Root exudates, as an important form of material input from plants to the soil, regulate the carbon input and efflux of plant rhizosphere soil and play an important role in maintaining the carbon and nutrient balance of the whole ecosystem. Root exudates are notoriously difficult to collect due to their underlying characteristics (e.g., low concentration and fast turnover rate) and the associated methodological challenges of accurately measuring root exudates in native soils. As a result, up until now, it has been difficult to accurately quantify the soil organic carbon input from root exudates to the soil in most studies. TIn recent years, the contribution and ecological effects of root exudates to soil organic carbon input and efflux have been paid more and more attention. However, the ecological mechanism of soil organic carbon input and efflux mediated by root exudates are rarely analyzed comprehensively. In this review, the main processes and influencing factors of soil organic carbon input and efflux mediated by root exudates are demonstrated. Soil minerals and soil microbes play key roles in the processes. The carbon allocation from plants to soil is influenced by the relationship between root exudates and root functional traits. Compared with the quantity of root exudates, the response of root exudate quality to environmental changes affects soil carbon function more. In the future, the contribution of root exudates in different plants to soil carbon turnover and their relationship with soil nutrient availability will be accurately quantified, which will be helpful to understand the mechanism of soil organic carbon sequestration.
1. Introduction
The soil carbon stock is more than twice the atmosphere carbon stock and plays a critical role in the global carbon (C) cycle
[1]. Soil can sequester and store organic carbon, thereby reducing the amount of carbon in the atmosphere, and thus mitigating climate change. However, one concern is that the mineralization of soil organic carbon increases atmospheric CO
2 concentrations. Soil is a non-homogeneous system, and the temporal scales of different forms of soil carbon turnover considerably vary under the influences of climate change, soil physicochemical properties, land management, and other factors. Due to the limits in monitoring, verification, sampling, and depth methods, it is also difficult to interpret and quantify a small increase or decrease in the soil carbon stock at different spatial and temporal scales and to assess the carbon sequestration potential of soil
[2]. The source forms of soil carbon can be divided into directly fixed soil inorganic carbon (such as calcium and magnesium carbonate) and indirectly fixed soil organic carbon (SOC). SOC is a dynamic organic continuum. In order to better understand the underlying processes of SOC formation and mineralization, Lavallee et al. (2020) suggested that SOC should be separated into particulate organic carbon (POC) and mineral-associated organic carbon (MAOC)
[3]. POC is mainly derived from litter and root inputs, while MAOC is mainly microbial derivatives
[4]. MAOC usually can keep longer
[3]. With the deepening of the research on the soil carbon cycle, the mechanism of SOC stabilization has been gradually revealed. Plant carbon inputs are the dominant organic constituent of SOC, and their relative importance outweighs other abiotic factors to SOC stabilization in the topsoil at regional and global scales
[5]. Plant inputs enter the soil as either aboveground C or belowground C and include both structural litter inputs (e.g., root and shoot detritus) and dissolved organic carbon (DOC) (rhizodeposition and leaf litter leachate). A suite of recent studies has found that belowground carbon inputs have a greater impact on the efficiency and stabilization of soil carbon inputs than aboveground carbon inputs do
[6][7][8][6,7,8]. Belowground carbon inputs retained in the mineral soil are between two and five times more efficient than aboveground carbon inputs at a certain spatial and temporal scale
[9]. Plant roots are the key structure connecting the aboveground and belowground parts of plants, and their morphological structure, physiological metabolism, and other characteristics affect the carbon allocation from plants to soil.
