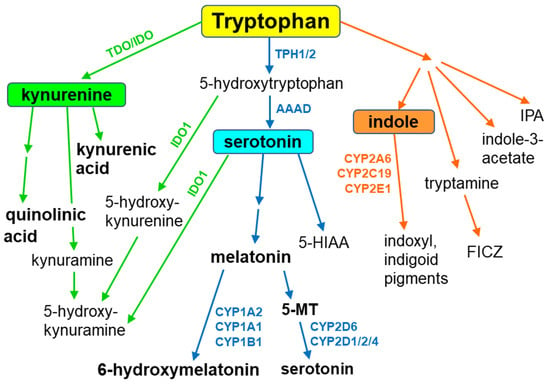
Tryptophan is metabolized along three main metabolic pathways, namely the kynurenine, serotonin and indole pathways. The majority of tryptophan is transformed via the kynurenine pathway, catalyzed by tryptophan-2,3-dioxygenase or indoleamine-2,3-dioxygenase, leading to neuroprotective kynurenic acid or neurotoxic quinolinic acid. Serotonin synthesized by tryptophan hydroxylase, and aromatic L-amino acid decarboxylase enters the metabolic cycle: serotonin → N-acetylserotonin → melatonin → 5-methoxytryptamine→serotonin. Recent studies indicate that serotonin can also be synthesized by cytochrome P450 (CYP), via the CYP2D6-mediated 5-methoxytryptamine O-demethylation, while melatonin is catabolized by CYP1A2, CYP1A1 and CYP1B1 via aromatic 6-hydroxylation and by CYP2C19 and CYP1A2 via O-demethylation. In gut microbes, tryptophan is metabolized to indole and indole derivatives. Some of those metabolites act as activators or inhibitors of the aryl hydrocarbon receptor, thus regulating the expression of CYP1 family enzymes, xenobiotic metabolism and tumorigenesis. The indole formed in this way is further oxidized to indoxyl and indigoid pigments by CYP2A6, CYP2C19 and CYP2E1. The products of gut-microbial tryptophan metabolism can also inhibit the steroid-hormone-synthesizing CYP11A1. In plants, CYP79B2 and CYP79B3 were found to catalyze N-hydroxylation of tryptophan to form indole-3-acetaldoxime while CYP83B1 was reported to form indole-3-acetaldoxime N-oxide in the biosynthetic pathway of indole glucosinolates, considered to be defense compounds and intermediates in the biosynthesis of phytohormones. Thus, cytochrome P450 is engaged in the metabolism of tryptophan and its indole derivatives in humans, animals, plants and microbes, producing biologically active metabolites which exert positive or negative actions on living organisms. Some tryptophan-derived metabolites may influence cytochrome P450 expression, affecting cellular homeostasis and xenobiotic metabolism.
