Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Rita Xu and Version 1 by Francesca Lionetto.
Fish collagen garnered significant academic and commercial featuring prospective applications in a variety of health-related industries, including food, medicine, pharmaceutics, and cosmetics. Due to its distinct advantages over mammalian-based collagen, including the reduced zoonosis transmission risk, the absence of cultural-religious limitations, the cost-effectiveness of manufacturing process, and its superior bioavailability, the use of collagen derived from fish wastes (i.e., skin, scales) quickly expanded. Moreover, by-products are low cost and the need to minimize fish industry waste’s environmental impact paved the way for the use of discards in the development of collagen-based products with remarkable added value.
- fish collagen
- fish industry waste
- collagen extraction
1. Introduction
In order to exploit natural resources as much as possible, a long-term plan titled “Blue Growth” was approved by the European Commission and has been implemented to pay particular attention to fish resources in order to preserve the environment from industrial pollution. The enormous amount of valuable protein that could be extracted [1,2,3,4,5][1][2][3][4][5] (about 30–40% of the total weight), is one of the most appealing aspects of seafood by-products. More than 20 million tons of them are produced annually from the fish tissues that are discarded as waste, including fins, heads, skin, and viscera [6,7,8][6][7][8]. Because of their elevated protein content, absence of disease transmission risks, high bioactivity, and less considerable religious and ethical restrictions, the use of fish by-products as a new source of collagen has drawn increasing attention [9,10,11][9][10][11].
The importance of both aquaculture and fishing to food security is expanding continuously, particularly in light of the rising global fish production and the United Nations’ 2030 program of sustainable development [12]. Approximately 70% of fish and other seafood are processed before being sold, resulting in enormous amounts of solid waste from processes such as beheading, de-shelling, degutting, separating fin and scales, and filleting [13,14][13][14]. More than half of the weight of fresh fish becomes by-products of the fish industry. Most of these by-products are buried or burned, causing environmental, health, and economic issues. A minor portion are employed as inexpensive ingredients in animal feeds. Fish waste is a rising problem that requires quick, creative methods and solutions. Numerous initiatives and programs have been performed globally to prevent food waste. In addition to reducing the cost of waste disposal, investing in waste from the fish industry can offer the opportunity to recover other important substances such as oils, proteins, pigments, bioactive peptides, amino acids, collagen, chitin, gelatin, etc. [15,16,17][15][16][17].
More than two decades ago, research on the extraction of collagen from fish waste started to be conducted. Collagens are one of the most abundant proteins in animals, which are found in the extracellular matrix of connective tissues, including skin, bones, tendons, ligaments, cartilage, intervertebral discs, and blood vessels [18]. Collagens are not only implicated in tissue architecture maintenance and strength, but they also cover regulatory roles (i.e., through mechano-chemical transduction mechanisms) during tissue growth and repair [19,20][19][20]. Thanks to their nature, collagens are intrinsically bioactive, biocompatible, and biodegradable [21]. Hence, collagens are valued as the most commonly required and used biomaterials in many fields, including medical, cosmetic, nutraceutical, food and pharmaceutical industries in the forms of injectable solutions, thin substrates, porous sponges, nanofibrous matrices, and micro- and nano-spheres [22,23,24,25][22][23][24][25]. Recent studies revealed many similarities in the molecular structure and biochemical properties between collagen derived from fish and mammalian sources, despite the fact that fish collagen typically has a lower molecular weight and lower denaturation temperature than mammalian collagen [8,12,20,22,24,26,27,28][8][12][20][22][24][26][27][28]. Various extraction techniques for fish collagen have been developed depending on the selected tissue type and fish species. Hence, a considerable collection of literature has been developed on this subject [29,30,31][29][30][31]. Only in the past five years have researchers concentrated on innovative materials with improved characteristics in addition to developing extraction techniques for mass manufacture.
Collagen nanotechnology has a bright outlook because science in this area is always progressing and will continue to do so in the future. Nano collagen is ordinary collagen that has been sized down to a nanometer scale [32,33][32][33]. According to its nano-scale-based technology, which offers a high surface-area-to-volume ratio, an optimal penetration into wound sites and higher cell interaction is enabled. [34]. Moreover, nano collagen has the ability to deliver drugs and to supply a durable microenvironment at wounded sites to promote cellular regrowth and healing [35]. Collagen nanotechnology still presents many shortcomings, including the fact that only a small minority of therapeutic compounds have received commercial approval and that there are still numerous unsolved problems [32]. The complexity of pathophysiological symptoms and the lack of data on its real physiological effects is a further challenge for nanotechnology. Despite these downsides, nanotechnology is still a growing trend, with a huge amount of unrealized potential. This gives rise to the expectation that further research will assist in minimizing these downsides, leading to the creation of secure and efficient nano-based systems. In order to create approved therapeutic agents that take advantage of nanotechnology, additional research and studies must be performed [32]. Indeed, Figure 1 reveals the continuous increasing research interest on collagen, fish collagen, and nano collagen investigation in the last twenty years. In particular, it appears clear that there has been a significant increase in scientific works in the last five years. Nano collagen can be used for a variety of improvements and treatments, such as bone grafting, drug delivery, nerve tissue formation, vascular grafting, articular cartilage regeneration, cosmetics, and wound healing [21,22,36][21][22][36]. It is clear that nano collagen is a progressed type of nanotechnology; thus, further investigation must be attempted to advance this technology with the expectation that, in the future, nano collagen scaffolds will be more widely available [37].