Although soil carbon inputs from living roots account for a small proportion of total plant photosynthate compared to those from litter, they determine the asymmetry of aboveground and belowground SOC distribution and are more effectively retained in MAOC and POC than litter carbon inputs
[6]. However, the release of organic compounds into the soil by living roots has been the most uncertain part of the soil carbon flux and cycle
[10]. Root carbon inputs can lead to SOC increase and promote SOC stabilization. For example, compared with plant shoot and mycorrhizal pathways, plant roots in temperate steppe contribute to the newest SOC formation under different soil fertility levels
[11]. However, root carbon inputs can also reduce SOC stabilization so that preserved C can be used by soil microbes, leading to SOC loss. Meanwhile, plant roots can also reduce the stability of organic carbon through the destruction of aggregates
[12]. Therefore, it is very important to identify the mechanism of root carbon inputs on soil carbon stabilization and to evaluate SOC input and efflux with precise quantification. In the root system, fine roots have been the focus of most studies, because they are generally essential for many root functions (e.g., root elongation, nutrient and water acquisition, association with symbionts, and carbon exudates). The roots of grassland plants form a dense network of fine roots in the deeper soil and can alleviate the decomposition of SOC to some extent
[13]. The links among fine root classes (including individual root orders), traits, and functions will be a key research area, as morphological and chemical traits of fine roots have been extensively studied, while physiological traits have rarely been considered
[14].
Rhizodeposition is formed when plant roots release organic and inorganic compounds in the roots to the adjacent soil, which mainly comes from plant detritus, the decomposition and renewal of root cells, and root exudates. Gross rhizodeposition carbon accounts for 5–20% of photosynthates
[15]. Root exudation can account for up to 2–11% of the total photosynthetic production
[16]. Root exudates are an important part of rhizodeposition, reflecting a physiological property of fine roots, and some dissolved organic materials in root exudates produced by living plants through primary metabolism, secondary metabolism, and abiotic metabolism are the main carriers for plant roots releasing organic carbon into the soil
[17]. CO
2 in the atmosphere that is fixed by plants through photosynthesis reaches the soil via aboveground plant residues, belowground root exudates, and root turnover, and these constitute the main sources of soil carbon. The contribution rate of root exudates and litter to soil organic carbon input in different degraded grassland ecosystems in Inner Mongolia ranged from 8.8 to 14.8% and 16.8 to 17.2%, respectively
[18]. While interacting with biotic and environmental factors, root exudates also affect SOC efflux
[19]. Most of
theour previous studies focus on root exudates of the crop plant (bean, wheat) to develop agriculture and the interaction between root exudates and the rhizosphere environment to reveal the relationships between root exudates and soil carbon and nitrogen. With the increasing development of basic research content and analytical methods, soil chemistry and ecology have gradually become the cross-content of root exudates research, and the research topic has gradually become ecological problem mechanisms from the application
[20]. The processes of the soil carbon cycle mediated by root exudates and its influencing mechanism are key and difficult points in the construction of the global carbon cycle model
[21]. In many studies, the physiological characteristics and ecological effects of root exudates have been summarized
[22][23][22,23], and their response to stress in the environment and feedback role
[24][25][24,25] and the influence of root exudates on SOC input and carbon efflux have also been mentioned
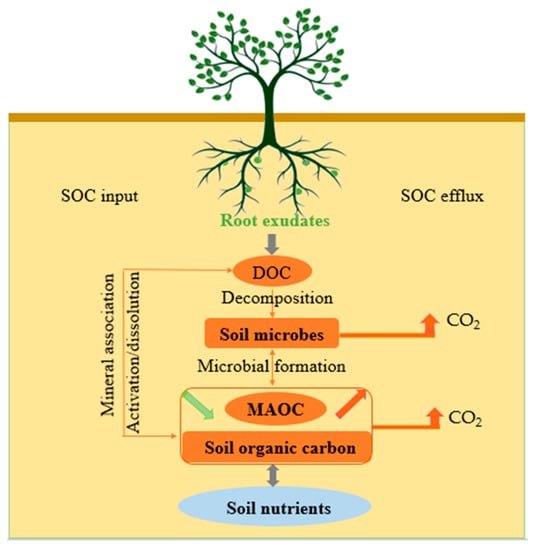
[12][26][12,26], but few studies have integrated SOC input and efflux to explore root exudates. In order to quantify the contribution of abiotic factors (e.g., physical and chemical factors) and biologic factors (e.g., microbial community factors) to soil carbon mineralization and stability mediated by root exudates, and to clarify the potential mechanism of root exudates in soil carbon sequestration, fully understanding the processes of SOC input and efflux mediated by root exudates and their effect mechanism is important. The processes of SOC input and efflux mediated by root exudates are illustrated and the mechanism of root exudates on SOC sequestration is revealed in this paper (
Figure 1).
