Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Pedro Barbosa and Version 3 by Peter Tang.
Glucose, fructose, and galactose are widely used in the food industry as sweeteners and food additives. The over-consumption of these carbohydrates has been identified as a possible trigger of non-communicable diseases. These include insulin resistance, obesity, and type 2 diabetes. These sugars induce an energy overload with consequent adipose tissue (AT) expansion, contributing to the development of obesity and inflammation.
- insulin resistance
- sugar metabolism
- adipose tissue
1. Introduction
The Western diet is characterized by the presence of highly processed foods, which are particularly rich in salt, saturated fats, poor-quality protein, and simple carbohydrates, such as those deriving from corn, refined cereals, and sugars (glucose, fructose, and sucrose) [1]. Many of these carbohydrates present high glycemic and high insulinemic indices that quickly induce glucose and insulin stimulation peaks for short periods of time [2]. In contrast, diets with large contents of high-quality protein and plant-based foods, such as vegetables, nuts, fruits, and honey, which contain carbohydrates with low glycemic and low insulinemic indices, are generally considered healthier [1]. The energy overload caused by the increased consumption of refined sugars (free sugar) and saturated fats can lead to a drastic expansion of adipose tissue (AT) depots, especially when associated with a lack of physical activity [3]. In addition, a reduction in the fiber content of ingested foods may support metabolic destabilization, since sugars are quickly and freely available in the system [4][5]. Therefore, the Western diet has been identified as a possible trigger of the development of non-communicable diseases from an early stage of life. Insulin resistance (IR), obesity, type 2 diabetes mellitus (T2DM), and metabolic syndrome are some of the most prevalent non-communicable diseases worldwide [1][6][7].
2. Dietary (Free) Sugars
Free sugars are mono- and disaccharides that are added to food, excluding the naturally present types, such as lactose in milk and sucrose in fruits and vegetables, although these naturally present sugars can be considered as free sugars when they are added to a product [3][8]. Generally, free sugars are widely used in the food industry as sweeteners and food additives, especially in beverages and during food transformation and preparation [3][8], appearing in large proportions in the Western diet [9]. The monosaccharides, glucose, and fructose, as well as disaccharides, sucrose, and lactose, are used as additives in the Western diet [3][10], with fructose and glucose representing almost 50% of the added sugars [11]. These two monosaccharides are commonly used individually or in the form of sucrose or high-fructose corn syrup (HFCS). Sucrose is extracted and purified from sugar cane and sugar beet, while fructose is obtained by the enzymatic degradation of cornstarch into glucose or glucose polymers to be further isomerized enzymatically into fructose, producing the HFCS [3]. The main difference between sucrose and HFCS lies in the fructose content, which varies between 42% and 55% in HFCS and is 50% in sucrose. Moreover, sucrose is composed of glucose covalently bonded to fructose, while in HFCS, these molecules remain in their free forms, rendering them highly bioavailable and increasing their absorption when consumed [3][12]. Lactose is also widely used not only in the food industry, where it is applied in the form of condensed milk and in caramel flavors, but also in the pharmaceutical industry as a drug carrier [13]. In similarity to sucrose, lactose is a disaccharide of glucose and galactose that is used frequently in the confectionery and bakery industries. The production of caramel flavors using lactose depends on the Maillard reaction [13]. In the bakery industry, this process is important for creating the brown color of products, since lactose is not degraded by yeast [13].3. Sugar Metabolism and Its Effect on Adipose Tissue
In different proportions, all the sugars described above are widely used in the Western diet, and there is no consensus regarding their potential harmful effects on metabolism [11][14]. Sucrose and HFCS are composed of monomers of fructose and glucose, and despite their similar structures, they follow different metabolic pathways, as described below [15], while galactose, resulting from lactose, follows the Leloir metabolic pathway [16] (Figure 1).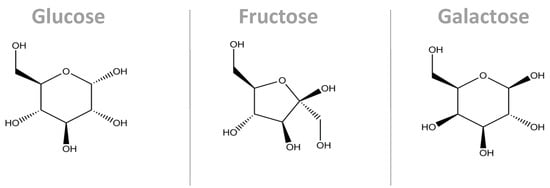
Figure 1. Molecular structure of the main monosaccharides that compose our diet. Despite their similarity in terms of the molecular structures, their conformations are different, causing them to interact with their microenvironments in different ways. Additionally, their metabolic pathways are different. These three molecules are the carbon skeleton for sucrose (glucose + fructose bonded by a α1,β2-glycosidic bond) and for lactose (glucose + galactose bonded by a β1,4-glycosidic bond) [17].
