1. Overview to T Cells and the Adaptive Immune System
T cells are thymus-derived lymphocytes that provide adaptive immune defense representing 56.6–84.6% of the B/T and NK cell populations [
327,
328]. The discovery of T cell CD4 (Leu-3 and T4) receptors occurred in the late 1970s, whereas a cluster of differentiation
(CD) markers were clarified in 1984 at the first workshop on HLA molecules [
329]. Furthe
r cellular phenotypes
were defined thereafter includ
eding T
H1, T
H2, T
H9, T
H17, T
H22, T
C, and T
REGS. T cell development occurs in bone marrow lacking characteristic CD4
+ and CD8
+ receptors (DN), undergoing SHM and selection processes that generate CD4
+CD8
+ double positive (DP) thymocytes or others that include CD4
+ or CD8
+ single positive (SP) thymocytes that emerge into the periphery as T cells. These generally exhibit a CD45RA
+CCR7
+ phenotype, although CCR7 is also expressed on DCs. The comparatively recent discovery of T
REG cells by Sakaguchi in 1995 defined with clarification that CD4
+CD25
+ cells express a nuclear transcription factor forkhead box P3 (FoxP3) that regulates development [
322]. These also undergo clonal expansion where activation/differentiation occurs into effector T cells that mediate pathogen clearance, after which T cells undergo apoptosis, while memory T cells can persist with variable responses, with further data below to compare these with historical comparable standards of previous viral pandemics such as influenza, RSV, or smallpox. Cytotoxic CD8
+ T cells are known to use a variety of proteolytic enzymes, including granzymes, while CD4
+ T cells regulate maintenance through T
H1 and T
H2 lineages in concert with this CD8
+ T cell response via regulation of exhaustion and antigen recognition receptors. T cells are further defined as naïve CD3
+ CD4
+ CD45RA
+ CCR7
+ (T
N), which differentiate into central memory (T
CM: CD45RA
−CCR7
+), effector-memory (T
EM: CD45RA
−CCR7
−), stem-cell memory (T
SCM: CD45RA
+CCR7
+CD95
+CD122
+), and peripheral resident memory (T
RM CD69, CD103, CD49) T cells [
330]. T
RM cells produce key cytokines IFN-γ, IL-17, TNF-α, and IL-2, and these cells can express PD-1, LAG-3, and CTLA-4. In serum, CCR7 distinguishes T cell homing to LN when expressed, or effector memory (T
EM) subsets migrating to tissues when absent. However, as previously stated
, in our earlier article cells can receptor characteristic immune cell recognition receptors that are named double negative DN(DN) B cells
andor DN T cells which do not express CD4 or CD8
athat are known to exist [
331]. Below is illustrated all above current T cell classifications by
clustCD marker
of differentiation marker (see Figure 1)(see Figure 1). Within these innate and adaptive cell receptor complexes lies complexities of many pathological conditions and protein recognition.
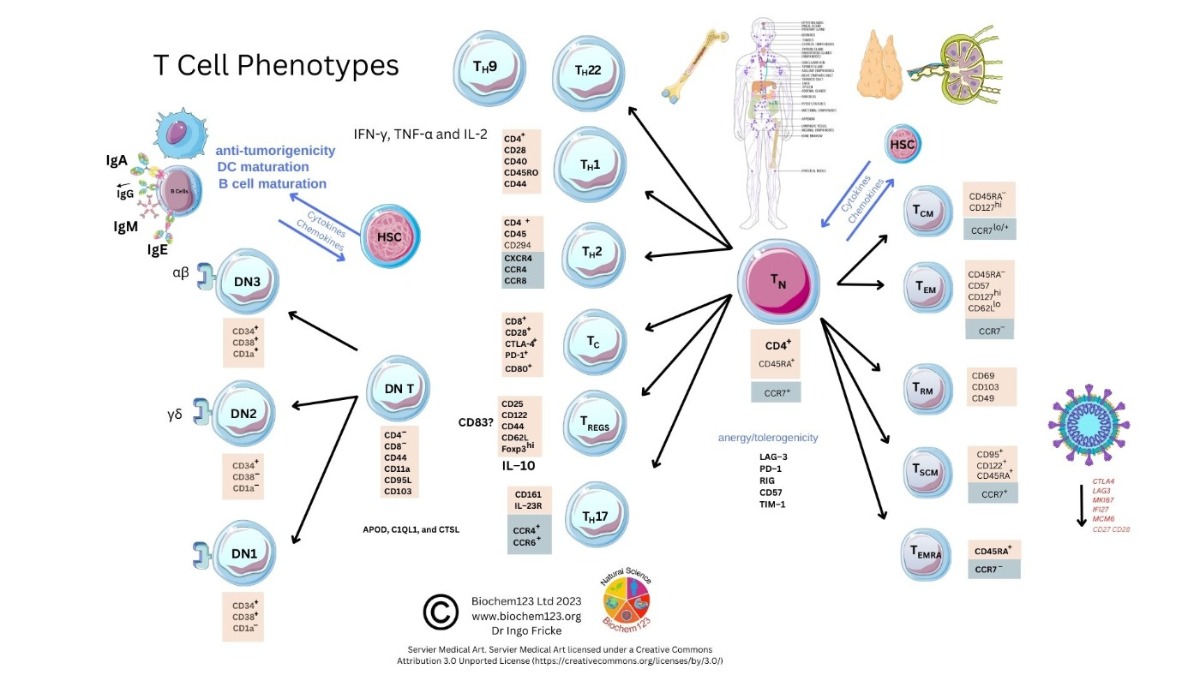 Figure 1. T cell phenotype diversity and developmental cellular markers.