- cytochrome P450
- tryptophan metabolism
- serotonin
- melatonin
1. Introduction
2. The Contribution of Cytochrome P450 to the Synthesis of Serotonin
In the brain, serotonin routes originate from neurons of the raphe nuclei, which are located in the brain stem. The dorsal raphe nuclei (DRN) and median raphe nuclei (MRN) neurons project to the forebrain, including the cortex, hippocampus, striatum and hypothalamus, and account for about 80% of forebrain serotonergic endings [26]. Serotonergic projections from the raphe nuclei innervate almost all the brain structures that are involved in controlling important physiological functions, i.e., the cortex (mood and sleep), hippocampus (stress, learning and memory), basal ganglia (motor functions), thalamus (sleep and epilepsy) and hypothalamus (neuroendocrine functions, food intake, circadian rhythm and thermoregulation) [27,28,29][27][28][29]. Thus, serotonin is engaged in the physiology and pathology of various psychiatric disorders and the action of neurological and psychotropic drugs. The biosynthesis of serotonin in the brain starts from the essential amino acid tryptophan and proceeds via hydroxylation to L-5-hydroxytryptophan and subsequent decarboxylation (Figure 2). The activity of tryptophan hydroxylase 2 (TPH2) is considered a concentration-limiting step in the biosynthesis of the neurotransmitter. Serotonin is then inactivated by monoamine oxidase (MAO-A) and aldehyde dehydrogenase to 5-hydroxyindole acetic acid (5-HIAA). Serotonin can also be formed in the gut by tryptophan hydroxylase 1 (TPH1) in enterochromaffin cells and stored in blood platelets. Peripheral serotonin plays an important role in gastrointestinal motility [30,31][30][31] and liver regeneration [32,33,34][32][33][34]. It is vitally important that serotonin produced in the periphery cannot penetrate the blood–brain barrier.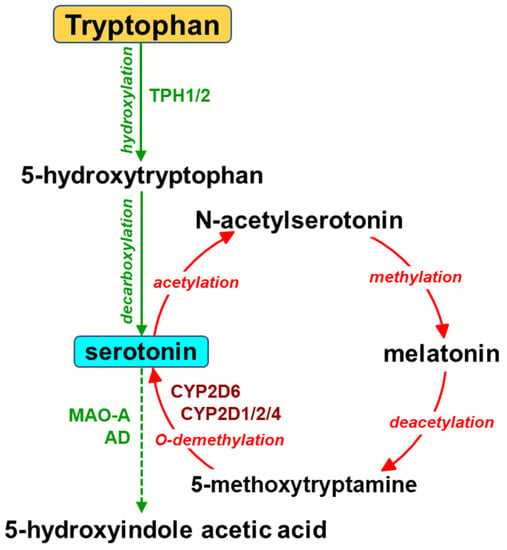
References
- Omura, T. Structural Diversity of Cytochrome P450 Enzyme System. J. Biochem. 2010, 147, 297–306.
- Fujiyama, K.; Hino, T.; Nagano, S. Diverse Reactions Catalyzed by Cytochrome P450 and Biosynthesis of Steroid Hormone. Biophys. Physicobiol. 2022, 19, e190021.
- Guengerich, F.P. Roles of Cytochrome P450 Enzymes in Pharmacology and Toxicology: Past, Present, and Future. Adv. Pharmacol. 2022, 95, 1–47.
- Nebert, D.W.; Nelson, D.R.; Coon, M.J.; Estabrook, R.W.; Feyereisen, R.; Fujii-Kuriyama, Y.; Gonzalez, F.J.; Guengerich, F.P.; Gunsalus, I.C.; Johnson, E.F. The P450 Superfamily: Update on New Sequences, Gene Mapping, and Recommended Nomenclature. DNA Cell. Biol. 1991, 10, 1–14.
- Crešnar, B.; Petrič, S. Cytochrome P450 Enzymes in the Fungal Kingdom. Biochim. Biophys. Acta 2011, 1814, 29–35.
- Hansen, C.C.; Nelson, D.R.; Møller, B.L.; Werck-Reichhart, D. Plant Cytochrome P450 Plasticity and Evolution. Mol. Plant 2021, 14, 1244–1265.
- Lamb, D.C.; Follmer, A.H.; Goldstone, J.V.; Nelson, D.R.; Warrilow, A.G.; Price, C.L.; True, M.Y.; Kelly, S.L.; Poulos, T.L.; Stegeman, J.J. On the Occurrence of Cytochrome P450 in Viruses. Proc. Natl. Acad. Sci. USA 2019, 116, 12343–12352.
- Zanger, U.M.; Schwab, M. Cytochrome P450 Enzymes in Drug Metabolism: Regulation of Gene Expression, Enzyme Activities, and Impact of Genetic Variation. Pharmacol. Ther. 2013, 138, 103–141.
- Roth, W.; Zadeh, K.; Vekariya, R.; Ge, Y.; Mohamadzadeh, M. Tryptophan Metabolism and Gut-Brain Homeostasis. Int. J. Mol. Sci. 2021, 22, 2973.