3.1. Sucrose and HFCS—Fructose and Glucose Metabolism
After ingestion, fructose is passively absorbed by the intestinal apical membrane via the high-affinity glucose transporter (GLUT)5, passed on to the portal circulation by GLUT2. On the other hand, glucose is absorbed by enterocytes, mainly through the co-transporters sodium-glucose linked transporter 1 (SGLT1) and GLUT2. Once in circulation, fructose is metabolized in the tissues by phosphofructokinase and glucokinase, and the latter is also regulated by insulin [9][15][18]. An increase in fructose consumption, associated with an unrestricted pathway, leads to a rapid increase in uric acid synthesis, gluconeogenesis, glycolysis, and de novo lipogenesis [15][19]. Unlike glucose, fructose is not regulated by the activity of phosphofructokinase, an enzyme that limits the glycolytic flux [19]. The hexo-phosphate and trios-phosphate intermediaries, resulting from fructolysis, are used as substrate for the pathways described above [15][19]. Moreover, excessive amounts of fructose will overload the liver’s capacity to oxidize it. Therefore, the fructose remains in circulation and can be used by other tissues, such as AT [20]. Fructose has been described as a potential lipogenic and adipogenic nutrient accelerating lipid deposition, particularly in visceral AT, and ectopic fat deposition in the other insulin-sensitive tissues, such as the liver and muscle [19][20][21]. In addition, fructose has been observed to disrupt insulin sensitivity in adults [21]. Stanhope and colleagues conducted a clinical study with 32 participants aged from 40 to 72 years. The subjects were divided into two groups, including glucose- and fructose-sweetened beverage consumers, for a period of 10 weeks. The study indicated an increase in the plasma lipids and lipoproteins in the fructose consumer group, while the glucose consumer group remained unchanged, except for the triglycerides, which showed an opposite pattern. Furthermore, the authors observed alterations in insulin sensitivity and glucose tolerance after 9 weeks of beverage consumption. The fructose consumer group showed increased insulin and glucose levels during an oral glucose tolerance test compared to baseline, while the glucose consumer group remained unchanged. Moreover, the insulin sensitivity index decreased by approximately 17% in the fructose consumption group [21]. Additionally, the same study indicated that the group who consumed fructose-sweetened beverages showed a higher expression of lipogenic genes in VAT [21]. Adipocytes lack the keto-hexokinase enzyme that converts fructose into fructose-1-phosphate. In these cells, fructose is converted to fructose-6-phosphate by hexokinase stimulating the conversion of pyruvate into acetyl-CoA, thus increasing the synthesis of fatty acids, and leading to consequent palmitate release. In this case, fructose is mostly used in anabolic pathways, in contrast to glucose [22]. Interestingly, Varma and collaborators (2015) postulated that fructose can trigger the oxidation of glucose to lactate in a dose-dependent manner while reducing the utilization of glucose in the glutamate and fatty acid synthesis pathways, using a 10% 13C2-D-glucose trace in cultured adipocytes [23]. Furthermore, the study also indicated a reduction in glucose conversion to glycogen. Consequently, glucose is driven to the one-carbon cycle and the glycine cleavage pathway (SOG pathway) through 3-phosphoglycerate to synthesize serine and other intermediates that are important for the generation of NADPH and ATP [23][24]. A treatment with 10% fructose solution in the drinking water of rats for 24 weeks induced the upregulation of genes related to the insulin-signaling pathway, particularly phosphoinositol-3-kinase (PI3K), protein kinase-B (AKT), insulin receptor(IR)-β, insulin receptor substrate (IRS)-1, and the mammalian target of rapamycin (mTOR), but also those related to adipocyte homeostasis, such as peroxisome proliferator-activated receptor (PPAR)γ and nuclear factor erythroid-2-related factor 2 (NrF2) [25]. Although, the authors did not find any correlation between insulin impairment and adipose tissue inflammation [25]. On the other hand, different studies have suggested that fructose-rich diets affect insulin action and AT metabolism, inducing changes in the secretory patterns of resistin, adiponectin, leptin, and specific adipokines, which, in turn, are linked to inflammation and insulin resistance [26][27]. Furthermore, it has been shown that fructose induces an increase in leptin secretion and, consequently, leptin resistance [28]. However, the mechanism determining how