Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Jan Mojzis and Version 2 by Lindsay Dong.
Chalcones, plant-derived polyphenols, received marked attention due to their biological activities as well as for relatively easy structural manipulation and synthesis of new chalcone derivatives. The chemotherapeutic effects of chalcones are mediated by numerous activities including the initiation of cell death machinery, suppression of cancer cell growth, modulation of several signaling pathways such as Wnt/β-catenin pathway, nuclear factor kappa B pathway, mitogen-activated protein kinase (MAPK) pathway and many others. In addition, chalcones seem to have an anticancer effect also via the inhibition of angiogenesis, production of reactive oxygen species (ROS)ROS, anti-H. pylori activity and anti-inflammatory action.
- chalcones
- colorectal cancer
- gastric cancer
- cell death
- angiogenesis
- inflammation
1. Introduction
A large family of natural compounds are polyphenols. These plant secondary metabolites are found in vegetables, fruits and seeds and are broadly distributed throughout plants [1][7]. They control several plant functions, such as protection from pathogens, UV irradiation and oxidative stress [2][8]. In addition to plant protection, numerous studies have documented the beneficial effects of these compounds on human health, including anticancer, anti-inflammatory, antioxidant and cardioprotective activities [3][4][9,10].
In recent decades, a large amount of attention has been focused on chalcones, which are the metabolic precursors of flavonoids. Not only are they abundant in nature and relatively easy to synthesize but their structural simplicity also carries a great deal of potential for the synthesis of a plethora of derivatives with pleiotropic bioactivities due to interactions between numerous macromolecules and various signaling pathways [5][6][11,12]. Many studies have described the broad spectrum of biological activities of chalcones, including anti-inflammatory, antimicrobial, antiparasitic, antidiabetic and antioxidant activities [7][8][9][10][11][13,14,15,16,17]. Moreover, the anticancer effects of both natural and synthetic chalcones have also been intensively studied [12][13][14][18,19,20].
Growing data have indicated that chalcones are multitarget anticancer compounds. The possible molecular mechanisms involved in the tumor-suppressive activity of these compounds are shown in Figure 1.
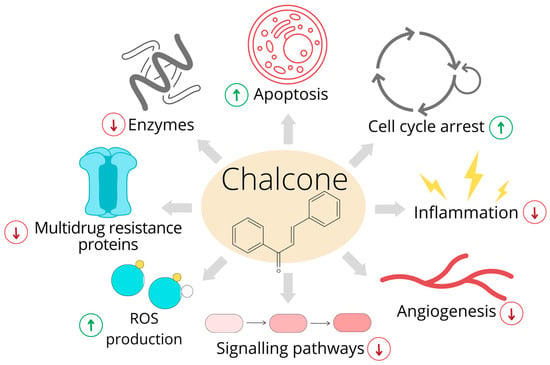
Figure 1. Anticancer effects of chalcones. The original figure was made using the Canva software by Radka Michalkova.
2. Molecular and Cellular Mechanisms of Action
2.1. Effects on Cell Cycle and Apoptosis
Cell cycle arrest prevents the proliferation of damaged cells. During this process, cells facilitate DNA repair or, if the DNA defects are too extensive, other signaling pathways are activated for cell removal [15]. Several chalcones have been found to block cancer cell proliferation via cell cycle arrest and the activation of cell death machinery. Despite their ability to inhibit the cell cycle in different phases [16][17][18], most chalcones have been observed to block cell cycle progression at the G2/M phase.
Scholars evaluated the mechanisms of numerous synthetic chalcone derivatives using either Caco-2 or HCT116 human colorectal cell lines. Flow cytometric analysis showed that several chalcones (e.g., (E)-2-(4′-methoxybenzylidene)-1-benzosuberone, (E)-2-(2′,4′-dimethoxybenzylidene)-1-tetralone and (2 E)-3-(acridin-9-yl)-1-(2,6-dimethoxyphenyl) prop-2-en-1-one) caused an accumulation of cancer cells in the G2/M phase, with a concomitant increase in cells with sub-G0/G1 DNA content (a marker for apoptosis) [19][20][21]. Three decades ago, chalcones were discovered to be antimitotic compounds [22]. Because the accumulation of cells in the G2/M phase could be a consequence of mitotic spindle dysregulation, scholars evaluated the effects of the studied chalcones on tubulin expression. The results showed that the expression of tubulins was deregulated on both the genomic and protein levels.