Figure 1. T cell phenotype diversity and developmental cellular markers.The TCR transcends T cell membranes, recognizing fragments of antigenic peptides bound to major histocompatibility complex (MHC class I/II) molecules in conjunction with other co-receptors.
MHC class I/II molecules present antigenic peptides processed intracellularly from varying cells in conjunction with other receptors or costimulatory molecules. Similarly, TCRs
unas BCRs and other receptors including MHC complexes undergo recombination during development to create
a heterodimeric receptor
s. Most (95%) are composed of alpha and beta chains (α/β), but also gamma and delta chains (γδ) (5%), that transduce signals through receptor/peptide/MHC complexes located near the CD3 receptor which, again, has CDR regions. Therefore, TCRs bind with different affinities and utilize other T cell co-receptors (CD4/CD8) to affect cellular requirements via other cells affected by cytokines expressed with MHC class I or II antigenic peptide presentation to these T cells, and all are affected by cytokines [
332].
2. Background to T Cells in Coronaviruses and Host SARS-CoV-2 Infection
Prior COVID-19 research on immunogens understandably focused on nAb responses, with less interest in overall cellular immunity. Interestingly, there is data accumulating which suggests that T cell responses are an important player i
n n
vaatural infection and adaptive immune responses as well as vaccine protection against chronic COVID-19 disease, even more so against recent variants of Omicron lineages (BA1, BA2, BA4/5, BQ1, and XBB
1.5) that display further epitope escape from recognition by nAbs. Epitopes represent the unique protein sequence that either viral antigens or vaccine immunogens stimulate recognition of, evoking protection against viral pathogens. These observations should have an impact for using current COVID-19 vaccines and for the development of next-generation vaccines against COVID-19 and other infectious diseases [
333]. Early in the pandemic in 2020, case reports appeared across the globe (e.g., Italy) describing people with a rare disorder showing deficiency in antibody production or even no B-cells at all (e.g., agammaglobulinemia-XLA) [
334]. Routine surveillance in this risk group identified two XLA patients who developed COVID-19 while receiving Ig infusions. These patients developed pneumonia which eventually resolved but never required ventilation or intensive care. This was the first time that B-cell response might be considered important, but it is not strictly required to overcome COVID-19 disease [
335,
336]. In addition, in 2020, groups in Germany and the USA independently discovered reactive T cells to the S glycoprotein during SARS-CoV-2 infection in around 30% of healthy donors when analyzing the T cell response of COVID-19 patients [
337,
338]. These T cells belonged to the CD4
+ lineage and were primarily directed against C-terminal-epitopes of S protein, which has higher homology among S glycoproteins of hCoVs than the N-terminal region. Braun et al. further showed that these cross-reactive T cells were functional against the S protein C-terminal domain of the human endemic coronaviruses hCoV-229E and hCoV-OC43, as well as that of SARS-CoV-2 [
338]. Daniela Weiskopf et al. mapped 142 T cell epitopes across the SARS-CoV-2 genome by using pre-pandemic blood samples to simplify the exact investigation of the SARS-CoV-2-specific CD4
+ T cell repertoire. They demonstrated an array of pre-existing memory CD4
+ T cells with cross-reactive capability with equal affinity to SARS-CoV-2 and the common cold coronaviruses hCoV-OC43, hCoV-229E, hCoV-NL63, and hCoV-HKU1 [
337].