- Ravanfar, R.; Sheng, Y.; Gray, H.B.; Winkler, J.R. Tryptophan-96 in Cytochrome P450 BM3 Plays a Key Role in Enzyme Survival. FEBS Lett. 2023, 597, 59–64.
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, Tryptophan Metabolism and the Brain-Gut-Microbiome Axis. Behav. Brain Res. 2015, 277, 32–48.
- Fiore, A.; Murray, P.J. Tryptophan and Indole Metabolism in Immune Regulation. Curr. Opin. Immunol. 2021, 70, 7–14.
- Yu, A.-M.; Idle, J.R.; Byrd, L.G.; Krausz, K.W.; Küpfer, A.; Gonzalez, F.J. Regeneration of Serotonin from 5-Methoxytryptamine by Polymorphic Human CYP2D6. Pharmacogenetics 2003, 13, 173–181.
- Haduch, A.; Bromek, E.; Sadakierska-Chudy, A.; Wójcikowski, J.; Daniel, W.A. The Catalytic Competence of Cytochrome P450 in the Synthesis of Serotonin from 5-Methoxytryptamine in the Brain: An in Vitro Study. Pharmacol. Res. 2013, 67, 53–59.
- Haduch, A.; Bromek, E.; Kot, M.; Kamińska, K.; Gołembiowska, K.; Daniel, W.A. The Cytochrome P450 2D-Mediated Formation of Serotonin from 5-Methoxytryptamine in the Brain in Vivo: A Microdialysis Study. J. Neurochem. 2015, 133, 83–92.
- Hardeland, R. Taxon- and Site-Specific Melatonin Catabolism. Molecules 2017, 22, 2015.
- Haq, S.; Grondin, J.A.; Khan, W.I. Tryptophan-Derived Serotonin-Kynurenine Balance in Immune Activation and Intestinal Inflammation. FASEB J. 2021, 35, e21888.
- Liu, J.-R.; Miao, H.; Deng, D.-Q.; Vaziri, N.D.; Li, P.; Zhao, Y.-Y. Gut Microbiota-Derived Tryptophan Metabolism Mediates Renal Fibrosis by Aryl Hydrocarbon Receptor Signaling Activation. Cell. Mol. Life Sci. 2021, 78, 909–922.
- Gillam, E.M.; Notley, L.M.; Cai, H.; De Voss, J.J.; Guengerich, F.P. Oxidation of Indole by Cytochrome P450 Enzymes. Biochemistry 2000, 39, 13817–13824.
- Yacoub, R.; Wyatt, C.M. Manipulating the Gut Microbiome to Decrease Uremic Toxins. Kidney Int. 2017, 91, 521–523.
- Mosa, A.; Gerber, A.; Neunzig, J.; Bernhardt, R. Products of Gut-Microbial Tryptophan Metabolism Inhibit the Steroid Hormone-Synthesizing Cytochrome P450 11A1. Endocrine 2016, 53, 610–614.
- Adak, S.; Lukowski, A.L.; Schäfer, R.J.B.; Moore, B.S. From Tryptophan to Toxin: Nature’s Convergent Biosynthetic Strategy to Aetokthonotoxin. J. Am. Chem. Soc. 2022, 144, 2861–2866.
- Rannug, A.; Rannug, U. The Tryptophan Derivative 6-FormylindoloCarbazole, FICZ, a Dynamic Mediator of Endogenous Aryl Hydrocarbon Receptor Signaling, Balances Cell Growth and Differentiation. Crit. Rev. Toxicol. 2018, 48, 555–574.
- Celenza, J.L. Metabolism of Tyrosine and Tryptophan--New Genes for Old Pathways. Curr. Opin. Plant Biol. 2001, 4, 234–240.
- Barry, S.M.; Kers, J.A.; Johnson, E.G.; Song, L.; Aston, P.R.; Patel, B.; Krasnoff, S.B.; Crane, B.R.; Gibson, D.M.; Loria, R.; et al. Cytochrome P450–Catalyzed L-Tryptophan Nitration in Thaxtomin Phytotoxin Biosynthesis. Nat. Chem. Biol. 2012, 8, 814–816.
