Several environmental stresses, including biotic and abiotic factors, adversely affect the growth and development of crops, thereby lowering their yield. However, abiotic factors, e.g., drought, salinity, cold, heat, ultraviolet radiations (UVr), reactive oxygen species (ROS), trace metals (TM), and soil pH, are extremely destructive and decrease crop yield worldwide. The response of different plant species to such stresses is a complex phenomenon with individual features for several species. Metaetabolomics is a promising way to interpret biotic and abiotic stress tolerance in plants. The study of metabolic profiling revealed different types of metabolites, e.g., amino acids, carbohydrates, phenols, polyamines, terpenes, etc, which are accumulated in plants. Among all, primary metabolites, such as amino acids, carbohydrates, lipids polyamines, and glycine betaine, are considered the major contributing factors that work as osmolytes and osmoprotectants for plants from various environmental stress factors. Therefore, In contrast, plant-derived secondary metabolomics practices are becoming essential and influential inites, e.g., phenolics, terpenoids, and nitrogen-containing compounds (alkaloids), have no direct role in the growth and development of plants by identifying different phytochemicals that are part of the acclimation responses to various stimuli. Nevertheless, such metabolites could play a significant role as a defense by protecting plants from biotic factors such as herbivores, insects, and pathogens. In addition, they can enhance the resistance against abiotic factors.
1. Introduction
Plants established several adaptive mechanisms to endure abiotic factors, containing variations of metabolism in various directions, to confirm their existence in combative environmental situations
[1][66] (
Table 1). Several plant metabolites could assist and reduce the effect of the harsh stress of salt, drought, and water by acting as osmolytes and osmoprotectants
[2][67]. Examples of such metabolites include dimethylsulfoniopropionate (DMSP) and glycine betaine; sugars, such as sucrose, trehalose, and fructan; amino acids, such as proline and ectoine, as well as some metabolites of polyols, sorbitol, and mannitol
[3][4][68,69]. In plants, a wide range of waxy layers known as epicuticular wax keeps water balance during water shortage and acts as a mechanical stoppage to encounter disease-causing agents. Additionally, ascorbic acids, glutamine, alpha-tocopherol, anthocyanins, and carotene shield plant tissues by foraging the intermediates of bustling oxygen produced during oxidative stress
[5][70]. Similarly, several other smaller compounds guard plants against oxidation damage related to various constrictions
[6][65].
Besides, the plant’s defense system is related to generating phytoalexins, stimulating the common phenylpropanoid pathway and producing lignin biosynthesis
[7][71]. Further, phytochemicals and hormones such as salicylic acid and methyl salicylate, methyl jasmonate and jasmonic acid, as well as other small molecules formed due to stress, play a significant role against environmental stresses
[8][9][10][72,73,74]. All of these may also function as signaling compounds by stimulating the resistance system and reactions of acclimation
[11][75]. Among the defense systems of plants, osmotic regulation is one of the broadly pronounced responses to the water shortage that needs the accretion of harmonious solutes, such as sugars, amino acids, polyols, and glycine betaine
[12][76]. These chemical compounds do a significant job in sustaining cell turgor and stabilizing cell membranes and protein. Moreover, other studies designate the importance of these compounds in rehabilitating redox stability through the scavenging of ROS, which could adversely affect cellular structures and metabolism
[3][13][68,77].
2. Plant PriAmary Metabolites Produced in Response to Abiotic Stresino Acidses
2.1. Amino Acids
Amino acids are considered a precursor for protein and other organic molecules, e.g., nucleic acids, which designate an active part in the responses of a plant under several stress factors. Amino acids could also play a significant role in signaling and controlling molecules
[26][92]. Various studies showed that many amino acids stored in plants are apparent to different abiotic stresses
[27][28][93,94]. Moreover, the exposure of plants to such stresses appearance an accumulation of proline and other amino acids. In plants, the role played by stored amino acids differs after acting as an osmolyte to adjust ions passage, reducing stomatal opening and reclamation of
trace metals (TM
) [17][95]. Moreover, amino acids can also disturb the synthesis and activity of several enzymes, gene expression, and redox state of homeostasis
[14][96]. The accumulation of proline and ectoine is considered the most extensively dispersed osmolytes, as they act as osmoprotectants to protect plants from harmful effects and exciting environmental stresses, including low and high temperature, salinity,
ultraviolet radiations (UVr
), water, and osmotic stresses
[3][15][68,97].