As research continued, all B and T cell markers CD3/CD4/CD8/CD19 were re-profiled as described above and below. Therefore, early indications of lymphocyte dysfunction elucidated this to show that T suppressor cell populations possessing the T cell activation marker CD28 required clarity. Furthermore, CD28 requires co-stimulatory molecules CD80 and CD86, of which CD80 is present on B cells, DCs, T cells, and Mϕ (requiring co-stimulatory molecules CD80/CD86). Hence, within chronic COVID-19 disease, and as there are changes within the T cell subsets, it is of note that, within T cell subsets, DN T cells exist that do not express classical CD4 or CD8 receptors. SARS-CoV-2 reactive T cells were found in peripheral blood and tonsil from donors unexposed to SARS-CoV-2, indicating that it is possible that antigens are present in other pathogens [
339,
340,
341,
342,
343,
344,
345,
346,
347]. A number of these samples were analyzed by reports prior to the COVID-19 pandemic, eliminating the likelihood of exposure of the donors to SARS-CoV-2 infection [
339,
340,
342,
343,
346,
347]. An explanation to this observed phenomenon is that researchers are observing a recall response of SARS-CoV-2-cross-reactive memory T (T
MEM) cells that were created upon encountering homologous proteins derived from other coronaviruses. Before SARS-CoV-2, six hCoVs were circulating with SARS-CoV, with MERS-CoV sharing the highest homology to SARS-CoV-2 in that it also caused respiratory syndromes [
348]. Le Bert et al. even reported in blood samples from convalescent SARS-CoV-1 (>17 years ago) that T cells were detected which recognized SARS-CoV-2 [
346]. Nevertheless, for SARS-CoV-1 and MERS-CoV, the dissemination was more restricted than for SARS-CoV-2, meaning that it is unlikely to find cross-reactivities over a wide range of the population. SARS-CoV-2 reactive T cells were found in peripheral blood and tonsils from donors unexposed to SARS-CoV-2 [
339,
340,
341,
342,
343,
344,
345,
346,
347]. Though it is still heavily debated and unclear whether common cold hCoVs mediated this, cross-reactive T cells are of benefit in host defense against COVID-19 disease. Interestingly, in 20% of SARS-CoV-2-naïve donors and a higher percentage of infected or vaccinated donors, a report determined that a SARS-CoV-2 conserved peptide, S816-830, found in common cold hCoVs, activates CD4
+ T cells, which led the authors to the conclusion that cross-reactive CD4
+ T cell response might be protective against COVID-19 disease [
349]. In contrast to these findings, a different group found the same epitope activated CD4
+ T cells in more donors with breakthrough infections after vaccination compared to vaccinated donors with no breakthrough infection [
350]. Moreover, it was reported by Bacher and colleagues that CD4
+ T cells cross-reactive to SARS-CoV-2 in individuals not exposed to the virus displayed low avidity, which similarly correlates with individuals with severe COVID-19 disease [
345]. These cross-reactive CD4
+ T cells could be determined
in vitro as responsive to SARS-CoV-2 proteins, leading to proliferation of the concomitant T cell clones stimulated by SARS-CoV-2. The relevance of these findings requires further investigation. On the other hand, four human coronaviruses (hCoV-OC43, hCoV-HKU1, hCoV-NL63, and hCoV-229E) circulate each year, causing the common cold. These four hCoVs share less homology with SARS-CoV-2. Indeed, it remains evident that cross-protective T cell epitopes seen in common cold hCoVs recognize SARS-CoV-2 [
342,
349,
350,
351]. Commensal bacteria as well as hCoVs can induce cross-reactive T cells to SARS-CoV-2 infection [
352,
353]. In COVID-19 patients, as well as in healthy controls, a public T
FH clonotype was detected that recognizes a SARS-CoV-2 epitope, S870-878 protein, and shares homology with a symbiotic bacterial antigen. Its abundance was found to be higher in patients with acute compared to chronic symptoms as compared to individuals with severe symptoms, leading to predictions that this clonotype has protective attributes [
352]. Unexpectedly, the old vaccine strain BCG displayed homology with eight epitopes of NSP3 from SARS-CoV-2. In this case, Eggenhuizen et al. demonstrated that
in vitro stimulation with SARS-CoV-2 proteins of BCG-primed CD4
+ and CD8
+ T cells led to enhanced cytokine production and proliferation in an HLA-dependent fashion. This mechanism might provide a partial explanation for the observation that BCG vaccination exerts some protection from COVID-19 [
354]. Nonetheless, further studies to elucidate the exact features of cross-reactive T cells that could differentiate between SARS-CoV-2 infection and other pathologies will be required.
