Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Sirius Huang and Version 1 by Loredana Raciti.
Consciousness is a multifaceted concept, involving both wakefulness, i.e., a condition of being alert that is regulated by the brainstem, and awareness, a subjective experience of any thoughts or perception or emotion. Disorders of consciousness (DOC) are usually caused by a severe acquired brain injury that leads to a loss of consciousness lasting at least 24 h. The management of sleep disorders in DOC patients is an increasingly hot topic and deserves careful diagnosis, to allow for the most accurate prognosis and the best medical treatment possible.
- disorders of consciousness
- sleep
- UWS and MCS
- wake-sleep cycle
1. General Characteristics of Sleep Pattern
Sleep is studied worldwide by researchers and clinicians using PSG, which includes the study of EEG, electromyography, electrooculography, electrocardiography, pulse oximetry, airflow, and respiratory strength [30][1]. Normally, sleep is divided into two stages, as first defined by Rechtschaffen and Kales in 1968 [31][2], and later modified by the American Association of Sleep Medicine [32][3]: NREM sleep and REM sleep. These stages are further divided into cycles that range from stage 1 to stage 3 of NREM (N1–N2–N3), ending in REM sleep and wakefulness (W) with a duration of 90–110 min/cycle; and each stage lasting 5–15 min. Every stage is characterized by a specific EEG pattern [33][4]: delta waves and, sometimes, spindles in the N3 stage (the deepest stage of sleep); spindle and K-complex in the N2 stage; theta waves in the N1 stage; and theta waves with sawtooth waves and rapid eye movements in the REM stage; meanwhile alpha waves appear in the W stage (Figure 1).
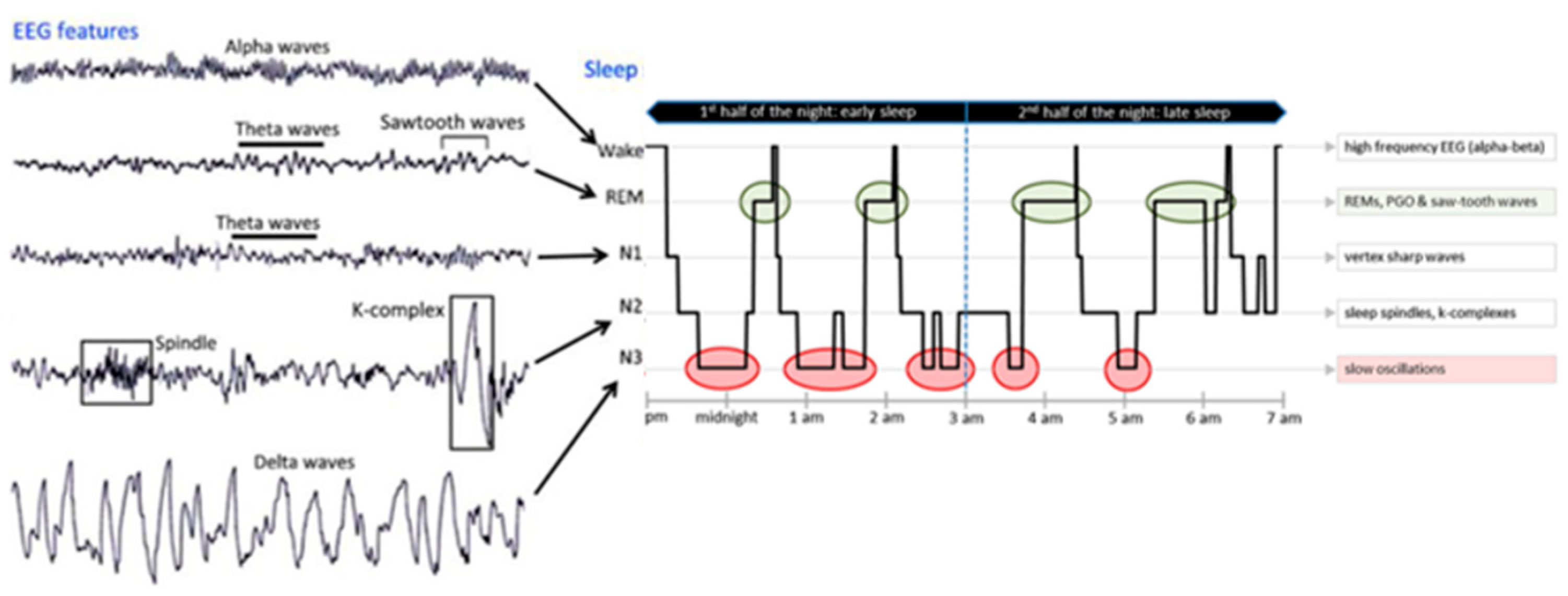
Figure 1.