leptin resistance is established is not well understood. Despite its different functions in the organism, leptin has a significant impact on inflammation and the inflammatory onset, not only locally at the tissue level but also systemically, perpetuating further inflammation [25][28]. In addition, Marek et al. (2015) provided important insights into the effects of fructose on the adipocyte endoplasmic reticulum (RE) redox status in mice. Moreover, increased levels of monocyte chemoattractant protein (MCP)-1, intercellular adhesion molecule (ICAM)-1, and tumor necrosis factor (TNF)-α expression by AT have already been described in response to fructose metabolism. The expression of these genes leads to an increase in macrophage infiltration on AT [28][29] but also the release of other pro-inflammatory cytokines by the adipocytes [28]. Interestingly, this inflammatory process caused by the excessive consumption of fructose-rich foods seems to have a gender-dependent impact, particularly regarding the expression of inflammatory markers in VAT [25][30]. Considering the above, Kovačević and collaborators tested the impacts of the ingestion of 10% (w/v) fructose solution on female and male Wistar rats for 9 weeks and noticed that, despite the diet used, there was no impact on the glucose or insulin levels. However, the fructose-treated females showed a significant reduction in the Akt and pAkt-Ser473 levels in VAT and an increase in the PTP1B protein levels in comparison to the standard chow, while the male rats only showed a decreased pAkt-Ser473/Akt ratio. Furthermore, the fructose-treated female showed increased levels of nuclear factor (NF)κB in VAT, followed by increased levels of TNF-α, interleukin (IL)-6, and IL-1β mRNA, as well as increased levels of F4/80, a macrophage marker. In contrast, the male fructose-treated rats showed no differences in the NFκB expression levels [30]. Data suggest that fructose-rich diets induce chronic inflammation in a dose-dependent manner [31]. Wang et al. (2020) fed six-week-old Sprague Dawley male rats with low (2.6 g/kg/day), medium (5.3 g/kg/day), and high doses (10.5 g/kg/day) of fructose for 20 weeks and identified a dose-dependent increase in the circulating levels of IL-6, TNF-α, and macrophage inflammatory protein (MIP)-2 when compared to the controls, while the opposite was observed for IL-10 in an inverse pattern. In the same study, the highest fructose dose led to an increase in the number of inflammatory cells in the pancreas, a 10% increase in liver steatosis, colon inflammation, and gut microbiota alterations. Furthermore, the acute inflammatory response to a fructose-rich diet during the postprandial state was recently studied in healthy subjects and in patients with T2DM. The data showed that the levels of IL-6 and ICAM-1 were increased in the healthy subjects in the postprandial state, while MCP-1 was increased in both the healthy subjects and in the patients with T2DM [18]. The glycemic load caused by the consumption of sucrose and HFCS has also been suggested to be a possible trigger of the inflammatory processes [32][33][34]. In addition, a recent study conducted by Patkar et al. (2021) indicated that the long-term (3 months) consumption of 5% (w/v) sucrose could be a trigger of the onset of systemic low-grade inflammation, without the induction of obesity, in male Wistar rats. Furthermore, they observed an increase in some of the immune cell populations in circulation, such as lymphocytes, basophils, and neutrophils [11].3.2. Lactose—Galactose and Glucose
Lactose is a disaccharide composed of galactose and glucose, and it is metabolized in the intestinal lumen by lactase. A complementary mechanism of lactose metabolism is through the colonic microbiota, primarily in adults [35]. This sugar differs from the other mono- and disaccharides since it has no specific transporter to pass through the intestinal barrier. However, concentrations of around 0.02 mmol/L of lactose have already been found in circulation in healthy young adult men, contrary to what had been postulated [35]. Despite not being metabolized in other tissues, lactose seems to play a role in systemic inflammation, a topic that will be discussed further in this resviearchw. On the other hand, lactose monomers are easily metabolized by the organism. Like fructose, galactose is absorbed by the endothelial cells, released into the blood stream, and transported to the liver via the portal vein [36]. A large amount of galactose is retained and metabolized in the liver, but small amounts remain in circulation and reach other tissues, such as AT and the skeletal muscle [36]. It