3. Helper T Cell Role during the Adaptive Immune Response to SARS-CoV-2
Historically, T cells were classified as T
H1/T
H2 and/or T
C types (CD4
+ and CD8
+), with
the HIV discovery elucidating CD4
+ receptor mediated entry some 30 years ago; however, between 2000 and 2014, further classification of chemokines as predominant homing ligands and receptors occurred, which was of key relevance to immune system regulation. T
H1 cells secrete IFN-γ, IL-2, and TNF in comparison to T
H2, where IL-4, IL-5, IL-6, and IL-10 are produced. CTLA4 (CD152) is preferentially expressed by T cells and regulates immune responses through interactions with B7-1 (CD80) and B7-2 (CD86) on antigen presenting cells (APC). A similar molecule, PD-1 (CD279), is also involved in immunoregulation through ligands PD-L1 and PD-L2. More recently, TIGIT has been found to regulate immune responses through interaction with CD112 and CD155 in a manner analogous to CTLA4. In contextual terms, the CD28 super-family of regulatory leukocyte receptors includes ICOS, CD28, CTLA4, PD-1, and BTLA, of which CD28 is a co-stimulatory T cell activation molecule for B cell ligands CD80 and CD86, compared to PD-1 as an inhibitory receptor for PD-L1/PD-L2. Recently, it has been observed that immunocompromised cohorts with immune-related pneumonitis possess more CD4 T
H2 cells expressing CCR7, but also that CXCR3
+GATA3 is preferentially expressed [
355]. Initially, it was proposed that immunity could be divided into three types, in short, initially, IFN-γ producing cells (NK cells, T
C and T
H1) regulating activation of monocytes; secondly, T
C and T
H2 regulation of mast cells, basophils, and eosinophils through IgE; and a third type mediated by RORγt
+ T cells and T
H17 regulation of monocytes and neutrophils [
70,
356]. Early in 2020, a noted marked reduction in T cells was seen in laboratory studies (n = 552) in SARS-CoV-2 infection with CD4
+CD8
+ cell counts (75.95%:71.54% respectively) during COVID-19 disease, accompanied by upregulation of PD-1 with suggestions that IL-6, IL-10, and TNF-α were not produced by T cells [
357]. This was a key finding, suggesting that cell dysfunction, as PD-1 (CD239), is an inhibitory receptor that regulates both chronic viral infection and cancer through affecting T cell apoptosis pathways. This dysfunction occurs via binding of PD-1 to PD-L1 and PD-L2 on APCs and effectively controlling IFN-γ cellular release. In addition, lymphocyte activation gene 3 (LAG-3), together with T cell Ig and mucin domain 3 (TIM-3) pathways, were upregulated with concurrent sCD28, sCTLA-4, sBTLA, CD270, and sCD80 increases [
358,
359]. Comparatively less is known about LAG-3, although it is not present on naïve T cells and is constitutively expressed on T
REGS implicated with T cell exhaustion. LAG-3 presents on DN T cells and intraepithelial lymphocytes, γδ T cells, and NK cells, and therefore remains a focus of cancer research as a checkpoint protein [
360,
361]. CD28 concurrent reduction and shedding could have significant effects on B lymphocyte signaling through respective ICOS ligands, and all of these would usually be expressed within the GC of primary lymphoid organs during follicular development. As discussed above, DC subtypes form the core of immune response cells through expression of MHC class II and changes to subtypes within SARS-CoV-2 infection, suggesting that systemic cDC2 cells undergo changes that could affect the maturation and priming of T
FH antibody responses [
362]. Furthermore, during infection, individuals with COVID-19 disease displayed a higher expression of T cell Fas (CD95) and sCD95 in CD4
+ T cells, but also PS correlated with T cell counts indicating an overall increase in T cell apoptosis and increases in caspase 1/3, and this intracellular activation within both CD4/CD8 T cells was confirmed [
363]. Notably, other studies indicate that PS receptors can be associated with increases in infection rates, with increased CD95 implicated in the regulation of auto-reactive T cells [
364].
