Abiotic stress in plants is one of the main obstacles to global agricultural production and food security. Therefore, there is a need for the development of novel approaches to overcome these problems and achieve sustainability. Nanotechnology has emerged as one such novel approach to improve crop production, through the utilization of nanoscale products, such as nanofertilizer, nanofungicides, nanoherbicides and nanopesticides. Their ability to cross cellular barriers makes nanoparticles suitable for their application in agriculture. Since they are easily soluble, smaller, and effective for uptake by plants, nanoparticles are widely used as a modern agricultural tool. The implementation of nanoparticles has been found to be effective in improving the qualitative and quantitative aspects of crop production under various biotic and abiotic stress conditions.
- climate changes
- abiotic stress
- nanoparticles
- molecular changes
1. Nanoparticles in Salt-Stress Tolerance
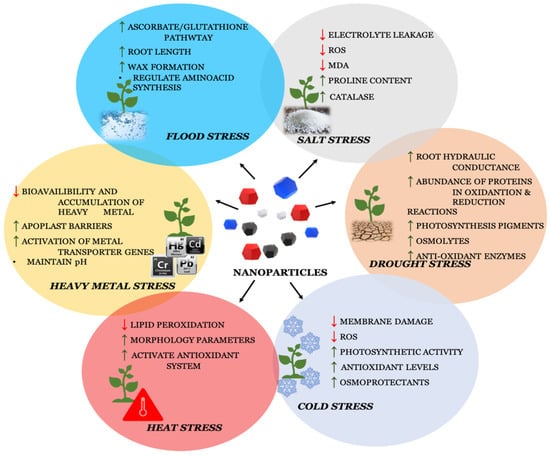
2. Nanoparticles in Drought-Stress Tolerance
3. Nanoparticles in Cold-Stress Tolerance
4. Nanoparticles in Heavy-Metal-Stress Tolerance
5. Nanoparticles in Flooding-Stress Tolerance
6. Nanoparticles in Heat-Stress Tolerance
References
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80.
- Mahmood, R.; Ijaz, M.; Qamar, S.; Bukhari, S.A.; Malik, K. Abiotic stress signaling in rice crop. In Advances in Rice Research for Abiotic Stress Tolerance; Mirza, H., Masayuku, F., Kamrun, N., Jiban, B., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 551–569.
- Mohamed, H.I.; Sajyan, T.K.; Shaalan, R.; Bejjani, R.; Sassine, Y.N.; Basit, A. Plant-mediated copper nanoparticles for agri-ecosystem applications. In Agri-Waste and Microbes for Production of Sustainable Nanomaterials; Kamel, A.A., Rajiv, P., Rajeshkumar, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 79–120.
- Dong, F.; Yang, F.; Liu, Y.; Jia, W.; He, X.; Chai, J.; Zhao, H.; Lv, M.; Zhao, L.; Zhou, S. Calmodulin-binding transcription activator (CAMTA)/factors in plants. In Calcium Transport Elements in Plants; Academic Press: Cambridge, MA, USA, 2021; pp. 249–266.
- Sinha, R.K.; Verma, S.S. Proteomics approach in horticultural crops for abiotic-stress tolerance. In Stress Tolerance in Horticultural Crops; Woodhead Publishing: Sawston, UK, 2021; pp. 371–385.
- Zulfiqar, F.; Ashraf, M. Nanoparticles potentially mediate salt stress tolerance in plants. Plant Physiol. Biochem. 2021, 160, 257–268.
- El-Sharkawy, M.S.; El-Beshsbeshy, T.R.; Mahmoud, E.K.; Abdelkader, N.I.; Al-Shal, R.M.; Missaoui, A.M. Response of Alfalfa under Salt Stress to the Application of Potassium Sulfate Nanoparticles. Am. J. Plant Sci. 2017, 8, 1751–1773.
- Khan, I.; Raza, M.A.; Awan, S.A.; Shah, G.A.; Rizwan, M.; Ali, B.; Tariq, R.; Hassan, M.J.; Alyemeni, M.N.; Brestic, M.; et al. Amelioration of salt induced toxicity in pearl millet by seed priming with silver nanoparticles (AgNPs): The oxidative damage, antioxidant enzymes and ions uptake are major determinants of salt tolerant capacity. Plant Physiol. Biochem. 2020, 156, 221–232.
