1. Introduction
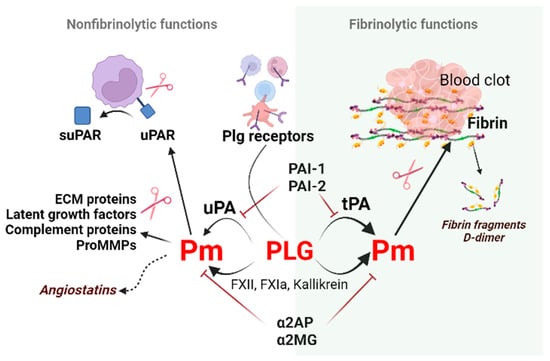
The primary function of the plasminogen (Plg/Pm) system is to ensure the destruction of fibrin deposits and maintain hemostatic balance. Hemostasis is the process of blood clot formation at the site of vessel injury. There is a careful balance between thrombin-stimulated clot formation and plasmin-induced clot lysis. Abnormal bleeding occurs when there is insufficient clot formation due to decreased thrombin (e.g., from F VIII deficiency) or increased clot lysis. Conversely, non-physiological thrombosis or functional clotting occurs when excessive thrombin production or impaired clot lysis is present. During the initial phase of hemostasis after tissue damage, endothelial injury induces platelet aggregation within 10–20 s. The sequential activation of coagulation factors (XIII-II) generates an initial hemostatic plug in 1–3 min. Fibrin, produced at the end of the coagulation cascade, adds to the clot stability by 5–10 min. Since a clot should only obstruct the vessel for a particular time, the body switches from clotting to antithrombotic control mechanisms. The activation of plasmin that leads to fibrin degradation and the occurrence of fibrin degradation fragments, a process called fibrinolysis, ultimately removes the clot to ensure tissue blood reperfusion (Figure 1).
Figure 1. Non-fibrinolytic and fibrinolytic functions of the plasminogen/plasmin system. Binding and subsequent plasminogen activation via plasminogen receptors (Plg-RKT, annexin 2, actin, etc.) generate plasmin. Plasmin induces proteolytic activity on the cell surface to cleave the extracellular matrix molecules necessary for cell migration. Plasmin generated by tPA on polymer fibrin, uPA, or clotting factors on activated cell surfaces dissolves fibrin and produces fibrin fragments such as D-dimers. This process is called fibrinolysis. The non-fibrinolytic functions of plasmin include proteolytic activity towards latent growth factors, the complement component C5, and pro-MMPs resulting in changes in molecule-linked signaling pathways. The cleavage of plasmin, elastase, and MMPs generate angiostatins–kringle-containing plasminogen fragments possessing anti-angiogenic properties. Abbreviations: Pm, plasmin; tPA, tissue-type plasminogen activator; PAI-1, plasminogen activator inhibitor-1; ECM, extracellular matrix; MMPs, matrix metalloproteinases; suPAR, soluble urokinase plasminogen activator receptor; α2AP, α2-antiplasmin; α2MG, α2-macroglobulin.
Besides the fibrinolytic function, the Plg/Pm system can alter cell receptors, extracellular matrix molecules, or growth factors due to its proteolytic capacity. In addition, components of the Plg/Pm system participate in intracellular signaling processes, thus regulating tissue development and remodeling during wound healing, inflammation, trophoblast invasion, angiogenesis, tumor growth, etc.
[1][2][3][4][5][1,2,3,4,5]. These functions are referred to as non-fibrinolytic functions (
Figure 1).
There are two major plasminogen activators (PAs) that can activate Plg in humans: tissue-type plasminogen activator (tPA) and urokinase plasminogen activator (uPA). Both PAs are serine proteinases of the trypsin type. However, differences in their structure cause functional differences: tPA mainly converts Plg to plasmin on fibrin clots, while the activity of uPA is regulated by its interaction with the cell surface
[6][7][6,7]. Plg activation is involved in tissue remodeling and inflammatory response (reviewed in
[8][9][8,9]). Plg levels are high at the inflammation site and regulate the wound-healing process by activating the early inflammatory reaction
[10] by increasing inflammatory cell infiltration.