The Gil-Mansos group recently conducted a comprehensive FACS analysis comparing a control vs COVID-19 disease group (n = 51) within recovered COVID-19 individuals to demonstrate a significant cell cluster at 10 months after recovery of CD4
+ CD45RA
− CCR4
− CCR10
− CD27
+ CCR6
−CXCR3
+ CD127
+ cells. Their analysis inferred a response within circulating T
FH1, PBs, and the follicular-dendritic-cells (foDC) axis [
17]. In this report, it was found that T
FH cells within the SARS-CoV-2 infected group expressed both ICOS and PD-1, which is relevant to normal responses occurring outside the GC that could affect T
FH/B cell lymphocyte development [
10]. Therefore, a potential imbalance within T
CM cells during acute COVID-19 was seen to occur, which polarizes T
FH lineages leading to potential dysregulation of the B cell response outside GC. This was quantified to find the CD4
+ T cell’s tendency to T
H2-like phenotypes in chronic SARS-CoV-2 infection that increases in CD45RA
+CD62L
− (T
CM) and CD45RA
−CD62L
− (T
EM), compared to the decreases in T
CM CCR6
+ T
H17-like cells [
365]. Through stimulation of these T
FH cells by agonism of the ICOS receptor that would produce normal synthesis of IFN-γ/IL-21, it was observed in samples from hospitalized COVID-19 subjects that T
FH cells (CD3
+ CD4
+ CXCR5
+ ICOS
+) within ambulatory subjects encompassed, on average, 15% T
FH of the peripheral blood CD4
+ compartment relative to the normal 8% T
FH population [
366]. Vanderbilt University’s recent single cell sequencing study expanded on the importance of this cell subset in a healthy cohort (n = 10) where analyses further showed functional T
H cells as CD4
+CD38
++ICOS
++Ki67
+ (but also expressing CTLA4, CD44, CD45RO, CD127, and others) alongside key chemokine homing receptors CXCR3, CXCR5, and HLA-DR, confirming that cell signaling receptors were expressed and that the T cell proliferation marker Ki67 was working normally in T
N subsets [
367]. Epitope determinants of SARS-CoV-2-specific CD4
+ T cell lines reveal SARS-CoV-2 M protein-driven dysregulation of ISG in chronic COVID disease (ISG15, IFITM1, IFI16, MX1, STAT1, OAS1, IFI35, IFIT3 and IRF7) [
368]. The importance of a T
H1 response cannot be underestimated in immunogen development and is historically thought to be the identifying factor producing IFN-γ, IL-2, and TNF to evoke cell-mediated immunity and phagocyte-dependent inflammation. Current
in vitro and in clinical settings research is indicative that, within the CD4
+ T
FH compartment, CXCR3
+ expression is upregulated in acute cases and then downregulated with severity in COVID-19 disease in PB cells, and also that CD4
+ T cells expressing PD-1
hi do not express CXCR5 [
162,
369,
370,
371,
372]. The dynamics of each cell type can only be identified by individual markers. Therefore, given the above, it is apparent that, in case control studies (n = 27) in other inflammatory conditions, such as RA, where autoantibodies affect homeostasis and TNF-α, production seems to localize with specific particular epitopes on T cells near the citrullinated vimentin co-receptor that also potentially SARS-CoV-2 utilizes [
12,
285,
374]. This has been observed to be blocked by utilizing anti-TNF therapeutics under research investigation. Interesting, this study measured migratory T
H CD4
+CD45RA
+ cells expressing CD62L
+CD95
+ that can be downregulated or shed, respectively, in dysregulated similar pathologies [
285,
371,
372].
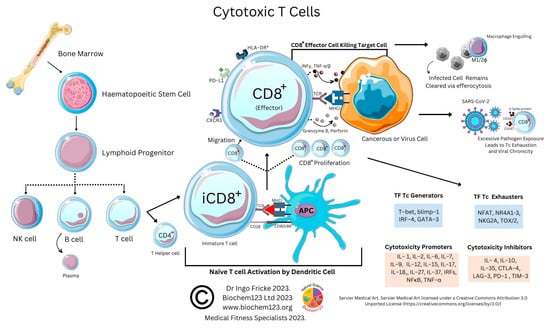
4. Cytotoxic T Cell Role during the Adaptive Immune Response to SARS-CoV-2
Cytotoxic CD8
+ T cells (T
C) are major killers of pathogens and neoplastic cells, with CD4
+ T cells playing important roles in the maintenance of the CD8
+ response and the prevention of exhaustion that utilizes a variety of mechanisms. Like above, CD8
+ T cells differentiate into effector cells and produce IFN type I with Mϕ and DCs. Other cytokines, such as TNF-α, IL-15, and IL-18, facilitate CD8
+ T cell response. T
C cells, for example, are generated in the presence of IL-2 and IL-12 and acquire the ability to secrete cytokines such as IFN-γ. Cytotoxicity of T
C cells is determined by granular enzyme release, such as granzyme B and perforin, with virulence toxicity varying between cytotoxic T cells depending on cytokine stimulation, but also transcription factors implicated in T
C generation, including T-bet, blimp-1, IRF-4, and GATA-3. Illustrated below (see
Figure 1) are interactions of T
C cells and relevant cytokines. It can now be seen that further information is available on these cells, which are decreased in immunocompromised cohorts with immune-related arthritis that have lower levels of CD8 T
CM cells [
355].