EEG pattern (on the
left
) and an 8 h recorded hypnogram (on the
right
) of normal sleep stages.
Recently, because of the time-consuming nature of the analysis of sleep recordings, automatic sleep stage classification has used depth network structures for sleep stage scoring, such as convolutional neural networks [34][5] and recurrent neural networks (RNNs) [35][6]. More recently, a method based on ICA-Relief-F has been proposed for the Sleep-EDF database [36,37][7][8]. However, a standard approach and procedure is still not available, to allow comparing the results of different studies. For these reasons, most of the studies in the field rely on the visual inspection of PSG recordings, which has severe limitations in terms of reproducibility and cost [18][9]. Normally, an EEG is characterized by more than 100 standard spindles and a full sleep–wave cycle. To better study the sleep stages, the time of sleep is conventionally divided into epochs, a continuous period of 30 s.
1.1. Anatomy and Physiology of the Sleep–Wake Cycle
The neuronal system that regulates the sleep–wake cycles is extended from the medulla, the brainstem, and hypothalamus up into the basal forebrain, which allows the fusing of the sleep–wake cells. The hypothalamus regulates sleep and wakefulness through the ventrolateral preoptic nucleus (VLPO), facilitated by the gamma-aminobutyric acid (GABA) and galanin neurons regulating normal sleep. In addition, the posterior lateral hypothalamus is facilitated by orexin/hypocretin neurons that regulate wakefulness, inhibiting the VLPO neurons [38][10].
Several neurotransmitters are released during the sleep–wake cycle, such as catecholamines, serotonin, and acetylcholine. Moreover, several factors specifically influence the wakefulness, such as olfactory, visual, vestibular, and other neuromodulators. [39][11]. The reticular activating system (RAS) has been shown to be the arousal system maintaining consciousness. The RAS, after receiving the impulses from the spinal cord, conveys them to the thalamus and successively to the cerebral cortex [39][11]. Wakefulness is promoted by the basal forebrain, lateral hypothalamus, and tuberomammillary nucleus. In particular, the neurons in the basal forebrain containing acetylcholine promote arousal involvement of the cerebral cortex during the W phase of sleep, activating alertness [40,41][12][13]. Accordingly, lesions in this area with the neurotoxin hypocretin-2-saporin cause severe loss of sleep [41][13]. On the other hand, the lateral hypothalamus neurons are defined as a “wake switch” for the connections to several wake-promoting areas, such as to the adrenergic, histaminergic, dopaminergic, and cholinergic nuclei [42][14]. According to the studies of Constantine von Economo, the anterior hypothalamus is associated with sleep, whereas the posterior hypothalamus is correlated to awareness of the histaminergic neurons [43,44][15][16] that project into the cerebral cortex, the amygdala, and the substantia nigra, and receive input from the hypocretinergic neurons of the lateral hypothalamus and GABAergic neurons in the ventrolateral preoptic area [45,46][17][18]. The brainstem sends wake-promoting neurons from the rostral reticular formation to the forebrain, regulating the sleep–wake cycle. The neurons of the rostral pons that project into the dorsal thalamic nuclei via a dorsal pathway, fire rapidly during W and REM sleep, but decrease during SWS, and are called W-on/REM-on neurons. Another pathway is the ventral pathway that projects into the magnocellular neurons in the substantia innominata, medial septum, and the diagonal band [47][19], actively firing during W and becoming inactive during SWS and REM sleep. As already reported, the wake phase is activated by various arousal systems that generate different neurochemicals [48][20], such as histamine- [43][15], glutamate [49][21], noradrenaline (NA)- [50[22][23][24],51,52], dopamine (DA)- [48[20][25],53], 5-hydroxytryptamine (5-HT)- [54[26][27],55], ACh- [56][28], and hypocretin (HCRT)-containing neurons [57,58][29][30] (Figure 2).

Figure 2. Structures and neurotransmitters responsible for arousal and sleep. In the figure: regulation of the ascending arousal system by the hypothalamic. Red lines represent the pathway of activation of the cortex, to enable the handling of inputs from the thalamus. The monoaminergic structures involved in this process are the locus coeruleus (LC), by noradrenaline (NA); the dorsal and median raphe nuclei, by serotonin (5-HT); the A10 cell group, by dopamine (DA); and the tuberomammillary nucleus (TMN), by histamine (His). Moreover, peptidergic structures are involved, such as the lateral hypothalamus (LHA), by orexin (ORX), or melanin-concentrating hormone (MCH) and basal forebrain (BF) neurons that contain γ-aminobutyric acid (GABA) or acetylcholine (Ach). The orange lane represents the input to the thalamus from cholinergic (ACh) structures, such as the upper pons, the pedunculopontine (PPT), and laterodorsal tegmental nuclei (LDT).
