Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Liping Lou and Version 2 by Catherine Yang.
Antibiotic resistance genes (ARGs) and antibiotic-resistant bacteria (ARB) in soil have become research hotspots in the fields of public health and environmental ecosystem. The soil environment is an important acceptor of many pollutants, including antibiotics. The external pressure of antibiotics and other pollutants can promote the proliferation and occurrence of antibiotic-resistant bacteria (ARB) and antibiotic-resistance genes (ARGs) in soil. Soil ARGs can spread in various ways, such as horizontal gene transfer (HGT) between soil microorganisms and vertical gene transfer (VGT) between parent and offspring, while soil components have important influence on the occurance and spread of ARGs.
- soil component
- antibiotic resistance genes
- horizontal gene transfer
1. The Occurrence and Spread of ARGs in Soil
1.1. The Pollution Status of ARB and ARGs in Soil
Soil is the largest reservoir of ARBs and ARGs [1][2][14,15]. The abundance of ARGs in soil has increased substantially since the beginning of the antibiotic era [3][4][16,17]. Similar to antibiotics, wastewater irrigation and manure application are two main routes for ARBs and ARGs entering the soil [2][5][15,18].
ARBs and ARGs have been widely detected in different types of soils around the world [6][19]. Taking E. coli, which has been widely studied, as an example, the existence of E. coli has been found in farmland soil, non-farmland soil, and even plant microbial communities [7][8][20,21]. Many E. coli strains isolated from soil carry ARGs, and most of them have multidrug resistance. Furlan et al. [9][22] isolated a total of 60 strains of E. coli from soil samples on Brazilian farms, of which 68.3% of them exhibited multidrug resistance profiles. Liu et al. [8][21] found that all soil E. coli isolated from Washington State (n = 1905) were resistant to at least four different antibiotics. Graves et al. [10][23] analyzed 616 strains of E. coli collected from swine manure, swine lagoon effluent, and soils that received lagoon effluent and found that these strains usually carried ARGs coding for streptomycin, spectinomycin, tetracycline, and sulfonamide.
At the same time, the relative abundance of various ARGs in soil has increased significantly in recent years, and the increase in tetracycline resistance gene levels was significantly more frequent than for other ARGs [11][24]. Tetracyclines ARGs are present in soils worldwide, and the relative abundance is between 10−9 and 10−2 copies/16S rRNA gene (Table 1). Tetracyclines ARGs are also the main types of ARGs carried by phages in the soil environment [12][25].
Table 1. Antibiotic-resistance genes (ARGs) contamination in soil (copies/16S rRNA gene).
| Place | Soil Type | ARGs | Relative Abundance |
|---|---|---|---|
| China [13][26] | Feedlot vicinity | tetM, tetO, tetQ, tetW | 10−5–10−2 |
| China [14][27] | Feedlot vicinity | tetB(P), tetM, tetO, tetW | 10−3–10−6 |
| China [15][28] | Feedlots | tetA(P), tetG, tetC, tetL, tetX, tetM, tetA | 10−2–10−4 |
| China [16][29] | Feedlots | tetA, tetB, tetM | 10−6–10−1 |
| China [17][30] |
]. Only competent bacteria can obtain extracellular DNA, and the competence can be naturally or artificially induced [38][52]. Most naturally transformable bacteria can develop into competent cells under specific circumstances (e.g., nutrient conditions, changes in bacterial density [39][53]). Johann et al. [40][54] listed 87 species of bacteria that can absorb extracellular free DNA through natural transformation, including Pseudomonas and Acinetobacter, which are commonly found in soil. The key steps of ARGs transformation are as follows: (1) bacteria actively or passively discharge ARGs into the environment; (2) extracellular ARGs become stable and ingestible in the environment; (3) extracellular ARGs are ingested into the bacterial cytoplasm; (4) exogenous ARGs integrate into bacterial chromosomes by homologous recombination or replicate autonomously as episomes [37][38][39][41][51,52,53,55]. Chen et al. [42][56] found that the plasmid pK5 carrying ARGs had a strong migration ability in soil, which confirmed the widespread occurrence of the transformation process in soil.