“Health comes first, and it enters through the mouth”. The mouth, in academic terms, is referred to as the oral cavity
[8]. The oral cavity is a structure of tissues and organs with complex organization and interaction. Initial parts of the digestive system, the immune system (Waldeyer’s tonsillar ring), and sensory-specific and non-specific receptors (taste, temperature, pain, tactile sensation, etc.) are located in the oral cavity. Both the nasal cavity and nasopharynx, as the beginning of the respiratory system, and the oral cavity are the entrance gateway for bacterial and viral infections. The first signs and symptoms of general infectious and non-infectious diseases can occur in the oral cavity (loss of taste or hemorrhagic rash on soft palatine mucosae in COVID-19
(coronavirus disease 2019), Koplik’s sign in measles, oral dryness and the decaying of multiple teeth in diabetes mellitus, etc.). The oral cavity environment is very sensitive to systemic and local changes in the organism and reacts to pathogen exposure with quantitative and qualitative changes in saliva and crevicular fluid. The Plg/Pm system of the oral fluid plays an integral part in the physiological regeneration and protective mechanisms of oral mucosae, the pathogenesis of several inflammatory or autoimmune diseases, and tumor growth in the oral cavity
[11].
2. Regulators of Fibrinolysis
The Plg/Pm system includes the proteolytic enzyme plasmin and its inactive precursor Plg, its activators (tPA and uPA), plasmin inhibitors (α2-antiplasmin and α2-macroglobulin), and inhibitors of plasminogen activators (PAI-1 and PAI-2)
[12]. These components regulate and interact with each other as well as with clotting system components, cell receptors, and pericellular adhesion molecules (
Figure 1)
[5]. In addition, many proteins can modulate the activity of the fibrinolytic system components: proteins such as vitronectin, thrombospondin, tetranectin, and histidine-rich glycoprotein, which can bind to Plg/Pm, fibrinolysis activators, or inhibitors.
PAs and components of the clotting cascade mediate Plg conversion to active plasmin. The tissue activator generates plasmin on fibrin and cell surface. Activation of Plg by uPA occurs on the cell surface in blood circulation or outside of it
[6][12][6,12]. Like thrombin, plasmin activates protease-activated receptor (PAR)1 and PAR2, modulating platelet activation, the release of proinflammatory cytokines by immune cells, and endothelial function
[13].
2.1. Plasminogen (Plg)
The inactive proenzyme Plg and its active derivative plasmin are key components of the fibrinolytic system. Plasmin belongs to the serine protease family of enzymes. Plg is secreted into the bloodstream mainly by hepatocytes and the kidney epithelium and can be synthesized and utilized out of circulation in the cornea
[14]. Interleukin (IL)-1a and -1b can stimulate the extrahepatic synthesis of Plg in the human cornea
[14].
Plg is a 92 kDa glycoprotein comprised of heavy (N-terminal domain followed by five kringle domains) and light (protease domain) chains linked by an activation loop
[15]. Two glycosylated forms of Plg (I and II) vary in their number of sugar remnants, their affinity to fibrin, and their activation susceptibility
[16]. Substrate or receptor binding leads to the dissociation of intramolecular bonds and proenzyme transition into an open form susceptible to activation
[17][18][17,18]. Proteolytic fragmentation of the Plg/Pm molecule produces kringle-containing fragments such as angiostatins
[19][20][19,20]. Angiostatins bind to plasminogen receptors or the hepatocyte growth factor receptor c-met. This binding results in the blockade of proliferation or angiogenesis and the induction of apoptosis
[21].
α2-antiplasmin and α2-macroglobulin
[22][23][22,23] are natural plasmin inhibitors (
Figure 1).
Proteolytically active plasmin has a broad specificity. It modulates the release and activation status of growth factors/cytokines (e.g., TGF-β, fibroblast growth factor-2
[24][25][24,25], hepatocyte growth factor
[26], insulin-like growth factor-binding protein 4, and IL-1β
[5]), hormones (e.g., prolactin
[27], lactogen, osteocalcin
[28], pro-opiomelanocortin
[29], proinsulin
[30][31][30,31]), receptors (e.g., uPAR
[32] and EPH receptor A4), and proteases (e.g., tPA, uPA, and MMPs
[33][34][33,34]) (
Figure 1).