1.2. Sleep
Sleep is activated by several structures: the suprachiasmatic nucleus, the ventrolateral preoptic nucleus (VLPO), and the basal forebrain and brainstem. At the suprachiasmatic nucleus level, sleep is modulated by two mechanisms: the homeostatic and the circadian. The homeostatic mechanism is characterized by an optimal sleep duration and intensity in a short period, which depends on the individual’s history of sleep–waking. For example, a subject deprived of sleep will show an increased number of SWSs and the recovery sleep will be characterized by a longer period of sleep and delta wave activity [59][31]. The circadian mechanism is controlled by the suprachiasmatic nucleus (SCN) of the hypothalamus, defined as the “master clock” of sleep, because it rearranges and coordinates the circadian rhythms of the peripheral tissues. Impairment of the SCN causes a loss of W and sleep establishment [60,61,62][32][33][34]. The anterior hypothalamus of the VLPO nucleus is defined as the “sleep-generating” center [63,64][35][36] because of its two nuclei, the VLPO nucleus associated with SWS, and the extended VLPO nucleus, linked to REM sleep generation [65,66][37][38] and activating the inhibitory transmitters GABA and galanin that project into the arousal neurons in the hypothalamus and the brainstem [59][31]. Stimulus of the basal forebrain generated sleep [67[39][40],68], inhibiting the TMN nucleus [69][41]. Meanwhile the brainstem sends an input to the thalamic and reticular nucleus through the pedunculopontine tegmental nuclei and the latero-dorsal tegmental nuclei [70[42][43],71], whose neurons fire during REM sleep. All these important neurobiological mechanisms can be damaged by brain injury, and this damage may account for the abnormal wake–sleep cycle in DOC patients.
2. Characteristics and Assessment of Sleep in UWS/MCS
Sleep characteristics in DOC patients are still controversial and not well studied, due to various factors, such as the etiology of brain damage, the kind of brain dysfunction, and the related sleep abnormalities. Severe impairment of circadian rhythms has been shown [22[44][45][46][47],72,73,74], and the interruption of the regular interval of sleep and wakefulness is possibly due to injury of the brain stem [75][48] or cortical dysfunctions such as in the diffuse axonal damage or gray matter abnormalities due to brain hypoxia. Consequently, it was demonstrated that normal sleep was reduced at night in MCS patients with a predominance of daytime sleepiness compared to control patients. On the contrary, UWS patients showed a more severe impairment of sleep cycle with sleep phases present only during the day [76][49]. It could be then hypothesized that DOCs are characterized by dysregulations involving excitatory wake-promoting and inhibitory sleep-promoting neurotransmitters, which are involved in maintaining efficient connections between high-order cortical areas and sub-cortical structures [77][50]. A reduction of both integration and segregation was empirically described when an alteration of the diurnal sleep–wake cycle intervened. [78,79,80][51][52][53]. Due to the non-conventional sleep pattern of DOC patients, the use of approved terminology and scores is sometimes not applicable and suitable. Therefore, further studies are necessary to identify specific criteria for sleep and the EEG/PSG pattern in DOC patients.
Usually, the sleep–wake cycle of DOC patients is influenced by various environmental elements, such as light, bronco aspirations, hygiene, and nocturnal enteral or parenteral therapy. All these factors can cause sleep fragmentation with frequent arousals and awakenings, and this can interfere with PSG registration and sleep assessment. Moreover, other factors such as brain neurosurgery interventions, hypothermia, skin lesions or medical advice, and recovery elements can cause interference in PSG recordings. For these reasons, an independent component analysis using the Wiener and Kalman filtering wavelet analysis has been suggested to remove the myogenic artefacts of UWS patients [81][54]. To achieve, better results, PSG must be recorded for 48–72 h to obtain EEG data for multimodal analysis and independent component analysis algorithms, due to excessive daytime sleepiness and lack of sleep during the night. [82,83][55][56]. The sleep patterns of K-complexes, slow oscillations, and REM-sleep have been described as potential prognostic factors in this patient population in [21,84,85][57][58][59]. Usually, DOC patients show altered sleep phases that are clearly abnormal, especially in UWS with respect to MCS patients [86][60]. In particular, it has been shown that in coma patients the sleep–wake cycle and spindles are absent. Spindles and an irregular sleep–wake cycle are rarely observed in UWS; meanwhile, they are present, even if irregular, in MCS patients, as well as in the NREM2 and slow-wave sleep (SWS) stages [87][61].