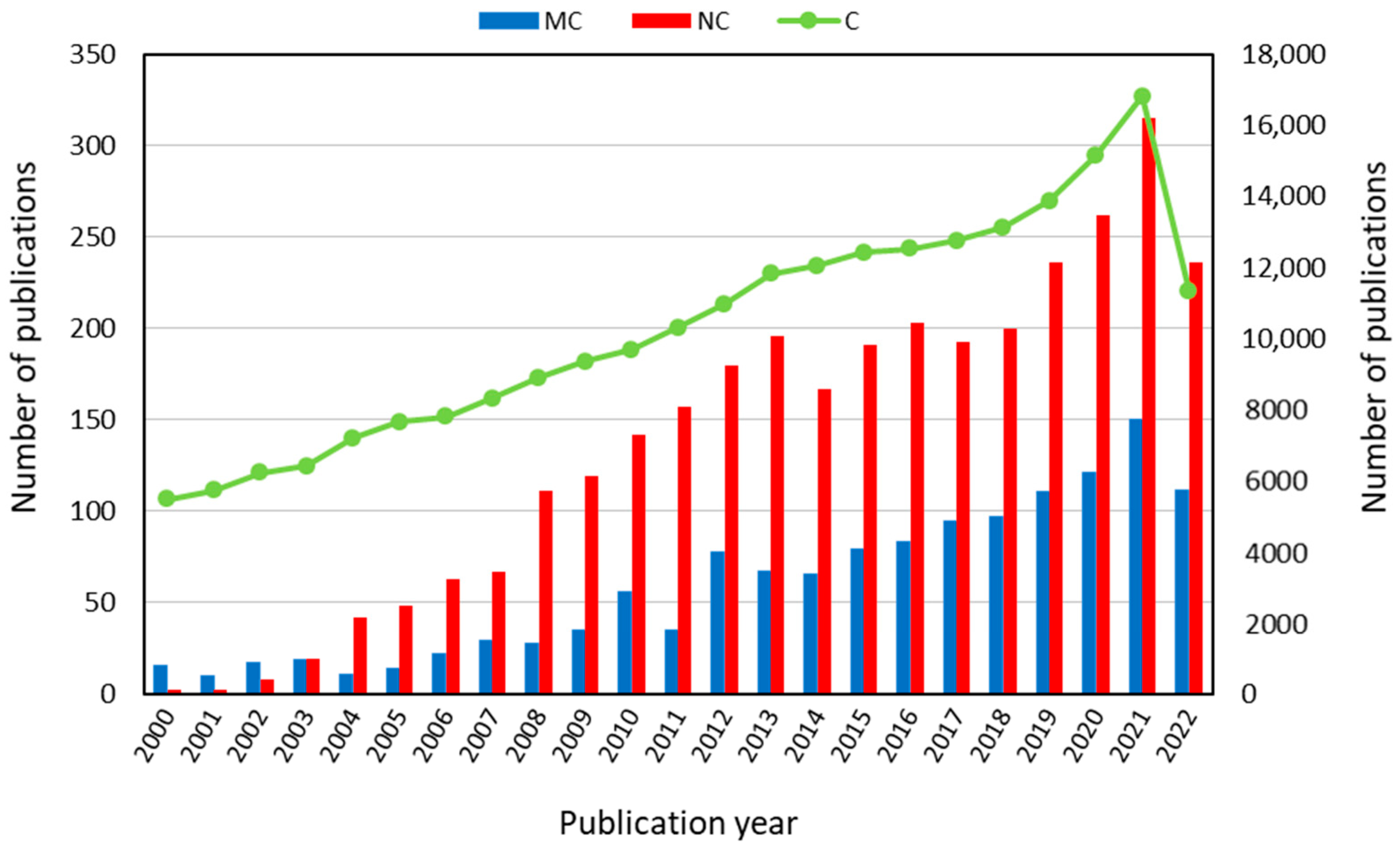
Figure 1. Increasing research interest in fish collagen (MC) and nano collagen (NC) compared with collagen (C), according to scientific papers analyzed by publication year in the last twenty years up to 2022 (from Scopus database: www.scopus.com, accessed on 15 September 2022).
2. Collagen: Structure and Properties
Collagens represent about 30% of a mammal’s weight [18,38][18][38]. Based on the historical order of their discovery, 28 types of collagens—type I through type XXVIII—have been identified and described up to the current day [39]. The oldest collagen identified to date was found in the soft tissue of a fossilized Tyrannosaurus rex bone that dates back 68 million years [40,41][40][41].
The molecular organization of collagens is highly variable, notwithstanding their general triple-helical structure and the triplet (Gly-X-Y)n repetition, where X and Y can be any amino acid, although proline and hydroxyproline are the most frequent occupants of these locations (Figure 2) [42,43][42][43]. Collagen’s unit is composed of three α-chains, the amino acid composition of which varies among collagen types. Furthermore, function and distribution in tissues play a role in the diversity of collagen, as well as molecular and supramolecular organization, such as occurrence and length of triple helical domains, packing of the triple helices, etc. [27,44][27][44].
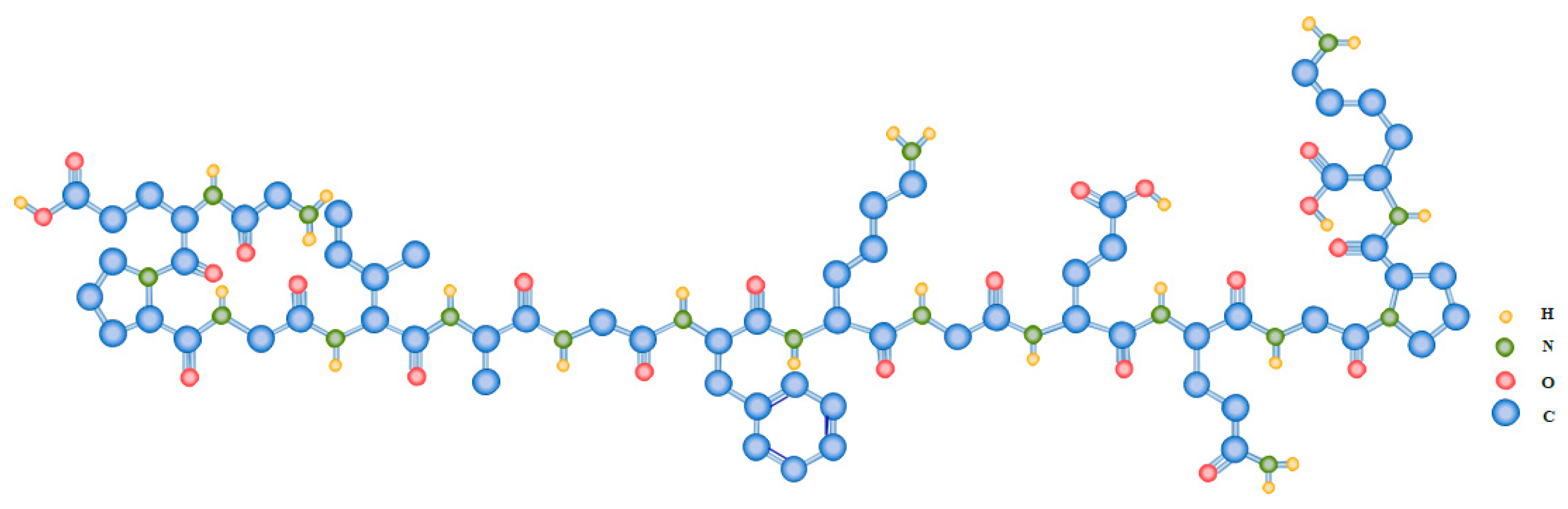
Figure 2. Exemplary amino acid repetition of the triplet (Gly-X-Y)n characteristic of type I collagen.
The most prevalent and thoroughly studied types of collagens are type I (almost present in all tissues and organs), type II (present in the cartilage, vitreous body, and nucleus pulposus), and type III (present in the skin, blood vessels, lungs, liver, and spleen) [45], which are used in tissue engineering and reconstructive medicine as well as in the pharmaceutical industry as compounds that extend the effects of drugs. Types I, II, and III collagens, especially type I, are also used as plastics in medicine and cosmetology. Type I collagen represents over 70% of the entire collagen family and makes up more than 90% of the collagen in the human body. It is mainly found in connective tissues such as body joints, cartilages, bones, sclerae, ligaments, tendons, intervertebral discs, corneas, adventitia of blood vessels, skin, and most hollow organs including gastrointestinal and genitourinary tracts [24,39,46][24][39][46]. In contrast, types II, III, and IV collagen are frequently seen. Type II collagen, for instance, is a structurally important part of the hyaline cartilage that lines the adult’s articular surfaces in addition to being present in other tissues including the intervertebral disc’s nucleus pulposus and the retina, sclera, and lens of the eye. Skin, lungs, intestinal walls, and blood vessel walls all contain type III collagen.