Figure 1.
Main processes of soil organic carbon input and efflux mediated by root exudates.
2. Root Exudates and Soil Organic Carbon Input
The interactions of root exudates with soil microbes and minerals are beneficial to SOC formation. Active compositions in root exudates effectively form MAOC by combining with organic minerals in the surrounding mineral soil
[27]. Unstable DOC (such as monosaccharides) compounds in root exudates provide soil microbes with important precursors involved in SOC formation
[28]. Firstly, there is spatial heterogeneity in SOC formed by root exudates in the rhizosphere. The effects of dissolved matter in root exudates on SOC storage and structural development are significantly different between surface and bottom soils, and the potential for SOC accumulation in deeper soil is higher than in surface soil due to the addition of root exudates. In forests, a large number of dissolved matter in root exudates can induce large aggregation of organisms and storage of carbon in deeper soil
[29]. Secondly, the composition complexity of root exudates affects the formation of different soil carbon components. Simple compositions in root exudates may promote the formation of MAOC, while complex compositions may promote the formation of POC
[30]. So, the same compositions of root exudates have different effects on SOC stability in different spaces, and the different quantities and compositions of root exudates have different effects on soil carbon turnover.
The quantity and quality of root exudates vary significantly among plant species
[31]. Phylogeny is a common controlling factor of root functional traits and root exudates
[32]. In recent years, root exudates have been gradually taken into account in the construction of multi-dimensional traits and economic spaces to study the functional mechanisms of roots. Root functional traits can reflect the characteristics of root morphogenesis, chemical metabolism, and symbiosis. For example, specific root length is commonly known as specific root area and is a compound trait determined by root diameter (RD) and root tissue density (RTD), which reflects the cost of root construction; root nitrogen concentration (RNC) is used to reflect the overall metabolic activity of roots; and mycorrhizal colonization represents the symbiotic character of plants. All these traits reflect the strategies and efficiency of plant uptake of soil nutrients
[33]. The root exudation rate has been shown to have different correlations with root functional traits in different species, and it can be used as a functional trait and a key indicator of the plant resource utilization strategy (utilization and conservation)
[34]. For instance, root exudates from the fine roots of woody plants are positively correlated with root N concentration
[35]. Some studies also report that the specific root exudation rate increases with the increase in root diameter and decreases with the increase in RTD, but it has no relationship with RNC
[36]. Evidence from studies on root exudates suggests that the root exudation rate is a survival strategy for plants to adapt to a low-nutrient environment, and an increase in the root exudation rate leads to a high cost of root construction and slow root growth
[35]. There is a negative correlation between mycorrhizal colonization and the root exudation rate, indicating that there is a trade-off between mycorrhizal symbiosis and root exudates in belowground carbon allocation
[37]. Based on the analysis and research on the characteristics of root exudates, the quality of root exudates (metabolic characteristics) is considered to play a more important ecological role in root functional traits. RD, RTD, and RNC change obviously in the metabolic composition of root exudates
[38]. The composition of primary metabolites in root exudates varies with the plant growth strategy, and the root of more competitive species (faster-growing species) secretes more primary metabolites (e.g., sugars, organic acids), while the root of conserved species (slower-growing species) secretes more amino acids
[34]. Therefore, root functional traits determine the carbon allocation of root exudates and nutrient utilization strategies of plants in different soil stocks, and the relationship between root exudates and root traits mainly depends on plant characteristics.