Conjugation refers to the process by which the plasmid or chromosome carrying ARGs enters the recipient bacteria through the conjugative fimbriae produced by the donor bacteria [43][44][57,58]. Conjugation is considered to provide better protection from the surrounding environment and a more efficient means of genetic material entering the host cell than transformation, while often having a broader host range than bacteriophage transduction [32][46]. Integrative and conjugative elements (ICEs) and plasmids are the main vectors for the delivery of ARGs in conjugation [45][46][47][59,60,61]. Plasmids, as important mediators of conjugation, can still persist among bacterial populations without antibiotic stress and invade new strains with high frequency [48][49][62,63]. It can be classified into three categories according to mobility: conjugative, mobilizable, and non-mobilizable [45][59]. A conjugative plasmid codes for its own set of mating pair formation (MPF) genes; if it uses an MPF of another genetic element present in the cell, it is called mobilizable; other plasmids are called non-mobilizable because they are neither conjugative nor mobilizable and usually spread through transformation and transduction [45][47][59,61]. In addition, non-mobilizable plasmids can also be transferred by physical association with conjugative plasmids [50][64]. Plasmid-mediated conjugation includes multiple processes such as mating pair formation and relaxosome formation [47][51][61,65]. In soil, this process is susceptible to a variety of factors, such as soil bacterial population structure [52][66], nutrient composition [53][67], selective pressure of antibiotics and heavy metals [54][68], etc. However, the understanding of the plasmid-mediated conjugative transfer process of the complex bacterial community in soil is still limited [55][69]. ICEs have the properties of transposons, plasmids, and phages: both ICEs and transposons can jump on chromosomes, but transposons cannot undergo HGT; both ICEs and plasmids can transfer DNA between cells in the form of conjugation, but most ICEs cannot self-replicate as plasmids do; ICEs and phages both can detach, integrate, and replicate with host chromosomes, the difference being that ICEs transfer DNA in the form of conjugation rather than transduction [56][70]. The study by Gonçalves et al. [57][71] confirmed the role of ICEs in the soil microbial HGT process.
In transduction, ARGs are transferred from one bacterium to another by means of phages (bacteriovirus) and can be integrated into the chromosome of the recipient cell [58][72]. The phage-mediated transduction progress does not need contact between the donor and recipient, or even the simultaneous appearance of them [59][73]. Soil is one of the important habitats for phages and their hosts [60][61][74,75]. It is estimated that the number of soil virus particles (mainly phages) accounts for 10% of the total number of viruses in the world, about 4.8 × 1030 [62][76]. The special protein capsid structure of phages can effectively protect nucleic acids, and the soil is highly heterogeneous and rich in biodiversity, providing a variety of parasitic environments for phages [36][50]. The opaque environment of soil protects the phages from sun damage [63][77], which is more conducive to their long-term survival and reproduction. Therefore, compared with free ARGs and bacteria in soil, phages are more resistant to adverse environmental factors [64][78] and persist longer, providing a material basis for their interactions with bacteria and gene transfer. Related studies have shown that the contribution of phages to HGT was likely underestimated [65][66][79,80]. Olatz et al. [67][81] found a large number of free and replicable phages containing ARGs in farmland soil, which may lead to the production and enrichment of ARB.
Although HGT is regarded as the major pathway of ARGs spreading, there is a significant involvement of VGT. Firstly, VGT raises the possibility of spontaneous mutation of bacterial DNA [41][55]. Secondly, VGT promotes HGT among the bacterial community: Li et al. [68][4] found that VGT can significantly promote the formation of conjugants and accelerate the spread of ARGs.
2. Effects of Soil Components on the HGT of ARGs from the Micro Perspective
2.1. The Effects of Soil Components on the HGT of ARGs in Pure Bacterial System
Another part of the studies discussed the effect of soil components on HGT processes such as ARGs transformation and conjugation using the pure bacterial system (Table 2 and Table 3).Table 2. Research on the effects of soil components on transformation of antibiotic resistance genes (ARGs).