Type I collagen is composed of three polypeptide chains, two identical α1(I) chains and one α1(I) chain, each of which has roughly 1000 amino acid residues [47]. Hydroxylation of proline residues is a typical post-traditional modification of type I collagen that accounts for about 11–14% of amino acid residues and it is commonly used as a marker to detect and quantify collagen in tissues [48,49][48][49]. Whereas proline and hydroxyproline are essential for maintaining the triple helical structure under physiological conditions by forming hydrogen bonds that inhibit uncontrolled rotation, glycine is critical for packing the three helices [50,51][50][51].
The idea that the type I collagen molecule is made up of a single extended polypeptide chain with all amide bonds was brought forward by Astbury and Bell in 1940 [51]. In 1951, Pauling and Corey provided the correct structures for the α-helix and β-sheet [52]. In that proposal structure, three polypeptide chains were connected in a helical configuration by hydrogen bonds. These hydrogen bonds necessitated the production of two of the three peptide bonds and involved four of the six main chain heteroatoms inside each amino acid triplet [52]. The collagen triple helix structure was reconstructed in 1954 by Ramachandran and Kartha as a right-handed triple helix of three staggered, left-handed helices with one peptide bond and two hydrogen bonds within each triplet [53]. In 1955, this structure was improved by Rich and Crick, North, and Colleagues thanks to which the triple-helical structure that is still used today was unveiled. This structure has helical symmetry and just one crosslinking hydrogen bond per triplet [54]. Changes in the proposed structure of collagen from the beginning and its modification to the final structure accepted by the scientific community are shown in Figure 3.
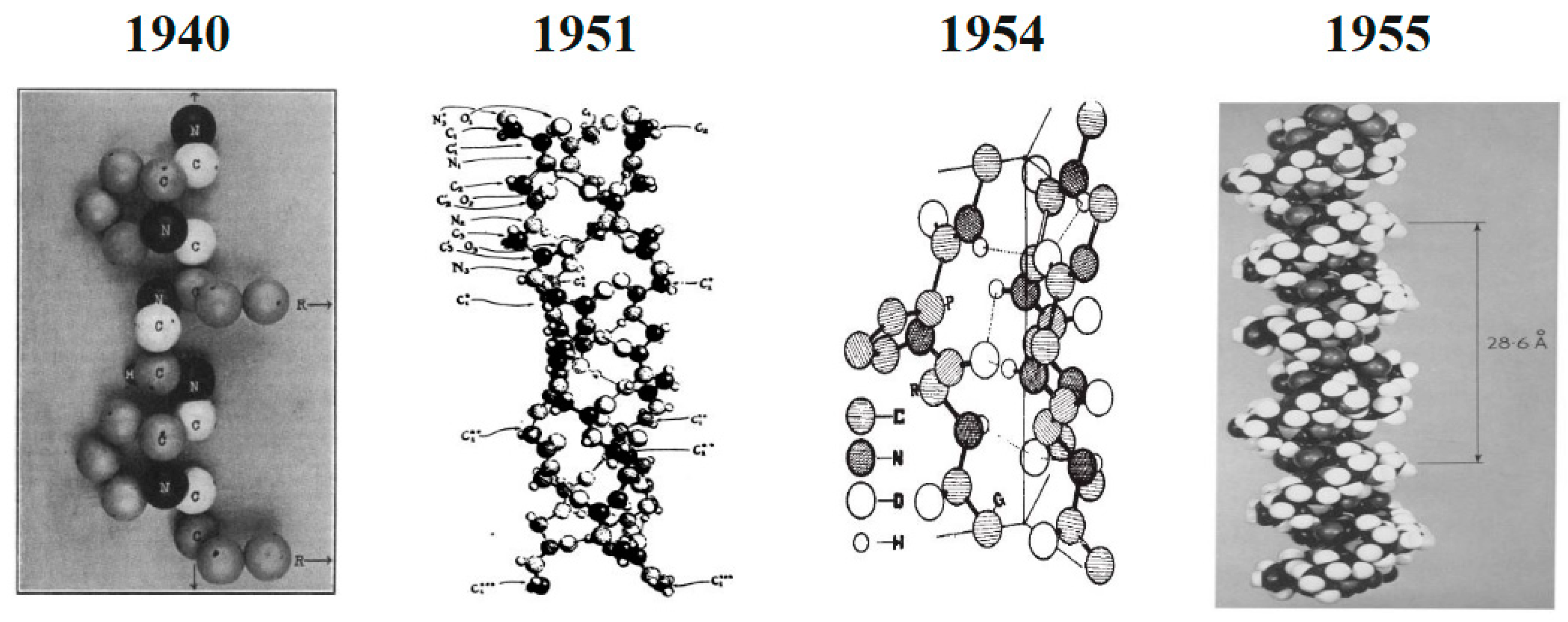
Figure 3. Changes in the proposed structure of type I collagen from the beginning and its modification to the final accepted structure. Adapted from [52]. Reproduced from [51] with permission from springer Nature, 1940. Reproduced from [53] with permission from springer Nature, 1954. Reproduced from [54] with permission from Elsevier, 1955.