follows the Leloir pathway once it enters the adipocytes [16]. Krycer et al. (2020) treated 3T3-L1 adipocytes with either 25 mM of glucose or 25 mM of galactose, and they found a reduction in lactate production by the galactose-treated cells, even after insulin stimulation [37]. On the other hand, mitochondrial respiration was increased upon treatment with galactose and upon insulin stimulation [37]. Thus, galactose appears to be used to feed a different pathway, rather than glycolysis, in the adipocytes. To test this, the authors used tracers to differentiate the galactose and glucose carbons using 13C-labels (both 25 mM) [37]. They found a reduction in glucose-6-phosphate after the galactose treatment [37]. The glucose-6-phosphate is considered a common point between the glucose and galactose oxidation pathways and how galactose enters the glycolytic pathway. The data indicate that galactose follows a different pathway from glycolysis and is a poor substrate for energy metabolism [16]. Interestingly, similar results were found in a study on mature adipocytes isolated from rats [37]. However, Krishna et al. (2020) did not identify any effect of 25 mM of galactose or 25 mM of lactose treatment on adipocyte differentiation [38]. Considering the low degradation rate of galactose compared to glucose, high amounts of galactose in circulation can lead to galactosemia and, consequently, to the glycation of different macromolecules, including amino acids, creating advanced glycation end products (AGEs) and reactive oxygen species (ROS) [5][39]. These molecules are responsible for tissue damage that, in turn, leads to accelerated aging [40][41]. Furthermore, studies have identified that high levels of galactose also induce an accelerated aging process, possibly due to the production of ROS and AGEs [41]. Additionally, it has been reported that both AGEs and ROS are involved in the inflammation onset through the action of the nuclear factor (NF)-κB gene [5]. On the other hand, a study conducted in cultured mammalian cells, HEK293 and HepG2, indicated that galactose induces the accumulation of fructose-6-phosphate that is necessary for the N-glycosylation process. This contributes to a reduction in starvation-induced endoplasmic reticulum stress, emphasizing the contribution of galactose to other pathways rather than energy production [42]. Interestingly, a recent study indicated that in the diet of 3-week-old, postweaning mice, the substitution of glucose for galactose (1:1, mimicking lactose) instead of glucose alone for 3 weeks, prior to an HFD for 9 weeks, was enough to reduce the levels of circulating leptin when compared to glucose alone, although no differences were found in white AT leptin receptor expression, especially in the female mice [43]. Despite the widespread use of these sugars in the Western diet, more studies are required to understand the impacts of lactose and galactose on human metabolism and tissue physiology. It is crucial to elucidate their underlying physiologic mechanisms of action, especially those of tissue-specific metabolism and inflammation. Furthermore, clinical trials are necessary to observe the impacts of these sugars on the organism, since the majority of the published studies were performed in vitro or on animal models.References
- Kopp, W. How Western Diet and Lifestyle Drive the Pandemic of Obesity and Civilization Diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236.
- Amano, T.; Watanabe, D.; Otsuka, J.; Okamoto, Y.; Takada, S.; Fujii, N.; Kenny, G.P.; Enoki, Y.; Maejima, D. Comparison of hydration efficacy of carbohydrate-electrolytes beverages consisting of isomaltulose and sucrose in healthy young adults: A randomized crossover trial. Physiol. Behav. 2022, 249, 113770.
- Arias-Chávez, D.J.; Mailloux-Salinas, P.; Altamirano, J.; Huang, F.; Gómez-Viquez, N.L.; Bravo, G. Consumption of combined fructose and sucrose diet exacerbates oxidative stress, hypertrophy and CaMKIIδ oxidation in hearts from rats with metabolic syndrome. Mol. Cell. Biochem. 2022, 477, 1309–1320.
- Butterworth, P.J.; Bajka, B.H.; Edwards, C.H.; Warren, F.J.; Ellis, P.R. Enzyme kinetic approach for mechanistic insight and predictions of in vivo starch digestibility and the glycaemic index of foods. Trends Food Sci. Technol. 2022, 120, 254–264.
- Omar, N.A.M.; Frank, J.; Kruger, J.; Bello, F.D.; Medana, C.; Collino, M.; Zamaratskaia, G.; Michaelsson, K.; Wolk, A.; Landberg, R. Effects of High Intakes of Fructose and Galactose, with or without Added Fructooligosaccharides, on Metabolic Factors, Inflammation, and Gut Integrity in a Rat Model. Mol. Nutr. Food Res. 2021, 65, e2001133.
