Diabetes is a metabolic disease characterized by a persistent over-normal level of blood glucose that causes impressive morbidity and mortality worldwide. Persistent hyperglycemia imposes damage on other organs, such as the eye, heart, kidney, and skin, as well as the nervous system, and is strongly correlated with a myriad of diabetes-related complications. Legumes, as an excellent source of protein, peptides, and phytochemicals, have played significant roles in human health throughout human history. Some legume-derived peptides with encouraging anti-diabetic potential have been gradually reported over the last two decades. Their hypoglycemic mechanisms have also been clarified at some classic diabetes treatment targets, such as the insulin receptor signaling pathway or other related pathways involved in the progress of diabetes, and key enzymes including α-amylase, α-glucosidase, and dipeptidyl peptidase-IV (DPP-4)
1. Introduction
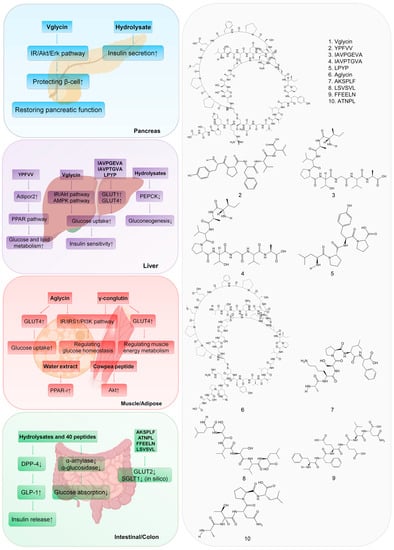
The hypoglycemic mechanism of legume diabetic peptides mainly includes reducing glucose absorption, promoting pancreatic β-cells proliferation, enhancing insulin secretion and sensitivity through signaling pathways associated with diabetes, and inhibiting carbohydrate-digesting enzymes (α-amylase and α-glucosidase) and dipeptidyl peptidase-IV (DPP-4) in target organs (Figure 1).
Figure 1. Mechanism of anti-diabetic effects of 10 representative legume-derived peptides. The amino acid sequence or molecular structure of the peptides are presented in the (right) panel. The anti-diabetic mechanisms of these peptides in corresponding organs are showed in the (left) panel. ↑ means a stimulative effect and ↓ means an inhibitory effect in the text. Peptide 1 works in pancreas; peptides 1 to 5 work in liver; peptide 6 works in muscle and adipose cells; and peptides 7–10 work in intestinal/colon tissue.
2. Targeting the Pancreas
One of the main factors contributing to the development of
type 2 diabetes (T2D
) is assumed to be a decrease in glucose-stimulated insulin secretion from pancreatic β-cells
[1][105]. An appealing idea for treating diabetes and its complications is to restore the function of poor β-cells. The IRS (insulin receptor substrate)/AKT (protein kinase B) pathway is the primary mechanism of the hypoglycemic effect of the body’s response to insulin stimulation
[2][106]. Vglycin peptide exhibited beneficial effects on hyperglycemia by facilitating glucose metabolism and enhancing insulin sensitivity, but it could not increase insulin secretion
[3][57]. Another study supports an apparent impact of vglycin on regulating β-cell preservation; a self-renew program was initiated in which vglycin directly promoted the proliferation of β-cells via activating the IR/Akt/Erk pathway
[4][14]. These results reveal the protective effects of vglycin on improving β-cell function in long-term glucolipotoxicity.
Some hydrolysates and fractions also exhibited a protective effect on the dysfunctional pancreas. Hydrolysates (<3 kDa) from hard-to-cook beans and common beans were reported to increase glucose-stimulated insulin secretion up to 57% and 45% from the basal state in INS-1E pancreatic β-cells, respectively
[5][6][66,76]. In another study, fermented meju water extracts (W-60), containing 15 kDa peptides, increased glucose-stimulated insulin secretion capacity and β-cell viability in Min6 insulinoma cells
[7][65]. Previous studies have identified that amino acid metabolism, especially glutamine and alanine, is essential for the functional maintenance of pancreatic cells and insulin secretion
[8][107]. Therefore, it can be inferred that the amino acids or oligopeptides in the hydrolysates of legumes may be beneficial for the exertion of hypoglycemic function of the pancreas directly or indirectly.