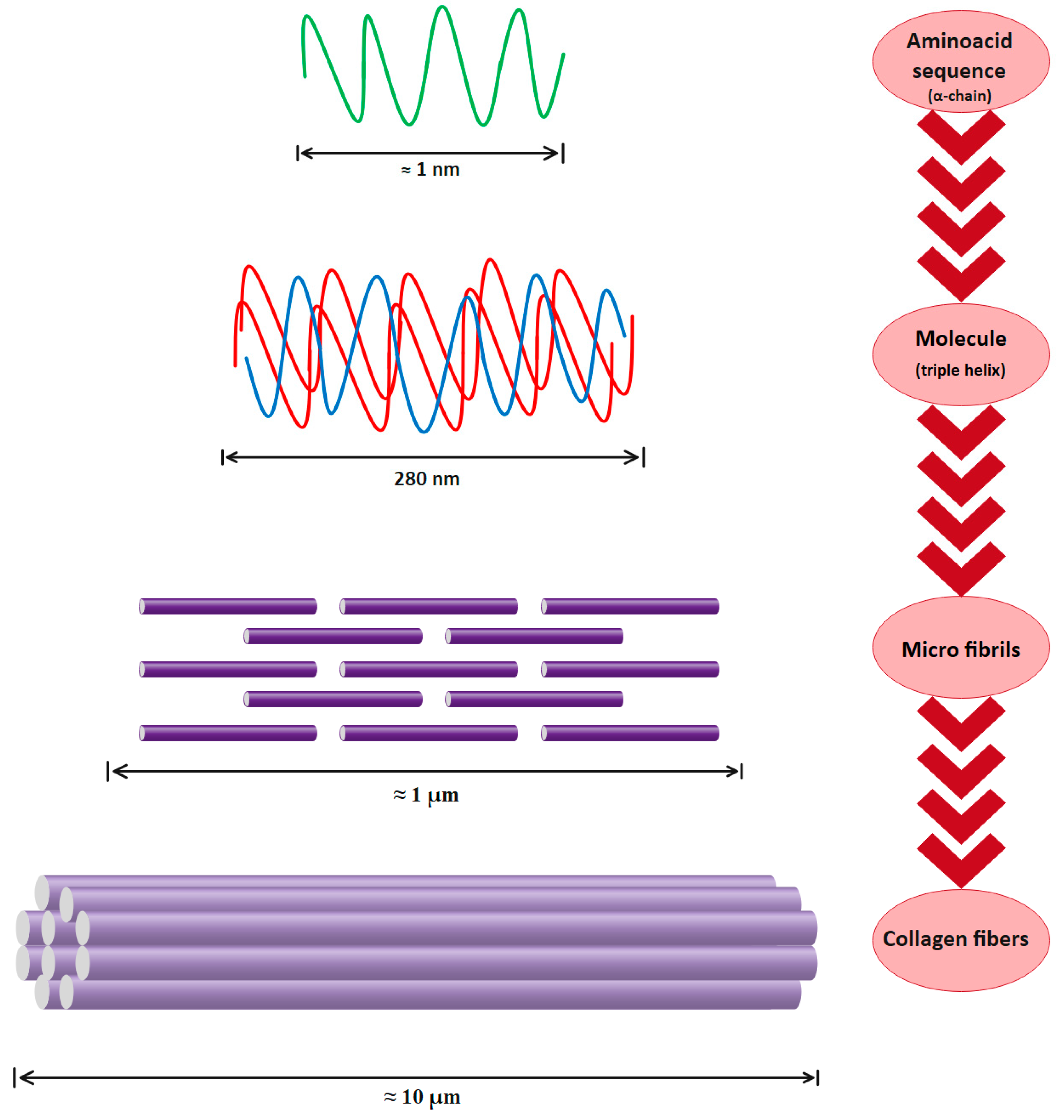

As is shown in Figure 4, three polypeptide α-chains form the trimeric molecule that represents the type I collagen unit (length ≈ 300 nm, diameter ≈ 1.5 nm). Three parallel, left-handed polyproline-II helices are arranged in a right-handed bundle [55,56][55][56]. Multiple collagen units are assembled into fibrils (length ≈ μm, diameter ≈ 100 nm) and then fibers (length ≈ mmm diameter ≈ 10 μm) with dimensions and orientation that are strictly tissue-dependent [28,57][28][57].
Figure 4. Type I collagen hierarchical organization.
Thus, type I collagen is a hierarchically organized protein. The primary structure of type I collagen consists of three α helices: two identical α1(I) and one α2(I) helices of approximately 1000 amino acids and a molecular weight of about 130–140 kDa and 110–120 kDa, respectively. The collagen molecule has a triple helical part and two non-helical parts at both ends (called telopeptides), with a molecular weight of 300–400 kDa, a length of 280 nm, and a width of 1.4 nm [58,59][58][59]. The secondary structure consists of each of these chains twisted in the form of a left-handed helix with three amino-acid repetitions in each turn. The tertiary structure, the inflexible structure, is created when the chains are then twisted three times around one another. Finally, in the quaternary structure, collagen molecules assemble into fibrils and then fibers. Because of the intermolecular and intramolecular interactions, this collagen organization is very stable [25,60][25][60]. Obviously, the collagen structure’s stability is directly dependent on its chemical composition. For instance, the triple helix of collagen grows stronger as the percentage of amino acids is higher, such as proline and hydroxyproline. The pyrrolidine rings are directly responsible for the polypeptide chain’s movement reduction [22,61][22][61]. Preservation of collagen’s structural integrity results in an improvement in physical properties, an increase in thermal stability, and a decrease in the denaturation temperature [62,63,64][62][63][64].
Theoretical examination of the mechanical characteristics of collagen at several levels, including the main monomer, individual collagen fibrils, and collagen fibers, is possible by studying collagen’s structured nature. Studying main monomers and fibrils made from type I collagen has likely provided the most direct measurements of the mechanical properties of collagen. Over the recent decades, researchers have used a variety of biophysical and theoretical methods, and recent developments in the Atomic Force Microscopy (AFM) approach have made it possible to perform more accurate evaluations [65]. According to estimates, the fracture strength of individual collagen triple helices is 11 GPa, which is much higher than that of collagen fibrils, which is 0.5 GPa [66]. This difference makes sense because, whereas the breaking of a fibril does not always entail the breakdown of covalent bonds, the breaking of individual collagen triple helices necessitates the unwinding of the triple helix and ultimately breaking of the covalent bonds [67]. In contrast to dehydrated type I collagen fibrils from mammalian sources, which have a Young’s modulus of about 5 GPa according to AFM tests, individual collagen triple helices monomers have a Young’s modulus between 6 and 7 GPa. Because collagen fibrils are anisotropic, another crucial measure of a collagen fibril’s strength is its shear modulus, which determines stiffness [68].
Furthermore, AFM indicated that the shear modulus of dehydrated fibrils of type I collagen from mammalian sources is between 30 and 35 MPa. These fibrils’ shear modulus was drastically decreased by hydration, but was increased by cross-linking. It is important to note that while some cross-linking is beneficial for the mechanical qualities of collagen fibrils, excessive cross-linking causes collagen fibrils to become highly brittle, which is a common sign of aging [69]. Investigation of the mechanical properties of collagen fibrils demonstrated that the length of the individual collagen triple helices monomer has been chosen by nature in a way to maximize the strength of the produced collagen fibril through effective energy dissipation. Simulations indicate that individual collagen triple helices monomers either longer or shorter than the length of a type I collagen triple helix, which is 300 nm, would form collagen fibrils with low mechanical properties [62]. The thermal and structural stability of the collagen triple helix is strongly influenced by the chemical composition of amino acid and its type, which is caused by the type of animal and the living conditions. Indeed, hydroxyproline stabilizes and strengthens the collagen structure [70]. In addition to preserving collagen’s structure and enhancing its mechanical properties, hydroxyproline also plays an important role in its thermal stability. The denaturation temperature and denaturation enthalpy of collagen increases due to the presence of the hydroxyl group in hydroxyproline and the bonding with the pyrrolidine ring. The quantity of hydrogen bonds formed between hydroxyproline and pyrrolidine significantly influences the increment in enthalpy of denaturation. Therefore, the triple helix does have greater thermal stability the more water molecules that there are surrounding it [25,71][25][71].
One of the most basic roles of collagen in the body is to provide connective tissues with stability, structure, and resistance to stresses [19,20][19][20]. Moreover, collagen has the ability to manage a wide range of nonstructural activities, including cell proliferation, migration, differentiation, and communication [60,72][60][72].