- Drake, I.; Sonestedt, E.; Ericson, U.; Wallström, P.; Orho-Melander, M. A Western dietary pattern is prospectively associated with cardio-metabolic traits and incidence of the metabolic syndrome. Br. J. Nutr. 2018, 119, 1168–1176.
- Vasbinder, A.; Anderson, E.; Shadid, H.; Berlin, H.; Pan, M.; Azam, T.U.; Khaleel, I.; Padalia, K.; Meloche, C.; O’Hayer, P.; et al. Inflammation, Hyperglycemia, and Adverse Outcomes in Individuals With Diabetes Mellitus Hospitalized for COVID-19. Diabetes Care 2022, 45, 692–700.
- Pepin, A.; Stanhope, K.L.; Imbeault, P. Are Fruit Juices Healthier Than Sugar-Sweetened Beverages? A Review. Nutrients 2019, 11, 1006.
- Febbraio, M.A.; Karin, M. “Sweet death”: Fructose as a metabolic toxin that targets the gut-liver axis. Cell Metab. 2021, 33, 2316–2328.
- McCain, H.; Kaliappan, S.; Drake, M. Invited review: Sugar reduction in dairy products. J. Dairy Sci. 2018, 101, 8619–8640.
- Patkar, O.L.; Mohamed, A.Z.; Narayanan, A.; Mardon, K.; Cowin, G.; Bhalla, R.; Stimson, D.H.R.; Kassiou, M.; Beecher, K.; Belmer, A.; et al. A binge high sucrose diet provokes systemic and cerebral inflammation in rats without inducing obesity. Sci. Rep. 2021, 11, 11252.
- Malik, V.S.; Popkin, B.M.; Bray, G.A.; Després, J.P.; Hu, F.B. Sugar-sweetened beverages, obesity, type 2 diabetes mellitus, and cardiovascular disease risk. Circulation 2010, 121, 1356–1364.
- Shendurse, A.M.; Khedkar, C.D. Lactose. Encycl. Food Health 2016, 509–516.
- Meyers, A.M.; Mourra, D.; Beeler, J.A. High fructose corn syrup induces metabolic dysregulation and altered dopamine signaling in the absence of obesity. PLoS ONE 2017, 12, e0190206.
- Taskinen, M.-R.; Packard, C.J.; Borén, J. Dietary Fructose and the Metabolic Syndrome. Nutrients 2019, 11, 1987.
- Krycer, J.R.; Quek, L.-E.; Francis, D.; Zadoorian, A.; Weiss, F.C.; Cooke, K.C.; Nelson, M.E.; Diaz-Vegas, A.; Humphrey, S.J.; Scalzo, R.; et al. Insulin signaling requires glucose to promote lipid anabolism in adipocytes. J. Biol. Chem. 2020, 295, 13250–13266.
- Blanco, A.; Blanco, G. Carbohydrates. In Medical Biochemistry; Elsevier: Amsterdam, Netherlands, 2022; pp. 77–103.
- Olofsson, C.; Eriksson, M.; Helin, A.-C.B.; Anderstam, B.; Orsini, N.; Stenvinkel, P.; Ekberg, N.R. Effects of Acute Fructose Loading on Markers of Inflammation—A Pilot Study. Nutrients 2021, 13, 3110.
- Hannou, S.A.; Haslam, D.E.; McKeown, N.M.; Herman, M.A. Fructose metabolism and metabolic disease. J. Clin. Investig. 2018, 128, 545–555.
- Legeza, B.; Marcolongo, P.; Gamberucci, A.; Varga, V.; Bánhegyi, G.; Benedetti, A.; Odermatt, A. Fructose, Glucocorticoids and Adipose Tissue: Implications for the Metabolic Syndrome. Nutrients 2017, 9, 426.
- Stanhope, K.L.; Schwarz, J.M.; Keim, N.L.; Griffen, S.C.; Bremer, A.A.; Graham, J.L.; Hatcher, B.; Cox, C.L.; Dyachenko, A.; Zhang, W.; et al. Consuming fructose-sweetened, not glucose-sweetened, beverages increases visceral adiposity and lipids and decreases insulin sensitivity in overweight/obese humans. J. Clin. Investig. 2009, 119, 1322–1334.