The presence of spindle waves shows the functional integrity of the thalamus. The appearance of slow-wave and REM sleep is a good outcome for the function of the brainstem nuclei, whereas the hypothalamic functional integrity is expressed by the circadian organization of sleep patterns [18][9]. The sleep–wake cycle and circadian rhythm [84,87][58][61] are also normally regulated by the levels of melatonin and orexin [88,89,90][62][63][64]. Melatonin is secreted from the pineal gland, and it only has a circadian rhythm with high levels during the late evening [91,92][65][66]; meanwhile, orexin neurons secreted by the lateral hypothalamic area and posterior hypothalamus [93][67] are secreted during wakefulness and silent during non-rapid eye movement (NREM) and rapid eye movement (REM) sleep [57][29]. Therefore, it was shown that improving the restoration of melatonin metabolism was related to better cognitive function, evaluated using the coma recovery scale-revised (CRS-R) score in patients with brain injury [83,94,95][56][68][69].
At the same time, regular secretion of urinary 6-sulphatoxymelatonin and free cortisol was linked to an upgrading of cognition and awareness in comatose patients. However, Yang et al. were not able to show any involvement of the pineal gland in the retention of awareness nor a correlation between levels of orexin and recovery of consciousness [94][68]. In a recent study by Mertel et al. on the sleep patterns of 32 DOC patients, the authors found that MCS patients had a sleep pattern like control subjects. Specifically, in contrast with UWS, MCS and control subjects slept more during the night than during the day. Regarding REM stage, 100% of control subjects and 88% MCS patents presented REM sleep, whereas 44% of UWS did not have REM sleep. On the other hand, spindles were absent in 62% of UWS and 21% of MCS, without any REM sleep, and with spindles in 12% and 21%, respectively. Otherwise, the UWS patients did not show a night–day sleep circle and 44% and 62% did not show REM sleep and sleep spindles. On the other hand, the number of SWS was practically the same in all groups [76][49].
The reappearance of the eye-opening periods, of the circadian rhythm, and of the behavioral sleep–wake cycle represent signals of a return of awareness and arousal from a coma state to MCS [96][70]. However, if significant damage of the cerebral cortex exists, such as a loss of the cortical neurons or a reduction in the number of interneuron connections in the cerebral cortex, axonal damage with degeneration of cortical neurons, or a reduction in the activity of the caudal group of the nuclei of the basal forebrain, the PSG is insufficient. Loss of cortical neurons and their connections is converted into a slowing down in cortical rhythm and the absence of high-frequency activity of subcortical structures and synchronized postsynaptic excitatory potentials of cortical neurons [97][71]. Thus, sleep spindles are absent in EEG recordings. Therefore, the presence of spindles designates a preserved cerebral cortex and a favorable prognosis [98][72]. Spindles represent the preservation of thalamocortical connectivity. Therefore, they have been considered as a predicted sign for plasticity and of early behavioral awakening in comatose patients [99][73] and when absent, a poor outcome should be considered [100][74].
Regarding the REM phase, even though it has been shown that the REM phase in UWS patients was significantly reduced, there was no correlation between the frequency of these activities and recovery of the clinical condition. [101][75]. Moreover, an impaired REM phase is a signal of compromised brain stem mechanisms. In one-third of UWS patients and in more than half of MCS patients, preserved sleep spindles, rapid eye movement, and slow-wave sleep with favorable outcomes were shown, but only when a high quality and quantity of sleep standard spindles were present [18,84][9][58]. To allow the best neurological prognosis, the EEG must be scored based on American Clinical Neurophysiology Society (ACNS) terminology [102][76]. The ACNS takes into consideration several EEG characteristics, such as time domain or frequency domain, that provided effective and comprehensible and applicable indices to describe brain electrical activities under different states of consciousness. Therefore, a standard EEG recording classified according to the ACNS terminology may improve the capability for neurological prognosis [22,102][44][76]. Recently, Wielek et al. investigated a multivariate machine learning technique to categorize sleep/wake stages in DOC patients, which may represent an alternative to the standard PSG [103][77].
References
- Rundo, J.V.; Downey, R. Polysomnography. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 160, pp. 381–392. ISBN 978-0-444-64032-1.
- Rechtschaffen, A.; Kales, A. A Manual of Standardized Terminology, Techniques and Scoring System for Sleep Stages in Human Subjects; National Institutes of Health Publications, US Government Printing Office: Washington, DC, USA, 1968.
- Iber, C.; Ancoli, I.-S.; Andrew, L.; Stuart, F. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications; American Academy of Sleep Medicine: Westchester, IL, USA, 2007.