Figure 2. Cytotoxic T cell interactions and cytokines
In SARS-CoV-2 infected patients, it was seen that T cells expressing CD28 could be downregulated, but CD8
+ T cells also express PD-L1 and CXCR3
+, potentially also significantly associated with survival in SARS-CoV-2 infection alongside N protein specific CD8
+ cells [
373]. As before, groups in Holland saw enrichment of CCR6
+ CCL20
+ within CD8 T cells during COVID disease (n = 22) in BALF samples compared to serum, with fewer T
H1 cells indicating preferential differentiation and migration of this subset in severity into the lung, but also an impaired T
H cell response [
374]. During SARS-CoV-2 infection, there was also a significant increase in terminally differentiated T
EM cells (T
EMRA) and T
EM cells localizing with T
CM (CCR7
+ CD45RA
−) differentiating into T
H2-like secreting IL-5, with T
CM cells expressing chemokines (CX3CR1, CCR6, CXCR6, and CXCR3 that represent lung-homing receptors). T
CM cells could secrete TNF-α, but also SARS-CoV-2 CD4 S-protein-specific T cells displaying a T
CM phenotype with specific CD8
+ cells heterogenous towards T
EM, T
EMRA, and T
SCM [
155,
370,
373]. Therefore, as lymphopenia can occur and cause potential insufficient antibody responses in SARS-CoV-2 infection, it is necessary to look at dysfunction inside and outside GCs in COVID-19 disease [
155,
370,
375]. Reports are conflicting around CD3
+CD8
+ cells expressing CD38 that downregulate MHC class II (HLA-DR) during SARS-CoV-2 infection, although others indicate that this correlates with disease severity [
376,
377]. In some cohort studies within immunocompromised patients, it does, however, appear that different immunogens evoke individually different responses that could vary by underlying comorbidity. For example, healthy controls would express B cells (CD38
+CD19
+) and T cells (CD8
+ HLA-DR
+) three weeks after two doses of vaccine (n = 42); in comparison to transplant patients, little change was seen in cells expressing HLA-DR
+, CD38
+, and PD1
+, but in immunocompromised cohorts, 13.3% produced an IgG S-protein-specific response [
378]. This single cell analysis showed changes, or rather increases, in CD8
+ T
EM/T
EMRA cells [
299]. More recently, in a yet to be reviewed pre-print, elevated or prolonged CD8
+ activation was seen to be accompanied by elevations in IL-4, IL-7, IL-17, and TNF-α [
379]. Furthermore, this was accompanied by CD8
+PD-1
+CXCR3
loCCR6
+ T cells in severity in the lung, which suggested a reduced cytotoxic antiviral function of cytotoxic T cells [
374]. Recent, suggestions have been made that, in severity compared to moderate illness, the CD4 CD8 dysregulation at 3 months is not related to the cytokines produced that are predominant but that affect each of the T cell compartments through IL-4, IL-7, IL-17, and TNF-α [
379]. Therefore, as immunogenic proteins are seen to affect the adaptive immune response, it was also quantified that although 86% of individuals possessed antibodies against S2 protein, more were produced against RBD, and that T
C epitopes at specific amino acid residues (884–891 and 1116–1123) were identified that could be responsible for cross-reactivity, with some indications that this was with another hCoV (OC-43) [
130].
5. Regulatory T Cell Overview during the Adaptive Immune Response
CD4
+ Foxp3
+ regulatory T cells (T
REGS) are a central component of immune regulation, as humans are reliant on T
REGS expressing Foxp3 gene, which has been extensively investigated in the context of cancer regulation and COVID-19. Follicular T
REGS (T
FR cells) can inhibit antibody production, whereas follicular T
H cells (T
FH cells) stimulate it. T
FR cells are found in blood; however, comparatively less is known about the developmental signals. T
REGS represent a fraction of total CD4
+ T cells in sera and tissue, which constitutively express CD25 that can be shed and is part of the IL-2 receptor. Nearly all laboratory case studies indicate that current therapeutic COVID-19 remedies do in fact act to increase this subset, with several molecular mechanisms proposed. T
REGS constitutively expresses high affinity IL-2 receptors and scavenges the T cell growth factor IL-2, thus preventing suppressor actions on proinflammatory T cells. T
REGS also express a surface receptor molecule CD39 (ENTPD1ATP), which hydrolyses ATP within and outside cells. In notable disorders such as autoimmune thyroid conditions (e.g., GD), where prevalence estimates are 2–5% of the population, the role of T
REGS is likely to be clearer, as it was seen that in untreated autoimmune thyroid diseases (AITD) there was a clear reduction in T
REGS (RORγt/Foxp3) [
380]. Autoimmune thyroid diseases (AITDs) also involve immunologic dysregulation, with the presence of cell and humoral immune responses against thyroid gland antigens, T and B cell infiltration, and autoantibody generation; they are among the most common autoimmune disorders. AITDs include Hashimoto’s or chronic lymphocytic thyroiditis, which is a hypothyroid condition, and GD, a hyperthyroid condition. AITDs emerge via epigenetic and gene-environment interactions, and affected individuals are at risk for additional autoimmune diseases through associated genes, including immunoregulators (e.g., HLA, CTLA-4) and thyroid specific components (e.g., TSH receptors, thyroglobulin, and others). Individuals with immune-related thyroiditis have more CD4 T
H17 cells at baseline [
355]. T
H1, T
H2, T
H17, T
H22, and T
H9 lymphocytes are involved in AITD development, as well as T