3. Fish Collagen
Collagen sources, types, pre-extraction conditions, and process methods are the main parameters that determine extracted product properties, including molecular weight of the peptide chain, amino acid composition, molecular structure, solubility, and functional activity. Although native type I collagen could be extracted from different mammalian sources, the main source of extraction is bovine due to availability and biocompatibility. [73]. There are other alternative sources for extracting type I collagen, among which pig, horse, sheep, and rat can be mentioned [74,75,76,77][74][75][76][77]. It is possible to obtain mammalian collagen from a wide range of tissues, notably skin, bones, tendons, lung tissue, and connective tissues. Due to some restrictions in terms of health, cultural, social, and religious issues that are implied by traditional sources, research has concentrated on the development of a new source of extraction. Various resources from the sea, including vertebrates as well as invertebrates, have been studied and considered as collagen extraction sources. In particular, several fish species (e.g., Rachycentron canadum, Esoxlucius, Spotless smooth hound, Sciaenops ocellatus, Sardinella fimbriata, Coryphaena hippurus, Alaska pollock, Takifugu flavidus, Pacu, Labeo rohita, Labeo catla, Tuna, Thunnus obesus, Scomber japonicus, Gadus morhua, Prionace glauca, Cichla ocellaris, Cyprinus carpio, Oreochromis niloticus, etc.) aquatic reptiles (such as the soft-shelled turtles), sponges, corals, octopuses, squids, starfish, jellyfish, cuttlefish and sea cucumbers, sea anemones, sea urchins, mussels, and shells were considered.
Skin, scales, bones, skull, swimming bladder, and remaining viscera, are by-products of fish that may be used as sources of collagen (Figure 5). Among all fish by-products, skin traditionally has been reported as the best source of fish collagen extraction [12,78,79,80,81][12][78][79][80][81].
Figure 5. By-products of fish as potential sources of collagen extraction.
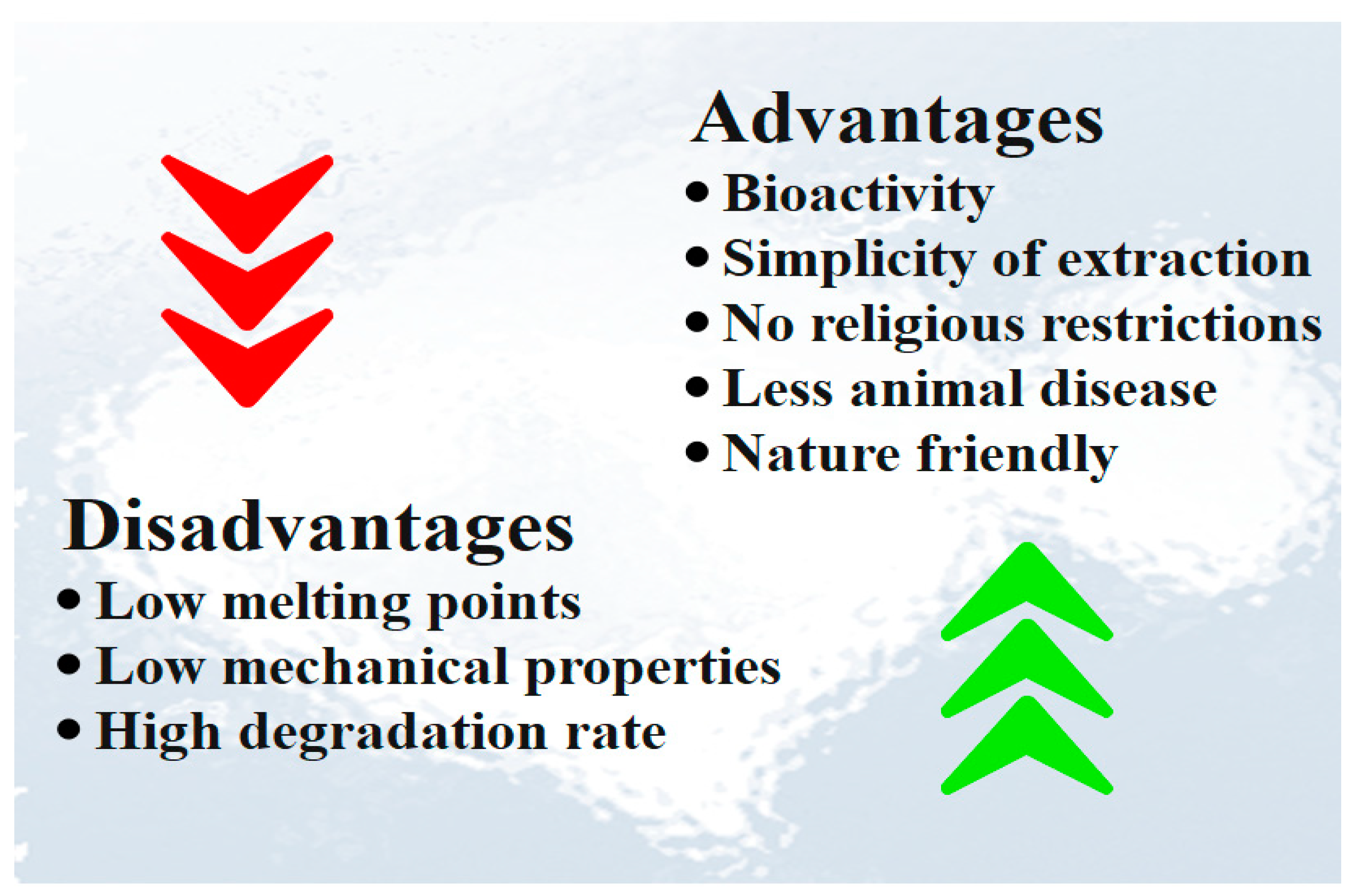
Fish collagen physicochemical properties were found to be similar to mammalian collagen, but with some advantages such as (1) capability of purification and extraction; (2) aquaculture and accessibility to fishing by-product; (3) lower risk of disease transmission compared to mammalian collagen due to high ontogenetic difference between fish and humans; (4) lack of religious and cultural limitation; (5) slightly different chemical composition; (6) low viscosity; (7) non toxicity; (8) reasonable homeostatic properties; (9) bio-resorbability; (10) more simple extraction method; (11) more adaptable and metabolic compatibility; and (12) minimal inflammatory response (Figure 6) [82,83,84,85][82][83][84][85]. Although fish collagen has several advantages, it suffers from several disadvantages such as low denaturation temperature, low mechanical properties, and high degradation rate [78]. The major drawback of fish collagen compared to mammalian collagen is the lower denaturation temperature, which limits its medical applications [70]. During denaturation, collagen turns into gelatin, where the hydrogen bonds that support the helical structure are partially or completely destroyed, and it loses its structural role and its conformation-related biological activity [42,43][42][43]. The second main drawback of fish-derived collagen is its low mechanical resistance which limits its applications. Many efforts have been made to improve its mechanical properties and degradation profiles, including chemical or enzymatic cross-linking [86,87][86][87]. The different advantages and disadvantages of fish collagen are shown in Figure 6.
Figure 6. Advantages and disadvantages of fish collagen.