3. Targeting the Liver
Soymorphin-5 (YPFVV), derived from the soy conglycinin subunit, plays a role in glucose and lipid metabolism
[9][58]. Although no differences were observed in the weights of mesenteric and epididymal fat and brown adipose tissues, plasma TG levels and liver weights in KKA
y mice were significantly reduced by soymorphin-5 treatment, suggesting that soymorphin-5 enhanced the liver lipid metabolism. In terms of mechanism, soymorphin-5 improved fatty acid β-oxidation and energy expenditure by increasing the hepatic expression of PPAR and its target genes, including Adipor2, AOX1, CPT1, and UCP2
[9][58]. In another study, vglycin was probably responsible for increased fatty acid β-oxidation via the AMPK pathway and inhibited fatty acid synthesis (FAS) by down-regulating expression of FAS in HFD-fed C57BL/6J mice
[10][15]. Soy peptides IAVPGEVA, IAVPTGVA, and LPYP enhanced glucose uptake through GLUT1 and GLUT4 activation, and modulated glucose metabolism by activating Akt and AMPK pathways in HepG2 cells
[11][63]. In addition, γ-conglutin hydrolysates decreased gluconeogenesis by 50% in a dual-layered enterocyte/hepatocyte system and down-regulated the expression of phosphoenolpyruvate carboxykinase (PEPCK) in HepG2 cells
[12][64].
4. Targeting Muscle and Adipose Tissue
Increasing glucose absorption and transformation in muscle and adipose tissue is an effective method to control hyperglycemia. As shown in a
streptozotocin (STZ)/high-fat diet (HFD)STZ/HFD-induced diabetic mice model, oral administration of aglycin could potentially attenuate or prevent hyperglycemia via enhancing the IR/IRS1 signaling pathway in skeletal muscle
[13][56]. Additionally, in C2C12 cells, aglycin increased glucose uptake by recruiting glucose transporter GLUT4 to the surface of cells
[13][56]. Cowpea peptides induced the Akt phosphorylation in skeletal cells and mimicked the actions of insulin by activating the same signaling pathway
[14][67]. Prior findings strongly implied that the translocation of GLUT4 was regulated by activation of the Akt signaling pathway in the skeletal muscle cells, which facilitated glucose uptake
[15][16][108,109].
Studies of fermented soy products have suggested that the small bioactive peptides produced during fermentation may contribute to preventing or slowing the progression of T2D
[7][17][65,110]. Water extracts (<3 kDa) from chungkookjang increased PPAR-r activity by about 59%, indicating that there exist certain small peptides that work as mild PPAR-r agonists in glucose and lipid metabolism
[7][65]. γ-conglutin hydrolysates also significantly increased the insulin-induced glucose uptake in mature 3T3-L1 adipocytes through an enhanced expression of GLUT4, similar to metformin treatment
[12][64]. Similarly, in C2C12 cells, γ-conglutin increased flottilin-2, caveolin-3 concentrations, and CBL phosphorylation, resulting in GLUT4 translocated on the cell membrane, which suggested that γ-conglutin plays an essential role in muscle energy metabolism through insulin-mimetic action
[18][73].
5. Targeting the Intestine and Colon
In STZ-induced Wistar rats and the Caco-2 cell models, black bean peptides AKSPLF, ATNPL, FFEELN, and LSVSVL may have reduced glucose absorption via blocking glucose transporters GLUT2 and SGLT1 (
Figure 1)
[19][60]. The mechanism of lowering postprandial hyperglycemia by inhibition of α-amylase and α-glucosidase is to inhibit the rate of glucose release and absorption in the intestine
[20][12]. The hypoglycemic mechanism of inhibiting the DPP-4 enzyme is to retard the decomposition of GLP-1 and increase the endogenous physiological level of GLP-1 in the human body
[21][34]. Recently, the structure of oligopeptides and their binding modes to these enzymes have been systematically investigated, which may provide a clear anti-diabetic mechanism for these peptides
[22][40].