- Dahlström, A.; Fuxe, K. Localization of Monoamines in the Lower Brain Stem. Experientia 1964, 20, 398–399.
- Törk, I. Anatomy of the Serotonergic System. Ann. N. Y. Acad. Sci. 1990, 600, 9–34; discussion 34–35.
- Di Giovanni, G.; Di Matteo, V.; Pierucci, M.; Esposito, E. Serotonin-Dopamine Interaction: Electrophysiological Evidence. Prog. Brain Res. 2008, 172, 45–71.
- Bromek, E.; Daniel, W.A. The Regulation of Liver Cytochrome P450 Expression and Activity by the Brain Serotonergic System in Different Experimental Models. Expert. Opin. Drug. Metab. Toxicol. 2021, 17, 413–424.
- Keating, D.J.; Spencer, N.J. What Is the Role of Endogenous Gut Serotonin in the Control of Gastrointestinal Motility? Pharmacol. Res. 2019, 140, 50–55.
- Mawe, G.M.; Hurd, M.; Hennig, G.W.; Lavoie, B. Epithelial 5-HT4 Receptors as a Target for Treating Constipation and Intestinal Inflammation. Adv. Exp. Med. Biol. 2022, 1383, 329–334.
- Lesurtel, M.; Graf, R.; Aleil, B.; Walther, D.J.; Tian, Y.; Jochum, W.; Gachet, C.; Bader, M.; Clavien, P.-A. Platelet-Derived Serotonin Mediates Liver Regeneration. Science 2006, 312, 104–107.
- Fang, Y.; Liu, C.; Shu, B.; Zhai, M.; Deng, C.; He, C.; Luo, M.; Han, T.; Zheng, W.; Zhang, J.; et al. Axis of Serotonin -PERK-YAP in Liver Regeneration. Life Sci. 2018, 209, 490–497.
- Kimura, M.; Moteki, H.; Ogihara, M. Role of Hepatocyte Growth Regulators in Liver Regeneration. Cells 2023, 12, 208.
- Prozialek, W.C.; Vogel, W.H. Deamination of 5-Methoxytryptamine, Serotonin and Phenylethylamine by Rat MAO in Vitro and in Vivo. Life Sci. 1978, 22, 561–569.
- Kobayashi, S.; Murray, S.; Watson, D.; Sesardic, D.; Davies, D.S.; Boobis, A.R. The Specificity of Inhibition of Debrisoquine 4-Hydroxylase Activity by Quinidine and Quinine in the Rat Is the Inverse of That in Man. Biochem. Pharmacol. 1989, 38, 2795–2799.
- Boobis, A.R.; Sesardic, D.; Murray, B.P.; Edwards, R.J.; Singleton, A.M.; Rich, K.J.; Murray, S.; de la Torre, R.; Segura, J.; Pelkonen, O. Species Variation in the Response of the Cytochrome P-450-Dependent Monooxygenase System to Inducers and Inhibitors. Xenobiotica 1990, 20, 1139–1161.
- Cheng, J.; Zhen, Y.; Miksys, S.; Beyoğlu, D.; Krausz, K.W.; Tyndale, R.F.; Yu, A.; Idle, J.R.; Gonzalez, F.J. Potential Role of CYP2D6 in the Central Nervous System. Xenobiotica 2013, 43, 973–984.
- Patel, S.; Dulluc, J.; Geffard, M. Comparison of Serotonin and 5-Methoxytryptamine Immunoreactivity in Rat Raphe Nuclei. Histochemistry 1986, 85, 259–263.
- Kuban, W.; Daniel, W.A. Cytochrome P450 Expression and Regulation in the Brain. Drug. Metab. Rev. 2021, 53, 1–29.
- Kot, M.; Daniel, W.A. Cytochrome P450 Is Regulated by Noradrenergic and Serotonergic Systems. Pharmacol. Res. 2011, 64, 371–380.