References
- Pal, G.K.; Nidheesh, T.; Suresh, P.V. Comparative study on characteristics and in vitro fibril formation ability of acid and pepsin soluble collagen from the skin of catla (Catla catla) and rohu (Labeo rohita). Food Res. Int. 2015, 76, 804–812.
- Jia, Y.; Wang, H.H.; Wang, H.H.; Li, Y.; Wang, M.; Zhou, J. Biochemical properties of skin collagens isolated from black carp (Mylopharyngodon piceus). Food Sci. Biotechnol. 2012, 21, 1585–1592.
- Wang, L.; An, X.; Yang, F.; Xin, Z.; Zhao, L.; Hu, Q. Isolation and characterisation of collagens from the skin, scale and bone of deep-sea redfish (Sebastes mentella). Food Chem. 2008, 108, 616–623.
- Wang, S.; Zhang, Y. Study of writing problem in college general english course—Reflection on the reform of college english course. J. Lang Teach. Res. 2017, 8, 176–183.
- Nagai, T.; Izumi, M.; Ishii, M. Fish scale collagen. Preparation and partial characterization. Int. J. Food Sci. Technol. 2004, 39, 239–244.
- Silvipriya, K.S.; Krishna Kumar, K.; Bhat, A.R.; Dinesh Kumar, B.; John, A.; Lakshmanan, P. Collagen: Animal Sources and Biomedical Application. J. Appl. Pharm. Sci. 2015, 5, 123–127.
- Shaw, C.; Knopf, K.; Kloas, W. Fish Feeds in Aquaponics and Beyond: A Novel Concept to Evaluate Protein Sources in Diets for Circular Multitrophic Food Production Systems. Sustainability 2022, 14, 4064.
- de Melo Oliveira, V.; Assis, C.R.D.; de Aquino Marques Costa, B.; de Araújo Neri, R.C.; Monte, F.T.D.; da Costa Vasconcelos Freitas, H.M.S.; Franca, R.C.P.; Santos, J.F.; de Souza Bezerra, R.; Figueiredo Porto, A.L. Physical, biochemical, densitometric and spectroscopic techniques for characterization collagen from alternative sources: A review based on the sustainable valorization of aquatic by-products. J. Mol. Struct. 2021, 1224, 129023.
- Sorushanova, A.; Delgado, L.M.; Wu, Z.; Shologu, N.; Kshirsagar, A.; Raghunath, R.; Mullen, A.M.; Bayon, Y.; Pandit, A.; Raghunath, M.; et al. The Collagen Suprafamily: From Biosynthesis to Advanced Biomaterial Development. Adv. Mater. 2019, 31, 1801651.
- Shavandi, A.; Hou, Y.; Carne, A.; McConnell, M.; Bekhit, A.E.-D.A. Marine Waste Utilization as a Source of Functional and Health Compounds. Adv. Food Nutr. Res. 2019, 87, 187–254.
- Furtado, M.; Chen, L.; Chen, Z.; Chen, A.; Cui, W. Development of fish collagen in tissue regeneration and drug delivery. Eng. Regen. 2022, 3, 217–231.
- Maschmeyer, T.; Luque, R.; Selva, M. Upgrading of marine (fish and crustaceans) biowaste for high added-value molecules and bio(nano)-materials. Chem. Soc. Rev. 2020, 49, 4527–4563.
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The Future of Aquatic Protein: Implications for Protein Sources in Aquaculture Diets. One Earth 2019, 1, 316–329.
- Mishra, P.K.; Gautam, R.K.; Kumar, V.; Kakatkar, A.S.; Chatterjee, S. Synthesis of Biodegradable Films Using Gamma Irradiation from Fish Waste. Waste Biomass Valorization 2021, 12, 2247–2257.
- Xu, C.; Nasrollahzadeh, M.; Selva, M.; Issaabadi, Z.; Luque, R. Waste-to-wealth: Biowaste valorization into valuable bio(nano)materials. Chem. Soc. Rev. 2019, 48, 4791–4822.
- Al Khawli, F.; Pateiro, M.; Domínguez, R.; Lorenzo, J.M.; Gullón, P.; Kousoulaki, K.; Ferrer, E.; Berrada, H.; Barba, F.J. Innovative Green Technologies of Intensification for Valorization of Seafood and Their By-Products. Mar. Drugs 2019, 17, 689.
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery Wastes as a Yet Undiscovered Treasure from the Sea: Biomolecules Sources, Extraction Methods and Valorization. Mar. Drugs 2020, 18, 622.
- Nimni, M.E.; Harkness, R.D. Molecular Structure and Functions of Collagen. In Collagen; CRC Press: Boca Raton, FL, USA, 2018; Volume 1, pp. 1–78.
- Coppola, D.; Lauritano, C.; Esposito, F.P.; Riccio, G.; Rizzo, C.; De Pascale, D. Fish Waste: From Problem to Valuable Resource. Mar. Drugs 2021, 19, 116.
- Mahmood, A.; Patel, D.; Hickson, B.; DesRochers, J.; Hu, X. Recent progress in biopolymer-based hydrogel materials for biomedical applications. Int. J. Mol. Sci. 2022, 23, 1415.
- Castile, J.D.; Taylor, K.M.G.; Buckton, G. The influence of incubation temperature and surfactant concentration on the interaction between dimyristoylphosphatidylcholine liposomes and poloxamer surfactants. Int. J. Pharm. 2001, 221, 197–209.
- El Blidi, O.; Omari, N.E.l.; Balahbib, A.; Ghchime, R.; El Menyiy, N.; Ibrahimi, A.; Kaddour, K.B.; Bouyahya, A.B.; Chokairi, O.; Barkiyou, M. Extraction Methods, Characterization and Biomedical Applications of Collagen: A Review. Biointerface Res. Appl. Chem. 2021, 11, 13587–13613.
- Sbricoli, L.; Guazzo, R.; Annunziata, M.; Gobbato, L.; Bressan, E.; Nastri, L. Selection of collagen membranes for bone regeneration: A literature review. Materials 2020, 13, 786.
- Xu, Q.; Torres, J.E.; Hakim, M.; Babiak, P.M.; Pal, P.; Battistoni, C.M.; Nguyen, M.; Panitch, A.; Solorio, L.; Liu, J.C. Collagen- and hyaluronic acid-based hydrogels and their biomedical applications. Mater. Sci. Eng. R Rep. 2021, 146, 100641.
- Naomi, R.; Ridzuan, P.M.; Bahari, H.; Vallejo-Giraldo, C. Current Insights into Collagen Type I. Polymers 2021, 13, 2642.
- Coppola, D.; Oliviero, M.; Vitale, G.A.; Lauritano, C.; D’Ambra, I.; Iannace, S.; de Pascale, D. Marine Collagen from Alternative and Sustainable Sources: Extraction, Processing and Applications. Mar. Drugs 2020, 18, 214.
- Gallo, N.; Natali, M.L.; Sannino, A.; Salvatore, L. An Overview of the Use of Equine Collagen as Emerging Material for Biomedical Applications. J. Funct. Biomater. 2020, 11, 79.