Plg and plasmin activators binding to annexin A2, urokinase plasminogen activator receptor (uPAR), and other docking sites colocalize enzyme and substrate, generating efficient plasmin at cell surfaces. Plg receptors can mediate the fibrinolytic function of this system and signal transmission inside the cell. Among the receptors for plasminogen on the cell surface are the highly specific Plg-RKT (plasminogen receptor with a C-terminal lysine) and the less specific αIIbβ3-integrin, αMβ2, αVβ3, α-enolase, gamma-actin, S100A10, annexin 2, histone H2B, amphoterin, or PAR. Aside from receptors, Plg interacts with partner proteins, such as fibrinogen/fibrin or tetranectin on cell surfaces
[22][35][22,35]. Plg-RKT is expressed on monocytes, macrophages, and neuronal cells. It is sterically close to uPAR, providing conditions for plasmin generation and the Plg/Pm-dependent inflammatory response
[36][37][36,37]. At the same time, annexin A2 reduces plasmin production and facilitates the autoproteolytic destruction of plasmin
[38].
2.2. Plasminogen Activators (PAs)
tPA activates Plg mostly on fibrin thrombi surfaces but also on some cell membranes, mediating plasmin formation for cell movement through the extracellular matrix and modulating cell signaling. uPA mainly acts on cell surfaces. However, recent research has demonstrated the involvement of both PAs and plasmin in cell signaling, migration, and extracellular matrix remodeling
[5][39][5,39]. Factor XIIa, an endogenous activator of the clotting system, can activate Plg and kallikrein and convert single-chain urokinase into double-chain urokinase. Nevertheless, their role in physiological fibrinolysis is considered insignificant
[40].
tPA: tPA (tissue-type plasminogen activator,
Plat gene) is a 70-kDa glycoprotein belonging to the serine protease family. tPA is synthesized mainly by endothelial cells, but mesenchymal cells, monocytes, smooth muscle cells, and fibroblasts can also produce it
[41]. It is the primary PA (>90%) in all tissues except the kidney and liver (65%) and the spleen (40%). Lung tissues yield the highest tPA activity, followed by kidney, brain, heart, adrenal, liver, aorta, spleen, and muscle tissue
[42]. In addition, stress, adrenergic stimulation, the diurnal cycle (and other circadian cycles), histamine, and thrombin enhance the synthesis and release of tPA. Most tPA in circulation exists in a complex with its primary inhibitor, PAI-1. tPA has a short half-life (3–4 min) and is removed from the bloodstream by the liver via mannose receptors
[41][43][41,43]. The serine proteinase domain of tPA has a narrow specificity for Plg activation. In addition, the PAI-1 binding site is located in the serine proteinase domain of the tissue activator
[44][45][46][44,45,46].
Besides having fibrinolytic functions, tPA can modulate cell signaling due to its ability to activate Plg on the cell surface or interact with specific receptors (reviewed in
[8]). tPA-related signaling accelerates ischemic revascularization and regulates synaptic plasticity, blood-brain barrier permeability, cytokine production, cell proliferation, etc. tPA can affect cell fate alone or as part of the PAI-1/tPA complex
[47]. The binding of tPA to Annexin A2 enhances proinflammatory cytokine production by macrophages through the generation of active plasmin and subsequent matrix metalloprotease-9 activation
[48]. In addition, it stimulates endothelial progenitor cell evasion from the bone marrow
[49].
The interaction of tPA with the low-density lipoprotein-related protein-1 (LRP1) receptor triggers cell survival and proliferation
[39]. After binding to the membrane receptor LRP1, tPA-mediated NF-κB activation increases the expression of proinflammatory chemokines in macrophages
[50]. tPA triggers a cascade of survival signaling involving extracellular signal-regulated kinase 1/2
[51]. In macrophages, enzymatically non-active tPA inhibits toll-like receptors through the N-methyl-D-aspartate receptor
[52].
uPA: uPA (
Plau gene) is a 54-kDa glycoprotein, synthesized as a single chain (sc-uPA) and converted into a two-chain uPA (tc-uPA) by plasmin and kallikrein
[41]. The uPA molecule contains a protease, EGF, and a kringle domain without a lysine binding site which ensures that it cannot bind to fibrin
[53].