- Fiorillo, L.; Fiorillo, L.; Puiatti, A.; Papandrea, M.; Ratti, P.-L.; Favaro, P.; Roth, C.; Bargiotas, P.; Bassetti, C.L.; Faraci, F.D. Automated sleep scoring: A review of the latest approaches. Sleep Med. Rev. 2019, 48, 101204.
- Mousavi, S.; Afghah, F.; Acharya, U.R. SleepEEGNet: Automated sleep stage scoring with sequence to sequence deep learning approach. PLoS ONE 2019, 14, e0216456.
- Phan, H.; Andreotti, F.; Cooray, N.; Chéen, O.Y.; De Vos, M. SeqSleepNet: End-to-End Hierarchical Recurrent Neural Network for Sequence-to-Sequence Automatic Sleep Staging. IEEE Trans. Neural Syst. Rehabil. Eng. 2019, 27, 400–410.
- Pan, J.; Zhang, J.; Wang, F.; Liu, W.; Huang, H.; Tang, W.; Liao, H.; Li, M.; Wu, J.; Li, X.; et al. Automatic Sleep Staging Based on EEG-EOG Signals for Depression Detection. Intell. Autom. Soft Comput. 2021, 28, 53–71.
- Chen, T.; Huang, H.; Pan, J.; Li, Y. An EEG-based brain-computer interface for automatic sleep stage classification. In Proceedings of the 2018 13th IEEE Conference on Industrial Electronics and Applications (ICIEA), Wuhan, China, 31 May–2 June 2018; IEEE: New York, NY, USA; pp. 1988–1991.
- Pan, J.; Wu, J.; Liu, J.; Wu, J.; Wang, F. A Systematic Review of Sleep in Patients with Disorders of Consciousness: From Diagnosis to Prognosis. Brain Sci. 2021, 11, 1072.
- Roth, T. Characteristics and determinants of normal sleep. J. Clin. Psychiatry 2004, 65, 8–11.
- Izac, M.S.M. Basic Anatomy and Physiology of Sleep. Am. J. Electroneurodiagn. Technol. 2006, 46, 18–38.
- Reichert, C.F.; Maire, M.; Schmidt, C.; Cajochen, C. Sleep-Wake Regulation and Its Impact on Working Memory Performance: The Role of Adenosine. Biology 2016, 5, 11.
- Gerashchenko, D.; Murillo-Rodriguez, E.; Lin, L.; Xu, M.; Hallett, L.; Nishino, S.; Mignot, E.; Shiromani, P.J. Relationship between CSF hypocretin levels and hypocretin neuronal loss. Exp. Neurol. 2003, 184, 1010–1016.
- Siegel, J.M. Hypocretin (orexin): Role in normal behavior and neuropathology. Annu. Rev. Psychol. 2004, 55, 125–148.
- Brown, R.E.; Stevens, D.R.; Haas, H.L. The physiology of brain histamine. Prog. Neurobiol. 2001, 63, 637–672.
- Watanabe, T.; Taguchi, Y.; Shiosaka, S.; Tanaka, J.; Kubota, H.; Terano, Y.; Tohyama, M.; Wada, H. Distribution of the his-taminergic neuron system in the central nervous system of rats: A fluorescent immunohistochemical analysis with histidine decarboxylase as a marker. Brain Res. 1984, 1, 13–25.
- Ko, E.M.; Estabrooke, I.V.; McCarthy, M.; Scammell, T.E. Wake-related activity of tuberomammillary neurons in rats. Brain Res. 2003, 992, 220–226.
- Takahashi, K.; Lin, J.-S.; Sakai, K. Neuronal Activity of Histaminergic Tuberomammillary Neurons during Wake-Sleep States in the Mouse. J. Neurosci. 2006, 26, 10292–10298.
- McCarley, R.W. Sleep Neurophysiology: Basic Mechanisms Underlying Control of Wakefulness and Sleep. In Sleep Disorders Medicine. Basic Science, Technical Considerations, and Clinical Aspects; Chokroverty, B.H., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 51–59.
- Jones, B.E. From waking to sleeping: Neuronal and chemical substrates. Trends Pharmacol. Sci. 2005, 26, 578–586.
- Lena, I.; Parrot, S.; Deschaux, O.; Muffat-Joly, S.; Sauvinet, V.; Renaud, B.; Suaud-Chagny, M.F.; Gottesmann, C. Variations in extracellular levels of dopamine, noradrenaline, glutamate, and aspartate across the sleep—wake cycle in the medial prefrontal cortex and nucleus accumbens of freely moving rats. J. Neurosci. Res. 2005, 81, 891–899.