| Medium | Important Conclusions | ||||||
|---|---|---|---|---|---|---|---|
| Results | Reasons | ||||||
| Kaolinite, illite, and montmorillonite [69][12] | Plasmids adsorbed on minerals could resist higher concentrations of nucleases and form more transformants than free plasmids. | The adsorption of the nuclease on minerals protected the plasmids, but it can still be involved in transformation. | |||||
| Kaolinite, Goethite, and montmorillonite [Dissolved biochar [81]70][95] |
[105Low concentrations (1–2 g/L) have little effect; high concentration (10 g/L) of kaolinite and montmorillonite inhibited transformation; high concentration (10 g/L) of goethite promoted transformation. | ]Kaolinite and montmorillonite: strong adsorption to competence stimulating factor, decrease the expression level of competent genes (phrC, comS); | The effects on conjugation were related to the concentration and source of biochar. goethite: increase cell membrane damage. |
||||
| Humic acid-like substance in dissolved biochar improved the conjugative efficiency. | The inhibitory effects of small-molecule matters dominated, decreasing conjugative transfer frequency. | Montmorillonite [71][96] | Low concentration (about 0–0.025 g/L) promoted transformation; high concentration (about 0.025–2 g/L) inhibited transformation. |
Low concentration: increase the contact between plasmids and cells; forming holes on cell membrane; High concentration: plasmids were adsorbed; heavy metals released from montmorillonite cause the aggregation of the plasmids. |
|||
| Pyroligneous acid and its three fractions [79][103] | Reduced the abundance of ARGs and MGEs in soil. | High content of organic acids inhibited the bacterial growth. | Biochar [72][97] | Significantly inhibited the transformation of extracellular antibiotic resistance genes (eARGs) | |||
| Dissolved biochar [78][102] | Biochar dissolutions: Induce intramolecular condensation and agglomeration of plasmids; decrease the cell membrane permeability; | biochar solids: Adsorb plasmids and deactivate |
Attenuated the promotion effect of Cu (Ⅱ) to conjugation.E. coli. | Dissolved biochar affected intracellular ROS production level, cell membrane permeability, and the expression level of global regulatory genes (korA, korB, trbA), pore formation and membrane trafficking genes (ompA, ompC), MPF system gene (trbB), DTR system gene (trfA), etc. | Farmland | tetB(P), tetM, tetO, tetQ, tetT, tetW | 10−8–10−2 |
| China [18][31] | Farmland | tet | |||||
| CeO2 nanoparticle [80][104] (soil pollutant) |
Inhibited conjugation at low concentration (1, 5 mg/L), while promoted it at high concentration (25, 50 mg/L). | CeO2 nanoparticle affected many aspects, such as intracellular ROS production, polysaccharide synthesis in EPS, cell-to-cell contact, ATP supply, and the expression level of conjugation-related genes (MPF system gene (trbBp), DTR system gene (trfAp), putative transmembrane ATPase gene (traG)), etc. | |||||
| Gut of C. elegans [82 | G, | tetY, tetZ | ][10−7–10−4 | ||||
| 106 | China [19][32] | Farmland | tetB(P), tetC, tetG, tetL, tetO, tetS, tetW, tetZ | 10−6–10−1 | |||
| Italy [20][33] | Feedlots | tetQ, tetW | 10−9–10−5 | ||||
| India [21][34] | Feedlots | tetA, tetW | 10−1 a | ||||
| ] |
America [22][35] | Farmland | tetO, tetW | 10−7–10−4 | |||
| Austria [23][36] | Farmland | tetW | 10−5–10−4 | ||||
| The Netherlands [11][24] | Typical sites | tetM, tetO, tetQ, tetW | 10−4–10−2 | ||||
| Scotland [3][16] | Typical sites | tetM, tetQ, tetW | 10−5–10−2 | ||||
| Scotland [24][37] | Farm | tetA, tetB, tetC, tetG, tetW | 10−6–10−5 | ||||
| Australia [25][38] | Residential area | tetM, tetW | 10−9–10−2 | ||||
a The ratio of phages carrying ARGs to the total number of phages.
1.2. Transmission Routes of ARGs
The causes, inheritance, and transmission mechanisms of antibiotic resistance are very complex and can be divided into genetic resistance and non-inherited resistance [26][39] (Figure 1). Non-inherited resistance refers to resistance that is not acquired through horizontal or vertical transfer of ARGs but through behaviors such as collaboration between groups [27][40]. Cooperative resistance, a population-based survival strategy that adapts to high antibiotic stress through the cooperation of multiple ARBs, is a typical non-inherited resistance [28][41].
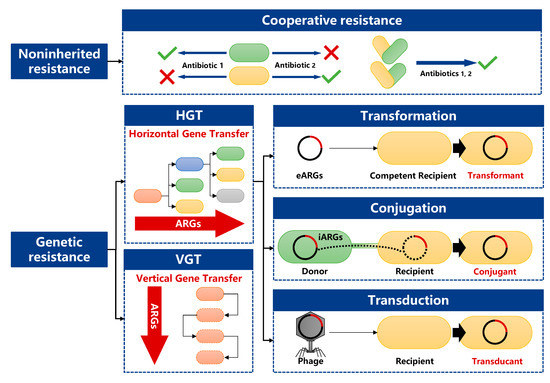
Figure 1.
Transmission routes of antibiotic resistance genes (ARGs: antibiotic resistance genes, eARGs: extracellular ARGs, iARGs: intracellular ARGs).
Genetic resistance can be divided into intrinsic resistance and acquired resistance. Intrinsic resistance refers to the natural existence of certain genes in bacterial genomes that could generate a resistance phenotype [29][43]. It is an ancient, natural, and widespread environmental phenomenon that predates the selective pressures caused by modern human use of antibiotics, and multiple ARGs have been detected in Arctic permafrost unaffected by human activities [6][30][19,44]. Acquired resistance is a consequence of spontaneous chromosomal mutations or ARGs gained through HGT [31][45], which refers to the exchange of genetic material between individuals of different organisms and is the key reason for the widespread existence of ARB in clinical systems [32][46]. After obtaining ARGs through HGT, ARGs will achieve the amplification and continuation of these genes through reproduction between parent and child generations in VGT [33][47].