- Miksys, S.; Rao, Y.; Sellers, E.M.; Kwan, M.; Mendis, D.; Tyndale, R.F. Regional and Cellular Distribution of CYP2D Subfamily Members in Rat Brain. Xenobiotica 2000, 30, 547–564.
- Miksys, S.; Rao, Y.; Hoffmann, E.; Mash, D.C.; Tyndale, R.F. Regional and Cellular Expression of CYP2D6 in Human Brain: Higher Levels in Alcoholics: CYP2D6 Is Higher in Brain of Human Alcoholics. J. Neurochem. 2002, 82, 1376–1387.
- Miksys, S.; Tyndale, R.F. Cytochrome P450-Mediated Drug Metabolism in the Brain. J. Psychiatry Neurosci. 2013, 38, 152–163.
- Mostalac-Preciado, C.R.; de Gortari, P.; López-Rubalcava, C. Antidepressant-like Effects of Mineralocorticoid but Not Glucocorticoid Antagonists in the Lateral Septum: Interactions with the Serotonergic System. Behav. Brain Res. 2011, 223, 88–98.
- Bromek, E.; Haduch, A.; Daniel, W.A. The Ability of Cytochrome P450 2D Isoforms to Synthesize Dopamine in the Brain: An in Vitro Study. Eur. J. Pharmacol. 2010, 626, 171–178.
- Borroto-Escuela, D.O.; Ambrogini, P.; Chruścicka, B.; Lindskog, M.; Crespo-Ramirez, M.; Hernández-Mondragón, J.C.; Perez de la Mora, M.; Schellekens, H.; Fuxe, K. The Role of Central Serotonin Neurons and 5-HT Heteroreceptor Complexes in the Pathophysiology of Depression: A Historical Perspective and Future Prospects. Int. J. Mol. Sci. 2021, 22, 1927.
- Correia, A.S.; Vale, N. Tryptophan Metabolism in Depression: A Narrative Review with a Focus on Serotonin and Kynurenine Pathways. Int. J. Mol. Sci. 2022, 23, 8493.
- Pourhamzeh, M.; Moravej, F.G.; Arabi, M.; Shahriari, E.; Mehrabi, S.; Ward, R.; Ahadi, R.; Joghataei, M.T. The Roles of Serotonin in Neuropsychiatric Disorders. Cell. Mol. Neurobiol. 2022, 42, 1671–1692.
- Haduch, A.; Pukło, R.; Alenina, N.; Nikiforuk, A.; Popik, P.; Bader, M.; Daniel, W.A. The Effect of Ageing and Cerebral Serotonin Deficit on the Activity of Cytochrome P450 2D (CYP2D) in the Brain and Liver of Male Rats. Neurochem. Int. 2020, 141, 104884.
- Haduch, A.; Danek, P.J.; Kuban, W.; Pukło, R.; Alenina, N.; Gołębiowska, J.; Popik, P.; Bader, M.; Daniel, W.A. Cytochrome P450 2D (CYP2D) Enzyme Dysfunction Associated with Aging and Serotonin Deficiency in the Brain and Liver of Female Dark Agouti Rats. Neurochem. Int. 2022, 152, 105223.
- Warner, M.; Gustafsson, J.A. Effect of Ethanol on Cytochrome P450 in the Rat Brain. Proc. Natl. Acad. Sci. USA 1994, 91, 1019–1023.
- Mann, A.; Miksys, S.; Lee, A.; Mash, D.C.; Tyndale, R.F. Induction of the Drug Metabolizing Enzyme CYP2D in Monkey Brain by Chronic Nicotine Treatment. Neuropharmacology 2008, 55, 1147–1155.
- Miller, R.T.; Miksys, S.; Hoffmann, E.; Tyndale, R.F. Ethanol Self-Administration and Nicotine Treatment Increase Brain Levels of CYP2D in African Green Monkeys. Br. J. Pharmacol. 2014, 171, 3077–3088.