- Aston-Jones, G.; Bloom, F.E. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J. Neurosci. 1981, 1, 876–886.
- Gervasoni, D.; Darracq, L.; Fort, P.; Soulière, F.; Chouvet, G.; Luppi, P.-H. Electrophysiological evidence that noradrenergic neurons of the rat locus coeruleus are tonically inhibited by GABA during sleep. Eur. J. Neurosci. 1998, 10, 964–970.
- McCarley, R.W.; Hobson, J.A. Neuronal excitability modulation over the sleep cycle: A structural and mathematical model. Science 1975, 189, 58–60.
- Smith, Y.; Kieval, J.Z. Anatomy of the dopamine system in the basal ganglia. Trends Neurosci. 2000, 23, S28–S33.
- Jacobs, B.L.; Fornal, C.A. Activity of brain serotonergic neurons in the behaving animal. Pharmacol. Rev. 1991, 43, 563–578.
- Wu, M.-F.; John, J.; Boehmer, L.N.; Yau, D.; Nguyen, G.B.; Siegel, J.M. Activity of dorsal raphe cells across the sleep-waking cycle and during cataplexy in narcoleptic dogs. J. Physiol. 2004, 554, 202–215.
- Jones, B.E. Activity, modulation and role of basal forebrain cholinergic neurons innervating the cerebral cortex. Brain Res. 2004, 145, 157–169.
- Mileykovskiy, B.Y.; Kiyashchenko, L.I.; Siegel, J.M. Behavioral Correlates of Activity in Identified Hypocretin/Orexin Neurons. Neuron 2005, 46, 787–798.
- Kiyashchenko, L.I.; Mileykovskiy, B.Y.; Maidment, N.; Lam, H.A.; Wu, M.-F.; John, J.; Peever, J.; Siegel, J.M. Release of Hypocretin (Orexin) during Waking and Sleep States. J. Neurosci. 2002, 22, 5282–5286.
- Murillo-Rodríguez, E.; Arias-Carrión, O.; Sanguino-Rodríguez, K.; González-Arias, M.; Har, R. Mechanisms of Sleep-Wake Cycle Modulation. CNS Neurol. Disord. Drug Targets 2009, 8, 245–253.
- Dijk, D.-J.; Czeisler, C.A. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci. Lett. 1994, 166, 63–68.
- Dijk, D.J.; Cajochen, C. Melatonin and the circadian regulation of sleep initiation, consolidation, structure, and the sleep EEG. J. Biol. Rhythm. 1997, 6, 627–635.
- Easton, A.; Meerlo, P.; Bergmann, B.; Turek, F.W. The Suprachiasmatic Nucleus Regulates Sleep Timing and Amount in Mice. Sleep 2004, 27, 1307–1318.
- Sherin, J.E.; Shiromani, P.J.; McCarley, R.W.; Saper, C.B. Activation of Ventrolateral Preoptic Neurons during Sleep. Science 1996, 271, 216–219.
- Szymusiak, R.; Alam, N.; Steininger, T.L.; McGinty, D. Sleep–waking discharge patterns of ventrolateral preoptic/anterior hypothalamic neurons in rats. Brain Res. 1998, 803, 178–188.
- Lu, J.; Greco, M.A.; Shiromani, P.; Saper, C.B. Effect of Lesions of the Ventrolateral Preoptic Nucleus on NREM and REM Sleep. J. Neurosci. 2000, 20, 3830–3842.
- Lu, J.; Bjorkum, A.A.; Xu, M.; Gaus, S.E.; Shiromani, P.J.; Saper, C.B. Selective Activation of the Extended Ventrolateral Preoptic Nucleus during Rapid Eye Movement Sleep. J. Neurosci. 2002, 22, 4568–4576.
- Sterman, M.B.; Clemente, C.D. Forebrain inhibitory mechanisms:cortical synchronization induced by basal forebrain stimu-lation. Exp. Neurol. 1962, 6, 91–102.
- Sterman, M.B.; Clemente, C.D. Forebrain inhibitory mechanisms: Sleep patterns induced by basal forebrain stimulation in the behaving cat. Exp. Neurol. 1962, 6, 103–117.
- Siegel, J.M. Brainstem Mechanisms Generating REM Sleep. In Principles and Practice of Sleep Medicine; Kryger, M.H., Roth, T., Dement, W.C., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2000; pp. 112–133.
- Hallanger, A.E.; Levey, A.I.; Lee, H.J.; Rye, D.B.; Wainer, B.H. The origins of cholinergic and other subcortical afferents to the thalamus in the rat. J. Comp. Neurol. 1987, 262, 105–124.