Primarily, proline is produced from a glutamate and proline metabolizing enzyme, pyrroline-5-carboxylate synthetase (P5CS), which reduces glutamate to pyrroline-5-carboxylate (P5C). At last, from the reduction of P5C, this stress-responsive amino acid forms by pyrroline-5-carboxylate reductases (P5CR)
[29][98]. In transgenic plants, the significant role of proline was established during osmotic stress. For example, overexpression of the
P5CS gene in soybean increased proline content and, thus, tolerance to salt stress in transgenic plants
[30][99]. Besides osmolytes, proline is thought to accomplish many other important functions related to plant resistance, e.g., ROS scavenging, redox balancing, cytosolic pH buffer, molecular chaperon, and a stabilizer of protein structure
[29][98]. Subsequently, in response to abiotic factors, the enlarged levels of proline were observed for several years to be the stress-responsive feature in plants. The relationship between the accumulation of proline as osmolytes and stress tolerance had a great share because of its applicability to different crops
[31][32][100,101].
Remarkably, some of the metabolites were related to drought resistance and drought vulnerability of the considered hybrids
[33][102]. Additionally, studies on drought responses at metabolomics levels indicated that Andean potatoes with a phenotype designating greater stress exposure have more proline related to the genetically assembled plant that was a higher dearth-tolerant
[34][103]. It was established that the cultivar with a sensitive phenotype has high-level certain amino acids, containing proline and Gamma-aminobutyric acid (GABA) when barley exposed to salinity stress
[35][104]. It may well advocate a greater liability of these plants to such stress. According to
[14][96], this accretion could be associated with the deterioration of the leaf and slowing the development of a more subtle genotype. Furthermore, studies on Arabidopsis revealed that proline could be a lethal compound under heat stress
[36][105], while Charlton et al. found that water deficiency was the cause of the decrease in isoleucine concentration in
Pea and
Arabidopsis plants
[20][106].
2.2. Polyamines
3. Polyamines
Plants are tested by different stress factors and adversely affect their growth, yield, and geographical circulation
[37][107]. To survive the combative environmental stress circumstances, plants have developed many adaptive strategies, amongst which the accumulation of metabolites plays an important defensive role
[38][108]. Metabolites strongly involved in stress resistance are the low-molecular-weight (LMW) acyclic polyamines
[39][109]. Polyamines are the LMW nitrogen-containing organic compounds with more than two amino groups with a positive charge at the cellular pH, allowing them to link with negatively charged molecules, such as nucleic acids, phospholipids, and proteins
[40][110]. Usually, polyamines are polycations essential for plant growth and development and play an important role in abiotic stress resistance in higher plants. Triamine spermidine, tetraamine spermine, and their diamine predecessor, putrescine, are the general polyamines
[41][111]. Because of their cationic nature, these compounds have often been correlated to environmental stresses, such as drought, chilling, heat, TM, and salinity
[42][112].
The results of Khan et al.
[17][95] and Capell et al.
[43][113] showed that the accumulation of spermidine with the up-regulation of spermidine synthase of
Cucurbita ficifolia augmented several stress responses in a recombinant Arabidopsis plant, such as waterlogging and salinity stresses. It was shown that spermidine acts as a signaling molecule and controls the assertion of intricate genes in drought resistance. Furthermore, it has been demonstrated that polyamines are attributed to being involved in maintaining membranes shielding from damage under stressful environments
[44][114] and controlling the formation of nucleic acid as well as enzyme activity
[45][115]. Additionally, different findings revealed that polyamines play a significant role in oxidative stress by mitigating the balance state of ROS through their direct contact or indirectly regulating the antioxidant system and suppressing ROS production. Moreover, some authors hypothesized that polyamines could act as a cellular signal in plants throughout the stress responses
[46][116].