3. Root Exudates and Soil Organic Carbon Efflux
It is generally believed that the increase of SOC input can effectively promote soil carbon sequestration, but more and more evidence shows that SOC input will also change the turnover of the original SOC, thus resulting in SOC loss. Root exudates, as an input form of active C substrate, can stimulate the decomposition of more stable SOC
[39], which is a process known as the “priming effect”. One mechanism is the “Microbial metabolism theory”, in which root exudates, as the energy matrix of soil microbes, stimulate the decomposition of SOC by affecting the structure, composition, and activity of microbial communities. Another new mechanism is that root exudates (oxalic acid) disrupt or reduce the stability of the MAOC complex by physical chemistry, thus allowing the release of protected C from the soil for microbial decomposition and utilization
[19]. There is no consensus on the mechanism of the rhizosphere priming effect. Although the priming effect has been proven to be prevalent in nature, most relevant studies are currently conducted in greenhouses. It is indeed necessary to strengthen field experiments as well as long-term observation and data accumulation
[39]. Frequent carbon inputs can induce a stronger priming effect than occasional carbon inputs do, so increasing carbon input frequency may weaken the carbon sink of grasslands
[40]. This means that the priming effect is likely to offset the contribution of root exudates to SOC input, and the involvement of root exudates in the formation of SOC does not improve the carbon sequestration potential of soil to some extent as a whole, but only provides a short-term prediction.
The quantity and composition of root exudates are different and make the intensity of the priming effect different. Higher rhizodeposition can accelerate soil carbon turnover
[41]. The simpler the structure of the root exudates C matrix, the faster the energy conversion, the more complex the structure of the root exudates C matrix, and the less the priming effect
[42]. Plant functional traits are key factors for the priming effect to affect SOC decomposition. An increase in the root exudation rate will increase the influence of the priming effect on organic matter decomposition
[43]. Due to differences in the strategies and efficiency of using C by different soil microbes, the distribution and turnover of root exudates in different soil microbial groups are different, and different bacterial functional groups in the soil have different effects on the priming effect. According to the relative distribution and turnover rate of root carbon deposition in the individual microbiome in the soil, the mean turnover rates of rhizodeposited C in microbial groups ranged from 0.04 to 0.13 day
−1. Gram-negative bacteria and fungi were the dominant microbiomes that converted root C deposition into SOC
[15].
The stability and activity of SOC also affect the intensity of the priming effect. SOC stability (characterized by chemical resistance and physical chemistry protection) can better account for differences in the priming effect than the plant, soil, and microbial characteristics
[44]. The priming effect of soil carbon increases with an increase in soil refractory carbon components but decreases with an increase in soil aggregate and mineral protection
[45]. The relatively more stable SOC has a higher priming effect than the unstable SOC does. The rapid turnover of root exudates mainly depends on material decomposition and microbial utilization. Root exudate decomposition is an important part of the carbon cycle in terrestrial ecosystems. Most rhizodepositions, especially active C in root exudates easily used by soil microbes, have an important impact on soil C dynamics
[46]. However, root exudates such as oxalate may also release organic compounds directly from organic mineral aggregates, so that soil microbes access these compounds easily, resulting in increasing net soil carbon loss
[47]. The part of the carbon released by microbial decomposition in the priming effect is not released into the atmosphere, and this part of carbon can compensate for the carbon loss caused by the priming effect, but where the carbon goes in the soil is still uncertain
[48]. Some studies have quantified the decomposition rate of SOC in the rhizosphere of grasslands and their responses to drought and nitrogen addition using
13C-CO
2 pulse labeling. The results show that the decomposition amplitude of accumulated rhizodeposition C was similar to that of accumulated SOC. Decomposition of rhizodeposited C accounted for 7–31% of the total below-ground respiration in grassland ecosystems. It may be a larger component of CO
2 released from the soil in grasslands
[49]. Therefore, it is necessary to further improve the understanding of the influence of abiotic factors on the intensity and direction of the priming effect.