α-amylase contains three distinct structural domains (A, B, and C). The active site region in domain A is associated with an extended substrate binding cleft, allowing it to recognize polymeric starch molecules
[23][111]. Asp197, Glu233, and Asp300 are three essential catalytic carboxylic acid residues located within this active site cleft between the carboxyl ends of domains A and B. Domain B is responsible for substrate specificity and enzyme stability, and domain C helps to maintain the stability of the catalytic site. Therefore, blocking the catalytic sites was an effective way to inhibit the activity of α-amylase
[22][40]. Peptides amply containing aromatic residues such as Leu, Pro, Gly, and Phe were effective inhibitors of α-amylase because the substrate-binding pockets of the α-amylase enzyme have many aromatic residues
[23][111]. Hence, apart from interactions involving hydrogen bonds and electrostatic and van der Waals interactions, aromatic–aromatic interactions have also played a crucial role in α-amylase inhibitory activity
[24][4].
The bioactive peptides LSSLEMGSLGALFVCM, PLPLHMLP, PPMHLP, PPHMLP, PPHMGGP, and PLPWGAGF, extracted from pinto beans, interacted or bound to the indicated domain of α-amylase, ultimately restricting protein conformation and affecting the degree of enclosure of the substrate. Mechanically, when the (α-amylase)–starch complex was formed, the uncompetitive peptide inhibitors bound to the complex, resulting in the starch no longer being able to hold on at its original position due to the altered conformation of α-amylase. Consequently, the starch was detached from the (α-amylase)–starch complex
[25][92]. The competitive inhibitors GSR and EAK formed hydrogen bonds with the catalytic residues
[26][62].
Structural analysis has clearly shown the intimate interaction between bean-derived peptides and α-amylase catalytic residues. The interaction prevented residue Asp300 from adopting its functional position and destroyed the water channel leading from the “flexible loop” to the heart of the active-site depression
[27][112]. Another inhibitory mechanism was identified in the interaction of porcine pancreatic α-amylase and the bean P. vulgaris-derived peptide, in which residue His305 participated in substrate binding and structural changes
[28][113].
Similar to peptides that inhibit α-amylase, certain peptides have the potential to inhibit α-glucosidase, reducing glucose absorption and delaying carbohydrate digestion, thereby reducing postprandial blood glucose levels. Pure peptides such as KTYGL were able to inhibit the enzyme mainly through polar interactions (Asn32, Asp34, Asp38, and Asp89), hydrophobic interactions (Trp36 and Trp81), and hydrogen bonds (Ser31) with the residues from the binding pocket of the α-glucosidase
[29][98]. AKSPLF showed only hydrogen bonding and polar interactions with α-glucosidase, and Ala and Lys were the only two amino acids interacting with ASP34, THR83, ASP89 and ASN32
[30][95]. Besides, α-glucosidase inhibitors also played crucial roles in the secretion of GLP-1 in T2D
[31][114].
DPP-4 is a serine exopeptidase that cleaves X-proline or X-alanine dipeptides from the N-terminus of polypeptides, which degrades and inactivates GLP-1 and GIP. Several legume-derived peptides have been identified as promising DPP-4 inhibitors, potentially contributing to glycemic control. DPP-4 has hydrophobic pockets, which are crucial targets for inhibition of the enzyme
[32][115]. Generally, the DPP-4 inhibitory peptides often contain high concentrations of hydrophobic amino acids such as alanine, glycine, isoleucine, leucine, phenylalanine, proline, methionine, tryptophan, and valine
[24][4]. The interactions between DPP-4 and common bean peptides (KTYGL and AKSPLF) were mainly H-bond, hydrophobic, polar and cation π bonds
[25][33][85,92]. The interactions of DPP-4 with peptide AKSPLF occurred primarily between amino acids GLU191, ASP192, ARG253, and LEU235
[30][95].
Collectively, legume peptides exhibit great inhibitory potential due to their affinity for and specificity of action on key enzymes such as α-amylase, α-glucosidase, and DPP-4, which are promising candidates for the development of anti-diabetic drugs. However, the structure–activity relationships between peptides and these enzymes remain to be uncovered.