2.3. Carbohydrates
4. Carbohydrates
Carbohydrates produced during photosynthesis are the main building units that provide energy and support to the plant biomass
[47][117]. Extensive studies revealed that non-living factors lead to the assemblage of non-structural saccharides, such as sucrose and lactose, simple sugars, or polyhydric compounds (alcohols and phenols), amongst various species of plants
[48][118]. Particularly, there is a robust association between carbohydrate accretion and osmotic stress resistance, including oxidative stress (ROS) conditions, salt stress, and the scarcity of water
[17][95]. As a source of carbon and energy in a cell, soluble carbohydrates may take a significant part in the metabolic processes of plants. Several stress factors may impact the level of these soluble carbohydrates because the accumulation of carbohydrates is associated with photosynthesis
[49][119]. Rosa et al.
[50][120] demonstrated that certain soluble sugars, such as sucrose and hexoses, improved stress tolerance by down-regulating the stress-related genes and up-regulating growth-related genes. Though, the contents of certain carbohydrates, such as raffinose, glucose, fructose, and maltose, are highly sensitive to environmental stresses and increase. However, the contents of myoinositol were reduced in barley roots during water-scarce conditions
[51][121]. The findings of Sperdouli and Moustakas
[52][122] revealed an increase and contact of augmented soluble carbohydrates, sustaining a great antioxidant defense in the leaves of
Arabidopsis thaliana under dry environmental stress conditions. Studies showed renovation of carbon metabolism under salt-related stress (paraquat) in
A. thaliana tissues and inferred by the researchers as a substitute approach to staying alive
[52][122].
In water-deficit conditions, soluble sugars function as osmoprotectants, decreasing the harmful impact of osmotic stress and helps in sustaining the turgidity of cell and cell membrane stability by keeping plants from humiliation
[53][123]. Under stress conditions, the increase in sugar quantity is generally the result of carbohydrate hydrolysis that needs enzymes with hydrolytic usage
[18][124]. Moreover, carbohydrates that are soluble, such as disaccharides (sucrose and trehalose), oligosaccharides (raffinose and stachyose), and polymer of fructose molecules (fructans) next to their linked metabolic enzymes are essential compatible osmolytes associated with the scavenging of unstable molecules (ROS) during their assortment in plant tissues
[54][125]. In low-temperature stress, sugar alcohols, such as polyols, function as osmoprotectants and shield cell membranes against ice adhesion
[13][77]. Moreover, carbohydrates may act as signaling molecules
[55][126]. The demonstrated data advocate a specific response of carbohydrates in plants. However, it should be noted that the accumulation of carbohydrates depends on the kind of stress to which it bared
[56][127].
2.4. Glycine Betaine
5. Glycine Betaine
Glycine betaine (GB) is a widely studied quat compound, which is active in retaining the water balance between the plant cell and the environment during drought conditions. Moreover, GB playing a significant role in stabilizing the macromolecules, shielding photosynthesis, detoxification of reactive oxygen radicals, and as an osmoprotectant
[21][57][128,129]. Several studies indicated their importance in improving plant tolerance under various abiotic factors. It has been shown that plants are distinguished according to the formation of GB, such as barley, spinach, maize, and wheat, produce and accumulate a higher quantity of GB in their chloroplast. However, some plant species cannot obtain substantial amounts of GB during stress, such as
A. thaliana, rice, and tobacco
[58][130]. Furthermore, it has been shown that transgenic plants could mitigate the impact of abiotic stresses. Therefore, efforts have been made to improve tolerance through glycine betaine biosynthesis to achieve transgenic plants. In transgenic plants, such as
Arabidopsis, the
cyanobacteria genes, such as glycine sarcosine methyltransferase, and in transgenic maize, a greater amount of GB accumulates. As a result, in transgenic
Arabidopsis, resistance to drought and salt is greater; nevertheless, a recombinant plant of maize retained well in cold-related to non-transgenic cultivars
[59][60][131,132].
Moreover, through genetic engineering, other transgenic plants with a GB-producing capacity have been achieved, including
Brassica juncea and tobacco with greater tolerance to salt and chilling, indicating a progressive ability to propagate and grow well related to wild-type in abiotic environmental conditions
[61][62][133,134]. Besides, transgenic tomatoes with GB synthesis were more resistant to cold stress and produced fruit at a rate from 10 to 30% higher than the wild type.