- Rossi, L.; Zhang, W.; Ma, X. Cerium oxide nanoparticles alter the salt stress tolerance of Brassica napus L. by modifying the formation of root apoplastic barriers. Environ. Pollut. 2017, 229, 132–138.
- Ozturk, M.; Unal, B.T.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2020, 172, 1321–1335.
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33.
- Zhang, H.; Zhao, Y.; Zhu, J.-K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Dev. Cell 2020, 55, 529–543.
- Kandhol, N.; Jain, M.; Tripathi, D.K. Nanoparticles as potential hallmarks of drought stress tolerance in plants. Physiol. Plant. 2022, 174, e13665.
- Alabdallah, N.M.; Hasan, M.; Hammami, I.; Alghamdi, A.I.; Alshehri, D.; Alatawi, H.A. Green Synthesized Metal Oxide Nanoparticles Mediate Growth Regulation and Physiology of Crop Plants under Drought Stress. Plants 2021, 10, 1730.
- Zahedi, S.M.; Hosseini, M.S.; Meybodi, N.D.H.; Peijnenburg, W. Mitigation of the effect of drought on growth and yield of pomegranates by foliar spraying of different sizes of selenium nanoparticles. J. Sci. Food Agric. 2021, 101, 5202–5213.
- El-Zohri, M.; Al-Wadaani, N.A.; Bafeel, S.O. Foliar Sprayed Green Zinc Oxide Nanoparticles Mitigate Drought-Induced Oxidative Stress in Tomato. Plants 2021, 10, 2400.
- Bishta, S.; Sharma, V.; Kumari, N. Biosynthesized magnetite nanoparticles from Polyalthia longifolia leaves improve photosynthetic performance and yield of Trigonella foenum-graecum under drought stress. Plant Stress 2022, 5, 100090.
- Potter, M.; Deakin, J.; Cartwright, A.; Hortin, J.; Sparks, D.; Anderson, A.J.; McLean, J.E.; Jacobson, A.; Britt, D.W. Absence of Nanoparticle-Induced Drought Tolerance in Nutrient Sufficient Wheat Seedlings. Environ. Sci. Technol. 2021, 55, 13541–13550.
- Aslam, M.; Fakher, B.; Ashraf, M.A.; Cheng, Y.; Wang, B.; Qin, Y. Plant Low-Temperature Stress: Signaling and Response. Agronomy 2022, 12, 702.
- Adhikari, L.; Baral, R.; Paudel, D.R.; Min, D.; Makaju, S.O.; Poudel, H.P.; Acharya, J.P.; Missaoui, A.M. Cold stress in plants: Strategies to improve cold tolerance in forage species. Plant Stress 2022, 4, 100081.
- Mohammadi, R.; Maali-Amiri, R.; Abbasi, A. Effect of TiO2 Nanoparticles on Chickpea Response to Cold Stress. Biol. Trace Elem. Res. 2013, 152, 403–410.
- Mohammadi, R.; Maali-Amiri, R.; Mantri, N. Effect of TiO2 nanoparticles on oxidative damage and antioxidant defense systems in chickpea seedlings during cold stress. Russ. J. Plant Physiol. 2014, 61, 768–775.
- Amini, S.; Maali-Amiri, R.; Mohammadi, R.; Shahandashti, S.-S.K. cDNA-AFLP Analysis of Transcripts Induced in Chickpea Plants by TiO2 Nanoparticles during Cold Stress. Plant Physiol. Biochem. 2017, 111, 39–49.
- Hasanpour, H.; Maali-Amir, R.; Zeinali, H. Effect of TiO2 nanoparticles on metabolic limitations to photosynthesis under cold in chickpea. Russ. J. Plant Physiol. 2015, 62, 779–787.
- Ghabel, V.K.; Karamian, R. Effects of TiO2 nanoparticles and spermine on antioxidant responses of Glycyrrhiza glabra L. to cold stress. Acta Bot. Croat. 2020, 79, 137–147.
- Wang, A.; Li, J.; Al-Huqail, A.A.; Al-Harbi, M.S.; Ali, E.F.; Wang, J.; Ding, Z.; Rekaby, S.A.; Ghoneim, A.M.; Eissa, M.A. Mechanisms of Chitosan Nanoparticles in the Regulation of Cold Stress Resistance in Banana Plants. Nanomaterials 2021, 11, 2670.