- Yue, J.; Miksys, S.; Hoffmann, E.; Tyndale, R.F. Chronic Nicotine Treatment Induces Rat CYP2D in the Brain but Not in the Liver: An Investigation of Induction and Time Course. J. Psychiatry Neurosci. 2008, 33, 54–63.
- Haduch, A.; Bromek, E.; Daniel, W.A. The Effect of Psychotropic Drugs on Cytochrome P450 2D (CYP2D) in Rat Brain. Eur. J. Pharmacol. 2011, 651, 51–58.
- Haduch, A.; Rysz, M.; Papp, M.; Daniel, W.A. The Activity of Brain and Liver Cytochrome P450 2D (CYP2D) Is Differently Affected by Antidepressants in the Chronic Mild Stress (CMS) Model of Depression in the Rat. Biochem. Pharmacol. 2018, 156, 398–405.
- Haduch, A.; Daniel, W.A. The Engagement of Brain Cytochrome P450 in the Metabolism of Endogenous Neuroactive Substrates: A Possible Role in Mental Disorders. Drug Metab. Rev. 2018, 50, 415–429.
- Haduch, A.; Bromek, E.; Pukło, R.; Jastrzębska, J.; Danek, P.J.; Daniel, W.A. The Effect of the Selective N-Methyl-D-Aspartate (NMDA) Receptor GluN2B Subunit Antagonist CP-101,606 on Cytochrome P450 2D (CYP2D) Expression and Activity in the Rat Liver and Brain. Int. J. Mol. Sci. 2022, 23, 13746.
- Daniel, W.A.; Bromek, E.; Danek, P.J.; Haduch, A. The Mechanisms of Interactions of Psychotropic Drugs with Liver and Brain Cytochrome P450 and Their Significance for Drug Effect and Drug-Drug Interactions. Biochem. Pharmacol. 2022, 199, 115006.
- Hedlund, E.; Wyss, A.; Kainu, T.; Backlund, M.; Köhler, C.; Pelto-Huikko, M.; Gustafsson, J.A.; Warner, M. Cytochrome P4502D4 in the Brain: Specific Neuronal Regulation by Clozapine and Toluene. Mol. Pharmacol. 1996, 50, 342–350.
- Danek, P.J.; Bromek, E.; Haduch, A.; Daniel, W.A. Chronic Treatment with Asenapine Affects Cytochrome P450 2D (CYP2D) in Rat Brain and Liver. Pharmacological Aspects. Neurochem. Int. 2021, 151, 105209.
- Danek, P.J.; Daniel, W.A. Long-Term Treatment with Atypical Antipsychotic Iloperidone Modulates Cytochrome P450 2D (CYP2D) Expression and Activity in the Liver and Brain via Different Mechanisms. Cells 2021, 10, 3472.
- Danek, P.J.; Daniel, W.A. The Atypical Antipsychotic Lurasidone Affects Brain but Not Liver Cytochrome P450 2D (CYP2D) Activity. A Comparison with Other Novel Neuroleptics and Significance for Drug Treatment of Schizophrenia. Cells 2022, 11, 3513.
- Bertilsson, L.; Alm, C.; De Las Carreras, C.; Widen, J.; Edman, G.; Schalling, D. Debrisoquine Hydroxylation Polymorphism and Personality. Lancet 1989, 1, 555.
- González, I.; Peñas-Lledó, E.M.; Pérez, B.; Dorado, P.; Alvarez, M.; LLerena, A. Relation between CYP2D6 Phenotype and Genotype and Personality in Healthy Volunteers. Pharmacogenomics 2008, 9, 833–840.
- Peñas-LLedó, E.M.; LLerena, A. CYP2D6 Variation, Behaviour and Psychopathology: Implications for Pharmacogenomics-Guided Clinical Trials. Br. J. Clin. Pharmacol. 2014, 77, 673–683.
- Hensler, J.G. Serotonergic Modulation of the Limbic System. Neurosci. Biobehav. Rev. 2006, 30, 203–214.