- Jafari, H.; Lista, A.; Siekapen, M.M.; Ghaffari-Bohlouli, P.; Nie, L.; Alimoradi, H.; Shavandi, A. Fish Collagen: Extraction, Characterization, and Applications for Biomaterials Engineering. Polymers 2020, 12, 2230.
- Ikoma, T.; Kobayashi, H.; Tanaka, J.; Walsh, D.; Mann, S. Physical properties of type I collagen extracted from fish scales of Pagrus major and Oreochromis niloticas. Int. J. Biol. Macromol. 2003, 32, 199–204.
- Nomura, Y.; Sakai, H.; Ishii, Y.; Shirai, K. Preparation and some properties of type I collagen from fish scales. Biosci. Biotechnol. Biochem. 1996, 60, 20922094.
- Nagai, T.; Suzuki, N. Isolation of collagen from fish waste material—skin, bone and fins. Food Chem. 2000, 68, 277–281.
- Rauta, P.R.; Mohanta, Y.K.; Nayak, D. Nanotechnology in Biology and Medicine: Research Advancements & Future Perspectives; CRC Press: Boca Raton, FL, USA, 2019.
- Lo, S.; Fauzi, M.B. Current Update of Collagen Nanomaterials—Fabrication, Characterisation and Its Applications: A Review. Pharmaceutics 2021, 13, 316.
- Naskar, A.; Kim, K.S. Recent Advances in Nanomaterial-Based Wound-Healing Therapeutics. Pharmaceutics 2020, 12, 499.
- Ucar, B. Natural biomaterials in brain repair: A focus on collagen. Neurochem. Int. 2021, 146, 105033.
- Dong, C.; Lv, Y. Application of Collagen Scaffold in Tissue Engineering: Recent Advances and New Perspectives. Polymers 2016, 8, 42.
- Arun, A.; Malrautu, P.; Laha, A.; Luo, H.; Ramakrishna, S. Collagen Nanoparticles in Drug Delivery Systems and Tissue Engineering. Appl. Sci. 2021, 11, 11369.
- Neff, L.S.; Bradshaw, A.D. Cross your heart? Collagen cross-links in cardiac health and disease. Cell Signal. 2021, 79, 109889.
- Gelse, K.; Pöschl, E. Reviews TA-A drug delivery, 2003 U. Collagens—Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546.
- Schweitzer, M.H.; Suo, Z.; Avci, R.; Asara, J.M.; Allen, M.A.; Arce, F.T.; Horner, J.R. Analyses of Soft Tissue from Tyrannosaurus rex Suggest the Presence of Protein. Science 2007, 316, 277–280.
- Torres, J.M.; Borja, C.; Gibert, L.; Ribot, F.; Olivares, E.G. Twentieth-Century Paleoproteomics: Lessons from Venta Micena Fossils. Biology 2022, 11, 1184.
- Bächinger, H.P.; Mizuno, K.; Vranka, J.A.; Boudko, S.P. Collagen Formation and Structure. Compr. Nat. Prod II Chem. Biol. 2010, 5, 469–530.
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978.
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R.P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958.
- Inoue, Y.; Itoh, H.; Aoki, M.; Ogawa, S.; Yamane, T.; Baba, T.; Tachibana, N.; Kohno, M.; Oishi, Y.; Kobayashi-Hattori, K. Accelerating effect of soy peptides containing collagen peptides on type I and III collagen levels in rat skin. Biosci. Biotechnol. Biochem. 2012, 76, 1549–1551.
- Minor, A.J.; Coulombe, K.L.K. Engineering a collagen matrix for cell-instructive regenerative angiogenesis. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 2407–2416.
- Bornstein, P.; Sage, H. Structurally distinct collagen types. Annu. Rev. Biochem. 1980, 49, 957–1003.
- Salvatore, L.; Gallo, N.; Aiello, D.; Lunetti, P.; Barca, A.; Blasi, L.; Madaghiele, M.; Bettini, S.; Giancane, G.; Hasna, M.; et al. An insight on type I collagen from horse tendon for the manufacture of implantable devices. Int. J. Biol. Macromol/ 2020, 154, 291–306.
- Ignat’eva, N.Y.; Danilov, N.A.; Averkiev, S.V.; Obrezkova, M.V.; Lunin, V.V.; Sobol’, E.N. Determination of hydroxyproline in tissues and the evaluation of the collagen content of the tissues. J. Anal. Chem. 2007, 62, 51–57.
- Privalov, P.L.; Tiktopulo, E.I.; Tischenko, V.M. Stability and mobility of the collagen structure. J. Mol. Biol. 1979, 127, 203–216.
- Astbury, W.T.; Bell, F.O. Molecular Structure of the Collagen Fibres. Nature. 1940, 145, 421–422.
- Pauling, L.; Corey, R.B. The structure of fibrous proteins of the collagen-gelatin group. Proc. Natl. Acad. Sci. USA 1951, 37, 272–281.
- Ramachandran, G.N.; Kartha, G. Structure of Collagen. Nature 1954, 174, 269–270.
- Rich, A.; Crick, F.H.C. The molecular structure of collagen. J. Mol. Biol. 1961, 3, 483–506.
- Cowan, P.M.; Mcgavin, S.; North, A.C. The Polypeptide Chain Configuration of Collagen. Nature 1955, 176, 1062–1064.
- Bella, J. Collagen structure: New tricks from a very old dog. Biochem. J. 2016, 473, 1001–1025.
- Lim, Y.S.; Ok, Y.J.; Hwang, S.Y.; Kwak, J.Y.; Yoon, S. Marine collagen as a promising biomaterial for biomedical applications. Mar. Drugs 2019, 17, 467.
- Chen, L.; Cheng, G.; Meng, S.; Ding, Y. Collagen Membrane Derived from Fish Scales for Application in Bone Tissue Engineering. Polymers 2022, 14, 2532.
- León-López, A.; Morales-Peñaloza, A.; Manuel Martínez-Juárez, V.; Vargas-Torres, A.; Zeugolis, D.I.; Aguirre-Álvarez, G. Hydrolyzed Collagen—Sources and Applications. Molecules 2019, 24, 4031.
- Liu, X.; Gao, Y.; Long, X.; Hayashi, T.; Mizuno, K.; Hattori, S.; Fujisaki, H.; Ogura, T.; Wang, D.O.; Ikejima, T. Type i collagen promotes the migration and myogenic differentiation of C2C12 myoblasts: Via the release of interleukin-6 mediated by FAK/NF-κB p65 activation. Food Funct. 2020, 11, 328–338.
- Song, Z.; Liu, H.; Chen, L.L.; Chen, L.L.; Zhou, C.; Hong, P.; Deng, C. Characterization and comparison of collagen extracted from the skin of the Nile tilapia by fermentation and chemical pretreatment. Food Chem. 2021, 340, 128139.