- Song, Y.; Jiang, M.; Zhang, H.; Li, R. Zinc Oxide Nanoparticles Alleviate Chilling Stress in Rice (Oryza Sativa L.) by Regulating Antioxidative System and Chilling Response Transcription Factors. Molecules 2021, 26, 2196.
- Elsheery, N.I.; Sunoj, V.; Wen, Y.; Zhu, J.; Muralidharan, G.; Cao, K. Foliar application of nanoparticles mitigates the chilling effect on photosynthesis and photoprotection in sugarcane. Plant Physiol. Biochem. 2020, 149, 50–60.
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy Metal Stress and Some Mechanisms of Plant Defense Response. Sci. World J. 2015, 2015, 756120.
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179.
- Tiwari, S.; Lata, C. Heavy Metal Stress, Signaling, and Tolerance Due to Plant-Associated Microbes: An Overview. Front. Plant Sci. 2018, 9, 452.
- Wang, Y.; Liu, Y.; Zhan, W.; Zheng, K.; Lian, M.; Zhang, C.; Ruan, X.; Li, T. Long-term stabilization of Cd in agricultural soil using mercapto-functionalized nano-silica (MPTS/nano-silica): A three-year field study. Ecotoxicol. Environ. Saf. 2020, 197, 110600.
- Cui, H.; Shi, Y.; Zhou, J.; Chu, H.; Cang, L.; Zhou, D. Effect of different grain sizes of hydroxyapatite on soil heavy metal bioavailability and microbial community composition. Agric. Ecosyst. Environ. 2018, 267, 165–173.
- Wang, K.; Wang, Y.; Wan, Y.; Mi, Z.; Wang, Q.; Wang, Q.; Li, H. The fate of arsenic in rice plants (Oryza sativa L.): Influence of different forms of selenium. Chemosphere 2021, 264, 128417.
- Cui, J.; Liu, T.; Li, F.; Yi, J.; Liu, C.; Yu, H. Silica nanoparticles alleviate cadmium toxicity in rice cells: Mechanisms and size effects. Environ. Pollut. 2017, 228, 363–369.
- Wu, H.; Tito, N.; Giraldo, J.P. Anionic Cerium Oxide Nanoparticles Protect Plant Photosynthesis from Abiotic Stress by Scavenging Reactive Oxygen Species. ACS Nano 2017, 11, 11283–11297.
- Bailey-Serres, J.; Colmer, T.D. Plant tolerance of flooding stress—Recent advances. Plant Cell Environ. 2014, 37, 2211–2215.
- Mustafa, G.; Sakata, K.; Komatsu, S. Proteomic analysis of flooded soybean root exposed to aluminum oxide nanoparticles. J. Proteom. 2015, 128, 280–297.
- Mustafa, G.; Sakata, K.; Komatsu, S. Proteomic analysis of soybean root exposed to varying sizes of silver nanoparticles under flooding stress. J. Proteom. 2016, 148, 113–125.
- Mustafa, G.; Komatsu, S. Insights into the response of soybean mitochondrial proteins to various sizes of aluminum oxide nanoparticles under flooding stress. J. Proteome Res. 2016, 15, 4464–4475.
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and responses of chloroplasts to heat stress in plants. Front. Plant Sci. 2020, 11, 375.
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273.
- Zhao, J.; Lu, Z.; Wang, L.; Jin, B. Plant responses to heat stress: Physiology, transcription, noncoding RNAs, and epigenetics. Int. J. Mol. Sci. 2020, 22, 117.
- Djanaguiraman, M.; Belliraj, N.; Bossmann, S.H.; Prasad, P.V.V. High-Temperature stress alleviation by selenium nanoparticle Treatment in Grain Sorghum. ACS Omega 2018, 3, 2479–2491.
- Iqbal, M.; Raja, N.I.; Mashwani, Z.-U.; Hussain, M.; Ejaz, M.; Yasmeen, F. Effect of silver nanoparticles on growth of wheat Under Heat Stress. Iran. J. Sci. Technol. Trans. A Sci. 2019, 43, 387–395.
- Hassan, N.S.; El Din, T.A.S.; Hendawey, M.H.; Borai, I.H.; Mahdi, A.A. Magnetite and zinc oxide nanoparticles alleviated heat stress in wheat plants. Curr. Nanomater. 2018, 3, 32–43.
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon Regulates Antioxidant Activities of Crop Plants under Abiotic-Induced Oxidative Stress: A Review. Front. Plant Sci. 2017, 8, 510.