REGS cells. Large scale, real-world studies are beginning to establish links between other disorders such as asthma and the risk of AITD. Such studies should provide information to optimize clinical guidance regarding how to diagnose and treat people with autoimmune disorders as researchers further unravel the complex endocrinological and inflammatory interactions and mechanisms that are involved [
381,
382]. T
REGS function, as above, can additionally be seen in DM1 patients where impairment of selected chemokines CCL3 and CCL4 has been suggested to impair adaptive T cell suppressive responses [
383]. Case studies are contrasting, with respect to T
REGS cells to date, with some indicating that T
REGS increase with COVID-19 severity and others indicate a decrease in quantity [
384]. A notable recent study (n = 19) examined these CD3
+CD4
+CD25
+CD127
low T cells up to 3 months after SARS-CoV-2 infection to find that IL-10 and TGF-β were produced upon stimulation with either S1 or N protein, and, importantly, that CD8
+ cells were producing IFN-γ, as all 3 cytokines are required for suppression, proliferation, and anti-viral activity [
385]. However, recently, studies on a novel cell type MDSCs, although not conclusive, are emerging to elucidate that, far from being clear, implicate TGF-β in COVID-19 MDSCs as suppressing CD4
+ CD25
+ Foxp3
+ cells and evoking CD4
+ CD25
−Foxp3
+ expansion, affecting T
REGS, the autoreactivity of T cells, and suppressing IFN-γ [
187,
386]. The differences between T
FR and T
REGS have been examined prior, in order to observe whether CD25 signaling is independent or dependent on IL-2. Clarification of blimp-1 and BCL6 elucidated that CD25
− T
FR cells lose part of their IL-2-dependency in T
REGS in exchange for enhanced expression of BCL6 and other T
FH-related genes with CXCR5 expression [
387]. T
REGS have also, interestingly, played a role through chemokine research developments in preventing bowel inflammation in colitis patients by suppressing cytokines related to T
H17 [
388,
389,
390,
391,
392].
6. TH17 Cell Overview during the Adaptive Immune Response
The complete cellular relationship between T
H17 and T
REGS cells is comparatively clearer, as both appear to have a symbiotic relationship in maintaining regulation [
388]. It is believed that this occurs through T
H17 transcription factors STAT3 and RORγt, which regulate host cell proliferation or suppression. T
H17 cells are characterized by cytokine secretion (IL17A, IL17F, IL21, and IL22), together with receptors (CD161, CCR4, and CCR6) [
393]. They are also considered to be regulated by DCs through IL-6, IL-1β, TGF-β and IL-23, which, in turn, polarize towards T
H17 cells that differentiate into one of four or more phenotypes, T
H1, T
H2, T
FH, or T
REGS cells expressing the IL-23 receptor, thereby being responsive to DC IL-23 secretion [
394]. For example, in IBD, excessive IL-17 secretion may cause excessive cell proliferation and differentiation, and similarities with IL-1 expression in mucosa between individuals affected by either Crohns or UC, with expression of CCL3 CCL4, CCL5, CCL7, CXCL5 CXCL8, CXCL10, CCL20, CXCL5, CXCL8, and CXCL10 in epithelial compartments with changes in other chemokines that bear similarities to post-acute sequelae of COVID-19 (PASC) [
395,
396,
397,
398,
399,
400]. Due to the dysregulation, or rather, upregulation, of T
H17 and T
H1, with a combination of reduction in the number of T
H2 and T
REGS and imbalances in the immune system in SARS-CoV-2 induced COVID-19 disease, the levels of inflammatory cytokines and chemokines changes occurring, such as CCL3, GM-CSF, TNF-α, IFN-γ, G-CSF, IL-2, IL-8, and IL-1β, are increased in localized tissues, leading to cytokine release syndrome (CRS), inducing further damage to the respiratory system with concurrent changes to vascular permeability [
389]. Early in the COVID-19 pandemic, researchers observed a skewed and increased ratio of T
H17 cells in individuals, in severity (n = 80) of this order: T
H17/T
REGS, RORγt/Foxp3, and IL-17/IL-10. This illustrates not only increased T
H17 cells, but that suppression occurred of T
REGS and IL-10 with less Foxp3 expressing T
REG cells [
401]. More recently, it has been confirmed, in Egypt, in a pediatric cohort of mixed COVID-19/MIS-C individuals, (n = 31) that IL-17A was upregulated [
402]. Therefore, given that this is consistent, it appears as though SARS-CoV-2-T
REGS-induced COVID-19 disease can be associated with a decrease in T
REG cells and an increase in T
H17 cells. However, different T cell populations, as above, are present in deceased COVID-19 patients and in improved patients. Disease progression in severity seems to reflect this (n = 7) compared with improved patients (n = 23) that had higher T
H1, lower T
H2, higher T
H1/T
H2 ratio, lower IL-4, higher IFN-γ, higher IFN-γ/IL-4 ratio (d), lower T
REGS, higher T
H17, higher T
H17/T
REGS ratio, lower IL-10, higher IL-17, higher IL-17/IL-10 ratio, and higher depleted T cells. It has been suggested that the T
H17-IL-17A axis may be a plausible therapeutic target for COVID-19 that could be researched to investigate in clinical trials. In PBMCs from a COVID-19 individual, T
H17 cells were significantly increased, leading to a cytokine storm, which, in turn, accelerated disease progression. Their results imply that overactivation of T
H17 cells and elevated CD8
+ T cell cytotoxicity may partly explain the severe immune damage in these patients alongside immune complex formation [
403]. Newer clinical trials are now starting to trial allogeneic CD45RA
− T
MEM therapy in adults affected by COVID-19 pneumonia in phase 2 trials with SARS-CoV-2 specific T cells, with early promise indicating a concurrent increase in NK cell activity (NCT04578210) [
404].