Chalcones play an important role in the induction of both intrinsic and extrinsic apoptosis pathways [23]. Several studies have demonstrated the ability of chalcones to induce the intrinsic apoptosis (mitochondrial) pathway. The structure and function of mitochondria are significantly changed in cells that have been exposed to apoptotic stimuli [24]. The modulation of Bcl-2 protein family activity leads to the permeabilization of the mitochondrial outer membrane, the loss of mitochondrial membrane potential (MMP) and the subsequent release of proapoptotic factors, followed by caspase activation and cell death [25][26]. In various colon cancer cells, chalcone treatment leads to either the activation of the proapoptotic members of the Bcl-2 protein family (e.g., Bax and Bad) or the suppression of antiapoptotic proteins (e.g., Bcl-xL and Bcl-2), resulting in the release of several proapoptotic proteins (e.g., cytochrome and Smac/DIABLO) and the activation of initiator caspase 9 or execution caspases 3/7. Moreover, a decrease in MMP has also often been detected [20][21][27][28][29].
Another important player in apoptosis regulation is the protein p53, which is a tumor suppressor activated by DNA damage. Its phosphorylation (i.e., activation) and subsequent caspase activation have been observed in sappanchalcone-treated HCT116 cells. On the other hand, colon cancer cells with mutated p53 have been found to be insensitive to sappanchalcone treatment [30]. A study by Shin et al. [31] showed a high level of p53 and apoptosis induction in chalcone-treated HCT116 cells. Interestingly, they also found apoptosis induction in p53-null HCT116 cells, suggesting that both p53-dependent and p53-independent mechanisms can play roles in apoptosis that is induced by this chalcone. In addition, several other publications have revealed an association between p53 activation and apoptosis induction in chalcone-treated colon cancer cells [32][33][34][35]. Furthermore, both natural and synthetic chalcones have been studied as potential anticancer agents in gastric carcinogenesis models [12][36].
Anticancer effects of chalcones. The original figure was made using the Canva software by Radka Michalkova.
2. Molecular and Cellular Mechanisms of Action
2.1. Effects on Cell Cycle and Apoptosis
Cell cycle arrest prevents the proliferation of damaged cells. During this process, cells facilitate DNA repair or, if the DNA defects are too extensive, other signaling pathways are activated for cell removal [21]. Several chalcones have been found to block cancer cell proliferation via cell cycle arrest and the activation of cell death machinery. Despite their ability to inhibit the cell cycle in different phases [22,23,24], most chalcones have been observed to block cell cycle progression at the G2/M phase.
Scholars evaluated the mechanisms of numerous synthetic chalcone derivatives using either Caco-2 or HCT116 human colorectal cell lines. Flow cytometric analysis showed that several chalcones (e.g., (E)-2-(4′-methoxybenzylidene)-1-benzosuberone, (E)-2-(2′,4′-dimethoxybenzylidene)-1-tetralone and (2 E)-3-(acridin-9-yl)-1-(2,6-dimethoxyphenyl) prop-2-en-1-one) caused an accumulation of cancer cells in the G2/M phase, with a concomitant increase in cells with sub-G0/G1 DNA content (a marker for apoptosis) [25,26,27]. Three decades ago, chalcones were discovered to be antimitotic compounds [28]. Because the accumulation of cells in the G2/M phase could be a consequence of mitotic spindle dysregulation, scholars evaluated the effects of the studied chalcones on tubulin expression. The results showed that the expression of tubulins was deregulated on both the genomic and protein levels.