Further proteolysis of sc- or tc-uPA by plasmin or matrix metalloproteases (MMPs) generates an amino-terminal fragment (ATF) that consists of the EGF and kringle domain (33 or 32 kDa)
[54]. The ATF can bind to the primary uPA receptor called uPAR. Thrombin cleaves the Arg156–Phe157 peptide bond near the active site and generates another proteolytic two-chain inactive uPA
[55]. Recent studies have demonstrated that soluble uPAR (suPAR) dimers, but not monomers, have a stronger binding ability to the ATF of uPA
[56].
2.3. Plasminogen Activator Inhibitors
Fibrinolysis shutdown is provided by plasmin inhibitors, plasminogen activator inhibitors, and indirect fibrinolysis inhibitors.
α2-antiplasmin and
α2-macroglobulin
[22][23][22,23] are naturally occurring specific plasmin inhibitors (
Figure 1). Fibrin-bound plasmin and receptor-bound plasmin are protected from inactivation by plasmin inhibitors
[7]. Indirect inhibitors (e.g., thrombin-activated fibrinolysis inhibitor, TAFI) regulate the rate of clot dissolution by fibrin modification
[57]. However, the most abundant fibrinolysis inhibitor in circulation is PAI-1, a glycoprotein that belongs to serine protease inhibitors (SERPINs) and is, therefore, also called serpin E1. PAI-1 can be found in blood, soft tissues, tissues of the parenchymal organs, vessel walls, nervous tissue, etc.
[58].
The primary function of PAI-1 in the bloodstream is to inhibit tPA and uPA, thus regulating the rate of fibrinolysis and the duration of blood clots. In tissues, PAI-1 also performs a signaling function and participates in the regulation of cell migration
[59]. PAI-1 is synthesized in the liver and spleen epithelium, adipocytes, hepatocytes, platelets, megakaryocytes, macrophages, smooth muscle cells, and placental and endothelial cells
[58]. In addition, pathological conditions can enhance PAI-1 expression in plasma (2–46 ng/mL) and other tissues. PAI-1 expression and release by cells such as platelets are regulated by various factors: growth factors (e.g., transforming growth factor-β (TGF-β), EGF
(epidermal growth factor), PDGF
(platelet-derived growth factor), tumor necrosis factor (TNF)-α, and interleukin-1 (IL-1)β), hormones (e.g., insulin, glucocorticoids, and angiotensin II), the glucosides and endotoxins of gram-negative bacteria, low-density lipoprotein, and very low-density lipoprotein
[60].
PAI-1 is a single-chain glycoprotein with a mass of 47–50 kDa
[61]. PAI-1 exists in several conformational forms. In addition to the active and inactive forms, there is also a latent form. PAI-1, synthesized as an active enzyme, is spontaneously transformed into an inactive form with a half-life of 1 to 2 h. PAI-1 is the only serpin that can perform a reverse conformational transition between active and latent states. In plasma, the active form of PAI-1 can be stabilized by binding to vitronectin, thereby increasing its half-life in the bloodstream. On the other hand, PAI-1 activity rapidly decreases at lower pH, as has been found in ischemic tissues
[59].
3.3. Soluble uPAR, a New Biomarker of Inflammation
3.3. Soluble uPAR, a new biomarker of inflammation
uPAR (CD87; Plasminogen Activator, Urokinase Receptor [Plaur gene]) is a membrane-linked protein found in immunologically active cells (monocytes, neutrophils, activated T lymphocytes, macrophages), endothelial cells, keratinocytes, fibroblasts, smooth muscle cells, megakaryocytes, and certain tumor cells. Apart from uPA, uPAR can also interact with integrin or other partners, including vitronectin, high molecular weight kininogen, G protein-coupled receptor, tyrosine kinase receptors that can trigger plasmin generation and degradation of the extracellular matrix (ECM) along the leading edge of a migratory cell [62] and activate downstream signaling pathways. uPA via plasmin can increase cell proliferation through the proteolytic activation of growth factors and adhesion molecules, remodeling of tissues/ECM, regulate adhesion and invasion of normal and cancer cells [63][54].