- Kayama, Y.; Ohta, M.; Jodo, E. Firing of ‘possibly’ cholinergic neurons in the rat laterodorsal tegmental nucleus during sleep and wakefulness. Brain Res. 1992, 569, 210–220.
- Wislowska, M.; del Giudice, R.; Lechinger, J.; Wielek, T.; Heib, D.P.J.; Pitiot, A.; Pichler, G.; Michitsch, G.; Donis, J.; Schabus, M. Night and day variations of sleep in patients with disorders of consciousness. Sci. Rep. 2017, 7, 266.
- Bekinschtein, T.A.; Golombek, D.A.; Simonetta, S.H.; Coleman, M.R.; Manes, F.F. Circadian rhythms in the vegetative state. Brain Inj. 2009, 23, 915–919.
- Blume, C.; Lechinger, J.; Santhi, N.; del Giudice, R.; Gnjezda, M.-T.; Pichler, G.; Scarpatetti, M.; Donis, J.; Michitsch, G.; Schabus, M. Significance of circadian rhythms in severely brain-injured patients: A clue to consciousness? Neurology 2017, 88, 1933–1941.
- Cruse, D.; Thibaut, A.; Demertzi, A.; Nantes, J.C.; Bruno, M.-A.; Gosseries, O.; Vanhaudenhuyse, A.; Bekinschein, T.A.; Owen, A.M.; Laureys, S. Actigraphy assessments of circadian sleep-wake cycles in the vegetative and minimally conscious states. BMC Med. 2013, 11, 18.
- Isono, M.; Wakabayashi, Y.; Fujiki, M.M.; Kamida, T.; Kobayashi, H. Sleep cycle in patients in a state of permanent unconsciousness. Brain Inj. 2002, 16, 705–712.
- Mertel, I.; Pavlov, Y.G.; Barner, C.; Müller, F.; Diekelmann, S.; Kotchoubey, B. Sleep in disorders of consciousness: Behavioral and polysomnographic recording. BMC Med. 2020, 18, 350.
- Mura, E.; Pistoia, F.; Sara, M.; Sacco, S.; Carolei, A.; Govoni, S. Pharmacological modulation of the state of awareness in patients with disorders of consciousness: An overview. Curr. Pharm. Des. 2014, 20, 4121–4139.
- Yeo, B.T.T.; Tandi, J.; Chee, M.W.L. Functional connectivity during rested wakefulness predicts vulnerability to sleep deprivation. Neuroimage 2015, 111, 147–158.
- Shao, Y.; Wang, L.; Ye, E.; Jin, X.; Ni, W.; Yang, Y.; Wen, B.; Hu, D.; Yang, Z. Decreased thalamocortical functional connectivity after 36 hours of total sleep deprivation: Evidence from resting state FMRI. PLoS ONE 2013, 8, e78830.
- Miraglia, F.; Tomino, C.; Vecchio, F.; Gorgoni, M.; De Gennaro, L.; Rossini, P.M. The brain network organization during sleep onset after deprivation. Clin. Neurophysiol. 2021, 132, 36–44.
- De Weer, A.-S.; Da Ros, M.; Berré, J.; Mélot, C.; Goldman, S.; Peigneux, P. Environmental influences on activity patterns in altered states of consciousness: Activity patterns in disorders of consciousness. Eur. J. Neurol. 2011, 18, 1432–1434.
- Schabus, M.; Wislowska, M.; Angerer, M.; Blume, C. Sleep and circadian rhythms in severely brain-injured patients—A comment. Clin. Neurophysiol. 2018, 129, 1780–1784.
- Blume, C.; Angerer, M.; Raml, M.; del Giudice, R.; Santhi, N.; Pichler, G.; Kunz, A.B.; Scarpatetti, E.; Trinka, E.; Schabus, M. Healthier rhythm, healthier brain? Integrity of circadian melatonin and temperature rhythms relates to the clinical state of brain-injured patients. Eur. J. Neurol. 2019, 26, 1051–1059.
- Arnaldi, D.; Terzaghi, M.; Cremascoli, R.; De Carli, F.; Maggioni, G.; Pistarini, C.; Nobili, F.; Moglia, A.; Manni, R. The prognostic value of sleep patterns in disorders of consciousness in the sub-acute phase. Clin. Neurophysiol. 2015, 127, 1445–1451.
- Cologan, V.; Drouot, X.; Parapatics, S.; Delorme, A.; Gruber, G.; Moonen, G.; Laureys, S. Sleep in the unresponsive wake-fulness syndrome and minimally conscious state. J. Neurotrauma 2013, 30, 339–346.