[63][135]. Though, the meditations of GB produced in every transgenic plant were scarce to control the osmotic stress to which plants were exposed. Similarly, previous studies showed that GB could enhance root growth and reduce oxidative stress. Additionally, the exogenous application of GB improves the stress tolerance of Cr in chickpea plants
[22][136] and salinity stress in wheat
[64][137]. Consequently, further protecting approaches of GB, such as defense against ROS and heavy metals stress, should be considered, which may enhance the tolerance level
[23][138].
2.5. Lipids
6. Lipids
Lipids are a fundamental component of biological membranes, particularly the plasma membrane, which serves as the contact between the cell and its surroundings
[8][72]. Lipids can be grouped into eight major types based on the chemical structure in conjunction with distinctive hydrophobic and hydrophilic components, such as fatty acids, glycerides, phosphoglycerides, sphingolipids, steroids, isoprenoids, glycolipids, and polyketides
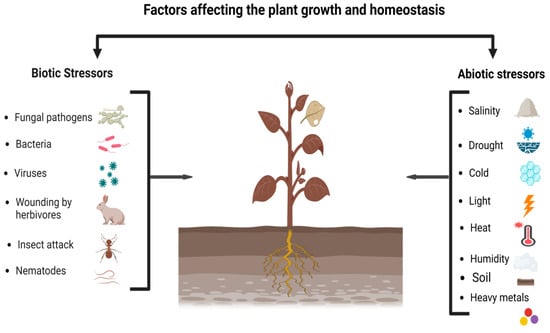
[65][139]. Being sessile organisms, plants are subjected to a wide variety of biotic and abiotic factors, such as temperature, drought, heavy metals, salinity, and pathogen attack. However, lipid-mediated signaling occurs in response to all these stressors (
Figure 1). The plasma membrane, which is typically the signaling source of lipids, is commonly used by plants to sense these stimuli and transform the signal into subsequent biochemical metabolism. Generally, these are acclimating enzymes that have all been proposed as signaling lipids, such as phospholipases, lipid kinases, and phosphatases
[66][140]. Commonly, lysophospholipid, fatty acid, phosphatides, triacylglycerol, inositol phosphate, oxylipins, sphingolipids, and nacylethanolamine are considered the major contributing signaling lipids molecules
[67][141]. The conformation and activity of cellular proteins and metabolites are influenced by signaling lipids because they have the ability to temporarily attract molecular markers to the membrane.
Figure 1. Environmental stresses of biotic and abiotic factors affecting the growth and homeostasis of plants.
Environmental stresses of biotic and abiotic factors affecting the growth and homeostasis of plants.
The enzyme phospholipase A (PLA) is very important in the formation of fatty acids and lysophospholipids. Usually, lysophospholipids are present in very limited amounts in plant tissues; however, in stressed conditions such as freezing their quantity increases
[68][142]. Some reports revealed the physiological role of lysophospholipids against various environmental stresses. Similarly, the phospholipase A2 (PlA2) has been shown to increase the production of some elicitors in poppy plants
[69][143], while lysophosphatidyl-choline and lysophosphatidyl-ethanolamine act as signals transducers in arbuscular symbiosis in potato
[70][144].
Fatty acids have also been demonstrated as stress-responsive lipids in plants. Oleic acids modulate nitric oxide-related proteins, thereby regulating nitric oxide and mitigating tolerance in
Arabidopsis [71][145]. Moreover, fatty acids also regulate drought, salt, and heavy metals tolerance, as well as the wound-induced responses of pathogens/herbivores in plants
[72][146]. Likewise, the responsive role of phosphatidic acid (PA), inositol polyphosphates, oxylipins, sphingolipids, and some other lipids have been studied in various plant species
[73][74][75][147,148,149]. Some of the environmental stress factors under which the plant lipid responses were reported to include chilling, freezing, and wounding
[76][150], pathogens
[24][151], low-temperature stress
[77][152], salt stress
[78][153], and water and drought
[25][154] stress response.