- Peng, X.; Cui, Y.; Chen, J.; Gao, C.; Yang, Y.; Yu, W.; Rai, K.; Zhang, M.; Nian, R.; Bao, Z.; et al. High-Strength Collagen-Based Composite Films Regulated by Water-Soluble Recombinant Spider Silk Proteins and Water Annealing. ACS Biomater. Sci. Eng. 2022, 8, 3341–3353.
- Bao, Z.; Sun, Y.; Rai, K.; Peng, X.; Wang, S.; Nian, R.; Xian, M. The promising indicators of the thermal and mechanical properties of collagen from bass and tilapia: Synergistic effects of hydroxyproline and cysteine. Biomater. Sci 2018, 6, 3042–3052.
- Bao, Z.; Gao, M.; Fan, X.; Cui, Y.; Yang, J.; Peng, X.; Xian, M.; Sun, Y.; Nian, R. Development and characterization of a photo-cross-linked functionalized type-I collagen (Oreochromis niloticus) and polyethylene glycol diacrylate hydrogel. Int. J. Biol. Macromol. 2020, 155, 163–173.
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958.
- Buehler, M.J. Nature designs tough collagen: Explaining the nanostructure of collagen fibrils. Proc. Natl. Acad. Sci. USA 2006, 103, 12285–12290.
- Shen, Y.; Levin, A.; Kamada, A.; Toprakcioglu, Z.; Rodriguez-Garcia, M.; Xu, Y.; Knowles, T.P. From Protein Building Blocks to Functional Materials. ACS Nano 2021, 15, 5819–5837.
- Lin, J.; Shi, Y.; Men, Y.; Wang, X.; Ye, J.; Zhang, C. Mechanical roles in formation of oriented collagen fibers. Tissue Eng.-Part B Rev. 2020, 26, 116–128.
- Yang, L.; Van Der Werf, K.O.; Fitié, C.F.C.; Bennink, M.L.; Dijkstra, P.J.; Feijen, J. Mechanical properties of native and cross-linked type I collagen fibrils. Biophys. J. 2008, 94, 2204–2211.
- Tihăuan, B.-M.; Pircalabioru, G.G.; Axinie Bucos, M.; Marinaș, I.C.; Nicoară, A.-C.; Măruțescu, L.; Oprea, O.; Matei, E.; Maier, S.S. Crosslinked Collagenic Scaffold Behavior Evaluation by Physico-Chemical, Mechanical and Biological Assessments in an In Vitro Microenvironment. Polymers 2022, 14, 2430.
- Hossain, M.S.; Ebrahimi, H.; Ghosh, R. Fish scale inspired structures—A review of materials, manufacturing and models. Bioinspir. Biomim. 2022, 17, 061001.
- Sharma, S.; Dwivedi, S.; Chandra, S.; Srivastava, A.; Vijay, P. Collagen: A Brief Analysis. Oral. Maxillofac. Pathol. J. 2019, 10, 11–17.
- Salvatore, L.; Gallo, N.; Natali, M.L.; Campa, L.; Lunetti, P.; Madaghiele, M.; Blasi, F.S.; Corallo, A.; Capobianco, L.; Sannino, A. Marine collagen and its derivatives: Versatile and sustainable bio-resources for healthcare. Mater. Sci. Eng. C 2020, 113, 110963.
- Ferraro, V.; Gaillard-Martinie, B.; Sayd, T.; Chambon, C.; Anton, M.; Santé-Lhoutellier, V. Collagen type I from bovine bone. Effect of animal age, bone anatomy and drying methodology on extraction yield, self-assembly, thermal behaviour and. Int. J. Biol. Macromol. 2017, 97, 55–66.
- Davison-Kotler, E.; Marshall, W.S.; García-Gareta, E. Sources of Collagen for Biomaterials in Skin Wound Healing. Bioengineering 2019, 6, 56.
- Paschou, A.M.; Katsikini, M.; Christofilos, D.; Arvanitidis, J.; Ves, S.; Katsikini, C.M. High pressure Raman study of type-I collagen. Wiley Online Libr. 2018, 285, 2641–2653.
- Bhuimbar, M.V.; Bhagwat, P.K.; Dandge, P.B. Extraction and characterization of acid soluble collagen from fish waste: Development of collagen-chitosan blend as food packaging film. J. Environ. Chem. Eng. 2019, 7, 102983.
- Subhan, F.; Hussain, Z.; Tauseef, I.; Shehzad, A.; Wahid, F. A review on recent advances and applications of fish collagen. Crit. Rev. Food Sci. Nutr. 2021, 61, 1027–1037.
- Heidari, M.G.; Rezaei, M. Extracted pepsin of trout waste and ultrasound-promoted method for green recovery of fish collagen. Sustain. Chem. Pharm. 2022, 30, 100854.
- Morikawa, T.; Reátegui-Pinedo, N.; Salirrosas, D.; Sánchez-Tuesta, L.; Quiñones, C.; Jáuregui-Rosas, S.R.; Barraza, G.; Cabrera, A.; Ayala-Jara, C.; Miliani Martinez, R.; et al. Characterization of Collagen from Three Genetic Lines (Gray, Red and F1) of Oreochromis niloticus (Tilapia) Skin in Young and Old Adults. Molecules 2022, 27, 1123.
- Felician, F.; Xia, C.; Qi, W. Collagen from marine biological sources and medical applications. Wiley Online Libr. 2018, 15, e1700557.
- Nirmal, N.P.; Santivarangkna, C.; Rajput, M.S.; Benjakul, S.; Maqsood, S. Valorization of fish byproducts: Sources to end-product applications of bioactive protein hydrolysate. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1803–1842.
- Fassini, D.; Wilkie, I.C.; Pozzolini, M.; Ferrario, C.; Sugni, M.; Rocha, M.S.; Giovine, M.; Bonasoro, F.; Silva, T.H.; Reis, R.L. Diverse and productive source of biopolymer inspiration: Marine collagens. Biomacromolecules 2021, 22, 1815–1834.
- Sanchez, A.; Blanco, M.; Correa, B.; Perez-Martin, R.I.; Sotelo, C.G. Effect of fish collagen hydrolysates on type I collagen mRNA levels of human dermal fibroblast culture. Mar. Drugs 2018, 16, 144.
- Lu, W.-C.; Chiu, C.-S.; Chan, Y.-J.; Guo, T.-P.; Lin, C.-C.; Wang, P.-C.; Po-Yu Lin, P.-Y.; Mulio, A.T.; Li, P.-H. An In Vivo Study to Evaluate the Efficacy of Blue Shark (Prionace glauca) Cartilage Collagen as a Cosmetic. Mar. Drugs 2022, 20, 633.
- Ahn, H.; Gong, D.J.; Lee, H.H.; Seo, J.Y.; Song, K.M.; Eom, S.J.; Yeo, S.Y. Mechanical Properties of Porcine and Fish Skin-Based Collagen and Conjugated Collagen Fibers. Polymers 2021, 13, 2151.
- Liu, S.; Lau, C.S.; Liang, K.; Wen, F.; Teoh, S.H. Marine collagen scaffolds in tissue engineering. Curr. Opin. Biotechnol. 2022, 74, 92–103.
More