- Forgacs, P.B.; Conte, M.M.; Fridman, E.A.; Voss, H.U.; Victor, J.D.; Schiff, N.D. Preservation of electroencephalographic organization in patients with impaired consciousness and imaging-based evidence of command-following. Ann. Neurol. 2014, 76, 869–879.
- Kotchoubey, B.; Pavlov, Y. Sleep patterns open the window into disorders of consciousness. Clin. Neurophysiol. 2018, 129, 668–669.
- Sebastiano, D.R.; Visani, E.; Panzica, F.; Sattin, D.; Bersano, A.; Nigri, A.; Ferraro, S.; Parati, E.; Leonardi, M.; Franceschetti, S. Sleep patterns associated with the severity of impairment in a large cohort of patients with chronic disorders of consciousness. Clin. Neurophysiol. 2018, 129, 687–693.
- Brown, R.; Basheer, R.; McKenna, J.; Strecker, R.E.; McCarley, R. Control of Sleep and Wakefulness. Physiol. Rev. 2012, 92, 1087–1187.
- Landgraf, D.; Shostak, A.; Oster, H. Clock genes and sleep. Pflügers Arch. Eur. J. Physiol. 2012, 463, 3–14.
- Sakurai, T. The neural circuit of orexin (hypocretin): Maintaining sleep and wakefulness. Nat. Rev. Neurosci. 2007, 8, 171–181.
- Claustrat, B.; Brun, J.; Chazot, G. The basic physiology and pathophysiology of melatonin. Sleep Med. Rev. 2005, 9, 11–24.
- Reiter, R.J. Pineal Melatonin: Cell Biology of Its Synthesis and of Its Physiological Interactions. Endocr. Rev. 1991, 12, 151–180.
- Nambu, T.; Sakurai, T.; Mizukami, K.; Hosoya, Y.; Yanagisawa, M.; Goto, K. Distribution of orexin neurons in the adult rat brain. Brain Res. 1999, 827, 243–260.
- Yang, X.-A.; Song, C.-G.; Yuan, F.; Zhao, J.-J.; Jiang, Y.-L.; Yang, F.; Kang, X.-G.; Jiang, W. Prognostic roles of sleep electroencephalography pattern and circadian rhythm biomarkers in the recovery of consciousness in patients with coma: A prospective cohort study. Sleep Med. 2020, 69, 204–212.
- Lucca, L.F.; Lofaro, D.; Pignolo, L.; Leto, E.; Ursino, M.; Cortese, M.D.; Conforti, D.; Tonin, P.; Cerasa, A. Outcome prediction in disorders of consciousness: The role of coma recovery scale revised. BMC Neurol. 2019, 19, 68.
- Gottshall, J.L.; Rossi Sebastiano, D. Sleep in disorders of consciousness: Diagnostic, prognostic, and therapeutic considerations. Curr. Opin. Neurol. 2020, 33, 684–690.
- Nekrasova, J.; Kanarskii, M.; Yankevich, D.; Shpichko, A.; Borisov, I.; Pradhan, P.; Miroshnichenko, M. Retrospective analysis of sleep patterns in patients with chronic disorders of consciousness. Sleep Med. X 2020, 2, 100024.
- Silverman, D. Retrospective study of EEG in coma. Electroencephalogr. Clin. Neurophysiol. 1963, 15, 486–503.
- Kang, X.-G.; Yang, F.; Li, W.; Ma, C.; Li, L.; Jiang, W. Predictive value of EEG-awakening for behavioral awakening from coma. Ann. Intensive Care 2015, 5, 52.
- Hulihan, J.F., Jr.; Syna, D.R. Electroencephalographic sleep patterns in post-anoxic stupor and coma. Neurology 1994, 44, 758–760.
- Oksenberg, A.; Gordon, C.; Arons, E.; Sazbon, L. Phasic Activities of Rapid Eye Movement Sleep in Vegetative State Patients. Sleep 2001, 24, 703–706.
- Scarpino, M.; Lolli, F.; Hakiki, B.; Lanzo, G.; Sterpu, R.; Atzori, T.; Portaccio, E.; Draghi, F.; Amantini, A.; Grippo, A. EEG and Coma Recovery Scale-Revised prediction of neurological outcome in Disorder of Consciousness patients. Acta Neurol. Scand. 2020, 142, 221–228.
- Wielek, T.; Lechinger, J.; Wislowska, M.; Blume, C.; Ott, P.; Wegenkittl, S.; del Giudice, R.; Heib, D.P.J.; Mayer, H.A.; Laureys, S.; et al. Sleep in patients with disorders of consciousness characterized by means of machine learning. PLoS ONE 2018, 13, e0190458.
More