Cleavage of uPAR at the GPI (glycosyl phosphatidylinositol) ananchor by proteases like plasmin can shed the extracellular part of uPAR, releasing the soluble form of the receptor (suPAR) into the blood, mucosa, urine, and saliva [64][65].
3. Fibrinolytic Ffactors Dduring Iinflammation in the Oral Coral cavity
Lined with keratinized and non-keratinized stratified squamous epithelium, oral mucosa covers the oral cavity. It is moistened with excretes of the major parotid, submandibular, sublingual, and minor salivary glands within the oral cavity. Mechanical mucosal trauma occurs while eating, drinking, and talking (and even tobacco inhalation). Oral mucosa is a first-line defense that interacts with pathogens (e.g., bacteria, viruses, or fungi) and provides specific (immune) or non-specific protective response against pathogenic microorganisms such as pattern recognition receptors including C-type lectin receptors (Dectin-1, Dectin-2) or TLR1-1 (TLRs, toll-like receptors). Gradual desquamation of mucosal epithelium is a protective mechanism to eliminate adherent pathogenic microorganisms and to prevent their further invasion into underlying tissues [65][74]. A host organism reacts with the release of proinflammatory cytokines and proteases to fight oral microorganisms in the gingiva and periodontal ligament space.
Salivary glands provide local mucosal-specific and non-specific immunity. Proper qualitative and quantitative composition of saliva and salivation rate protect and maintain the integrity of the oral cavity. Salivary mucins avert plaque formation on teeth surfaces via bacteria binding. Salivary lysozyme, an enzyme that lyses bacteria cell walls prevent the overgrowth of oral microbiota. Several studies are available regarding the fibrinolytic properties of salivary glands and other fluids of the oral cavity [66] [75] during steady state and stress/inflammation.
Salivary suPAR, tumor necrosis factor α (TNF α), and interleukins like (IL)-1β levels increased in healthy subjects exposed to psychological stress [76] and showed strong positive baseline and post-stress correlations. Elevated saliva suPAR levels were detected and proposed as a biomarker of gingivitis and periodontitis [67][68][78,79]. Aside from suPAR, Plg receptors such as glyceraldehyde-3-phosphate dehydrogenase, α-enolase, and annexin A2 are also found in the saliva [69][70][71][72][81-84], but their function in the oral cavity remains unclear. Excretory ducts and acinar cells highly expressed PAI-2. tPA was expressed in serous but not mucous acini [66][75]. Overall, several independent studies confirmed the existence of tPA in saliva, but the results of different investigators varied on the presence/dominance of PAI-1 or PAI-2 in salivary gland tissues. PAI-1 increases in saliva were associated with insulin resistance and inflammation and ascribed as a proinflammatory marker and valuable diagnostic marker to track periodontal therapy [73][88].
Major and minor salivary glands, oral mucosae, and periodontium play an important role in maintaining oral cavity Plg/Pm system balance. Oral cavity chronic diseases and hyposalivation cause a dysbalance of the Plg/Pm system in the oral cavity, supporting inflammation. Viruses, including SARS-CoV2, enter the body via the oral and nasal cavities, where the initial replication of viruses occurs. The non-fibrinolytic proteolytic function of plasmin supports the initial stages of infection with SARS-CoV-2 (severe acute respiratory syndrome coronavirus 2). For example, the SARS-CoV-2 virus can enter the buccal epithelium after binding its S protein to ACE2. Plasmin cleaves the S-protein of the SARS-CoV-2 virus and ENaC (epithelial Na+ channel) α, β, γ, and facilitates SARS-CoV-2 infection into susceptible cells (non-fibrinolytic function).