7. T cell Exhaustion in COVID-19
As above, SARS-CoV-2 infection elicits the activation of both innate and adaptive immunity [1][2]. In addition to nAbs, T cell immunity plays a crucial role in virus clearance and prevention [3]. Adaptive T cells have been reported to provide some long-term immunity against COVID-19 disease [4]. Adaptive T cell immunity consists of two major cellular populations known as CD4+ and CD8+ T cells. Antigen presenting cells (APCs) possess a critical role in activation and differentiation of these T cells. In general, infected pathogens are recognized, processed, and presented by APCs. The processed immunogenic peptides are later presented by either HLA class I or HLA class II molecules expressed on APCs surface for T cell recognition via T cell receptors (TCR). Upon activation, T cells play a vital role in pathogen clearance with CD8+ cytotoxic T cells capable of secreting a wide range of molecules such as perforins, granzymes, INF-γ and apoptotic protein Fas ligand to clear pathogen-infected cells from the host [5][6]. Meanwhile, CD4+ helper T cells play an essential role in stimulation of cytokines, proliferation of CD8+ T cells and recruitment of B cells to produce nAbs to eradicate pathogens [7]. Recent evidence shows the importance of SARS-CoV-2-specific CD8+ T cell responses in protecting against development of severe COVID-19 disease in patients [5][8]. However, persistent infection by the pathogen may result in T cell exhaustion leading to functionally ineffective and/or compromised pathogen clearance [9][10][11][12][13]. T cell exhaustion due to persistent antigenic exposure has been widely reported in many chronic diseases including human immunodeficiency virus (HIV), hepatitis C virus (HCV), hepatitis B virus (HBV), cancer, and malaria [14][15][16][17][18]. Although it is still a controversial issue on the association of COVID-19 disease with immune exhaustion, some studies demonstrate a reduction in CD4+ and CD8+ T cells in ICU-admitted COVID-19 disease severe patients [8][19][20][21]. The previous evidence shows that circulating serum isolated from these severe COVID-19 patients (n = 522) had remarkably higher levels of pro-inflammatory cytokines, especially TNF-α, IL-6, and IL-10 in comparison to those with mild-moderate patients [18][20]. The uncontrollable rise of pro-inflammatory cytokines in response to COVID-19, resulting from the release of a large amount of the cytokines in the body all at once, could be a strong indicator of “cytokine storm” that causes organ failure and death in COVID-19 [22][23]. In addition, the study also indicates that severeCOVID-19 disease patients possess higher ratio of inhibitory receptors (IR) expressing T cells [20]. T cell profiling demonstrates both CD4+ and CD8+ T cell subsets in severe COVID-19 disease patients that exhibit exhausted cellular phenotypes with remarkable expression of IR factors that include PD-1, NKG2A and TIM-3, suggesting exhausted T cells might not be able to perform viral clearance effectively in severe COVID-19 disease patients [19]. This could be indicative of exhausted CD8+ T cells, which may have lost their potential to differentiate into memory T cells which are important for sterilizing immunity [24][25]. In addition, characterization of cells from lung bronchoalveolar lavage fluid COVID-19 disease patients (n = 13) using single-cell RNA sequencing shows that patients with severe disease had relatively decreased levels of CD8+ T cells with an increased proportion of pro-inflammatory monocyte-derived macrophages and natural killer cells, indicating the importance of functional T cells in SARS-CoV-2 sterilizing immunity and COVID-19 recovery [25].