HGT mainly includes three pathways mediated by mobile genetic elements (MGEs), namely extracellular DNA-mediated transformation, plasmid-mediated conjugation, and phage-mediated transduction [32][46]. A large number of research results have shown that HGT can widely occur in the soil environment [34][35][36][48,49,50].
Transformation refers to the process by which competent bacteria take up DNA from outside. Unlike conjugation, transformation does not require physical contact between the donor and recipient cells, and free DNA released by cell lysis can serve as the donor for transformation [37][51
Existing studies on transformation not only discuss the changes of ARGs vectors (plasmids, chromosomes, etc.) and recipient bacteria in soil microcosms, EPS, sediments, or other media but also include studies on the addition of single components to simulate soil conditions [70][71][72][76][95,96,97,101]. Relevant research (Table 2) has shown that plasmids [69][12] and chromosomes [73][98] adsorbed by soil components can still participate in transformation; Chamier et al. [74][99] found that the plasmid adsorbed on sand transformed significantly less efficiently than the plasmid in solution; but Dong et al. [76][101] considered that sediment-adsorbed plasmids had higher transformation efficiency than episomal plasmids. Montmorillonite at low concentrations (0–0.025 g/L) [71][96] and goethite at high concentrations (10 g/L) [70][95] promote transformation, while high concentrations of kaolinite (10 g/L), montmorillonite (0.025–2 g/L and 10 g/L), and biochar (2, 4, and 8 g/L) inhibit it [70][71][72][95,96,97].
The research on conjugation (Table 3) showed that birnessite and low concentrations of goethite (0–0.5 g/L) promoted conjugation; the effects of kaolinite and montmorillonite were irregular; goethite at high concentration (5 g/L) inhibited conjugative transfer [77][13]. Liu et al. [78][102] found that biochar can weaken the promoting effect of heavy metals on conjugation, while Zheng et al. [79][103] reported that pyroligneous acid and its three fractions at different temperatures had inhibitory effects on conjugative transfer. Some studies illustrated the mechanisms of soil components affecting the process of conjugation by detecting the expression of related genes [77][78][80][13,102,104], but most of them are speculation based on transcriptome results, and the understanding of related pathways and mechanisms is still unclear, which is worth exploring in depth.
| Soil microcosm | ||
| [ | ||
| 73 | ||
| ][98] | DNA adsorbed on soil particles still transformed competent cells | Minerals did not inhibit the transformation, but blocked DNA contact with the recipient. |
| Soil microcosm [74][99] | Plasmid adsorbed on sand transformed significantly less efficient than did plasmid in solution; the transformation by sand-adsorbed chromosomal was as high as that by plasmid in solution. |
Transformation occurred by direct uptake of DNA from the mineral surfaces; transformation requires multiple plasmids, and the probability of multiple free plasmids meeting bacteria at the same time is higher than that on mineral surfaces; the chances of bacteria taking up DNA on the mineral surface are proportional to the size of the DNA, and chromosomes of the same mass are larger and easier to take up. |
| Activated sludge EPS [75][100] | The transformation ability of free ARGs was higher than that in activated sludge extracellular polymeric substances (EPS) when calculated per ng DNA, and lower when calculated per g volatile suspended solids. | Activated sludge EPS is rich in ARGs. |
| Sediment [76][101] | The transformation efficiency of adsorbed eARG was higher than that of free eARGs. | Sand adsorbed bacteria and plasmids at the same time, facilitating contact between the two, and was related to the conformation of the plasmid. |
Table 3. Research on the effects of soil components on conjugation of antibiotic resistance genes (ARGs).
| Medium | Results | Reasons |
|---|---|---|
| Kaolinite, goethite, birnessite, and montmorillonite [77][13] | Birnessite promoted conjugation. The effects of kaolinite and montmorillonite were irregular. Goethite promoted conjugation at low concentration (0–0.5 g/L) and inhibited it at high concentration (5 g/L). |
Birnessite promoted the production of intracellular reactive oxygen species (ROS); increased the expression levels of oxidative stress-regulated genes (rpoS) and outer membrane protein genes (ompA, ompF, ompC). Birnessite altered the expression levels of conjugation-related genes (globally regulation genes (korA, korB, trbA); mating pair formation (MPF) system genes (trbBp, traF); DNA transfer and replication (DTR) system genes (trfAp, traJ)). |
| (soil animal) | ||
| The conjugation efficiency in gut was higher than soil, and increased with time and temperature. | ||
| The abundance of MPF system gene ( | trb | Bp) and DTR system gene (trfAp) was increased. |