Chalcones play an important role in the induction of both intrinsic and extrinsic apoptosis pathways [36]. Several studies have demonstrated the ability of chalcones to induce the intrinsic apoptosis (mitochondrial) pathway. The structure and function of mitochondria are significantly changed in cells that have been exposed to apoptotic stimuli [37]. The modulation of Bcl-2 protein family activity leads to the permeabilization of the mitochondrial outer membrane, the loss of mitochondrial membrane potential (MMP) and the subsequent release of proapoptotic factors, followed by caspase activation and cell death [38,39]. In various colon cancer cells, chalcone treatment leads to either the activation of the proapoptotic members of the Bcl-2 protein family (e.g., Bax and Bad) or the suppression of antiapoptotic proteins (e.g., Bcl-xL and Bcl-2), resulting in the release of several proapoptotic proteins (e.g., cytochrome and Smac/DIABLO) and the activation of initiator caspase 9 or execution caspases 3/7. Moreover, a decrease in MMP has also often been detected [26,27,31,40,41].
Another important player in apoptosis regulation is the protein p53, which is a tumor suppressor activated by DNA damage. Its phosphorylation (i.e., activation) and subsequent caspase activation have been observed in sappanchalcone-treated HCT116 cells. On the other hand, colon cancer cells with mutated p53 have been found to be insensitive to sappanchalcone treatment [42]. A study by Shin et al. [43] showed a high level of p53 and apoptosis induction in chalcone-treated HCT116 cells. Interestingly, they also found apoptosis induction in p53-null HCT116 cells, suggesting that both p53-dependent and p53-independent mechanisms can play roles in apoptosis that is induced by this chalcone. In addition, several other publications have revealed an association between p53 activation and apoptosis induction in chalcone-treated colon cancer cells [44,45,46,47]. Furthermore, both natural and synthetic chalcones have been studied as potential anticancer agents in gastric carcinogenesis models [18,48].
Additionally, it has been reported that the antiproliferative effects of other chalcones, such as isoliquiritigenin analog [37] and flavokawain B [38], are associated with G2/M cell cycle arrest. Furthermore, it is important to mention that several chalcones also initiate cell death via the activation of the extrinsic apoptosis pathway [39][40][41][42] or the induction of non-apoptotic cell death [38][43][44][45]. General pathways involved in the cell cycle and apoptosis are presented in
Additionally, it has been reported that the antiproliferative effects of other chalcones, such as isoliquiritigenin analog [52] and flavokawain B [53], are associated with G2/M cell cycle arrest. Furthermore, it is important to mention that several chalcones also initiate cell death via the activation of the extrinsic apoptosis pathway [54,55,56,57] or the induction of non-apoptotic cell death [53,58,59,60]. General pathways involved in the cell cycle and apoptosis are presented in Figure 2
.
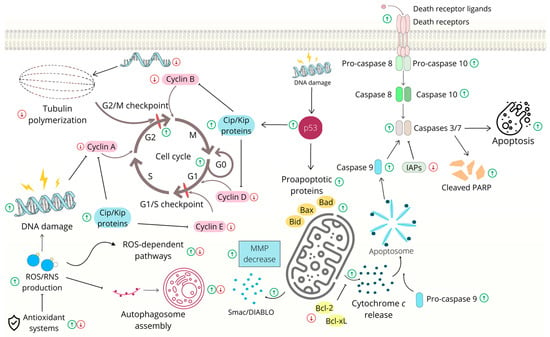
Figure 2.
Cell cycle arrest and apoptosis induced by chalcones in gastrointestinal cancer cells. Abbreviations: (pro)Casp—(pro)caspase, Bcl-2/Bcl-Xl—antiapoptotic proteins, Bad/Bax/Bid—proapoptotic proteins, Cip/Kip—CDK interacting protein/Kinase inhibitory proteins, IAPs—inhibitors of apoptosis, MMP- mitochondrial outer membrane permeabilization, PARP—poly (ADP-ribose) polymerase, ROS/RNS—reactive oxygen species/reactive nitrogen species, Smac/DIABLO—second mitochondria-derived activator of caspase/direct inhibitor of apoptosis-binding protein with low pI. The original figure was made using the Canva software by Radka Michalkova.
2.2. Modulation of Signaling Pathways
Several signaling pathways that are involved in cell survival and death frequently mutate during cancer progression [46][61].