- Varma, V.; Boros, L.G.; Nolen, G.T.; Chang, C.-W.; Wabitsch, M.; Beger, R.D.; Kaput, J. Metabolic fate of fructose in human adipocytes: A targeted 13C tracer fate association study. Metabolomics 2015, 11, 529–544.
- Varma, V.; Boros, L.G.; Nolen, G.T.; Chang, C.-W.; Wabitsch, M.; Beger, R.D.; Kaput, J. Fructose Alters Intermediary Metabolism of Glucose in Human Adipocytes and Diverts Glucose to Serine Oxidation in the One–Carbon Cycle Energy Producing Pathway. Metabolites 2015, 5, 364–385.
- Tedeschi, P.M.; Markert, E.K.; Gounder, M.; Lin, H.; Dvorzhinski, D.; Dolfi, S.C.; Chan, L.L.Y.; Qiu, J.; DiPaola, R.S.; Hirshfield, K.M.; et al. Contribution of serine, folate and glycine metabolism to the ATP, NADPH and purine requirements of cancer cells. Cell Death Dis. 2013, 4, e877.
- Pektas, M.B.; Koca, H.B.; Sadi, G.; Akar, F. Dietary Fructose Activates Insulin Signaling and Inflammation in Adipose Tissue: Modulatory Role of Resveratrol. BioMed Res. Int. 2016, 2016, 8014252.
- Rodrigues, D.F.; Henriques, M.C.D.C.; Oliveira, M.C.; Menezes-Garcia, Z.; Marques, P.E.; Souza, D.D.G.; Menezes, G.B.; Teixeira, M.M.; Ferreira, A.V.M. Acute intake of a high-fructose diet alters the balance of adipokine concentrations and induces neutrophil influx in the liver. J. Nutr. Biochem. 2014, 25, 388–394.
- Zhang, D.-M.; Jiao, R.-Q.; Kong, L.-D. High Dietary Fructose: Direct or Indirect Dangerous Factors Disturbing Tissue and Organ Functions. Nutrients 2017, 9, 335.
- DiNicolantonio, J.J.; Mehta, V.; Onkaramurthy, N.; O’Keefe, J.H. Fructose-induced inflammation and increased cortisol: A new mechanism for how sugar induces visceral adiposity. Prog. Cardiovasc. Dis. 2018, 61, 3–9.
- Marek, G.; Pannu, V.; Shanmugham, P.; Pancione, B.; Mascia, D.; Crosson, S.; Ishimoto, T.; Sautin, Y.Y. Adiponectin Resistance and Proinflammatory Changes in the Visceral Adipose Tissue Induced by Fructose Consumption via Ketohexokinase-Dependent Pathway. Diabetes 2015, 64, 508–518.
- Kovačević, S.; Brkljačić, J.; Milutinović, D.V.; Gligorovska, L.; Bursać, B.; Elaković, I.; Djordjevic, A. Fructose Induces Visceral Adipose Tissue Inflammation and Insulin Resistance Even Without Development of Obesity in Adult Female but Not in Male Rats. Front. Nutr. 2021, 8, 749328.
- Wang, Y.; Qi, W.; Song, G.; Pang, S.; Peng, Z.; Li, Y.; Wang, P. High-Fructose Diet Increases Inflammatory Cytokines and Alters Gut Microbiota Composition in Rats. Mediat. Inflamm. 2020, 2020, 6672636.
- Alkhouri, N.; Dixon, L.J.; Feldstein, A.E. Lipotoxicity in nonalcoholic fatty liver disease: Not all lipids are created equal. Expert Rev. Gastroenterol. Hepatol. 2009, 3, 445–451.
- Della Corte, K.W.; Perrar, I.; Penczynski, K.J.; Schwingshackl, L.; Herder, C.; Buyken, A.E. Effect of dietary sugar intake on biomarkers of subclinical inflammation: A systematic review and meta-analysis of intervention studies. Nutrients 2018, 10, 606.
- Jaiswal, N.; Agrawal, S.; Agrawal, A. High fructose-induced metabolic changes enhance inflammation in human dendritic cells. Clin. Exp. Immunol. 2019, 197, 237–249.
- Pimentel, G.; Burton, K.J.; Rosikiewicz, M.; Freiburghaus, C.; von Ah, U.; Münger, L.H.; Pralong, F.P.; Vionnet, N.; Greub, G.; Badertscher, R.; et al. Blood lactose after dairy product intake in healthy men. Br. J. Nutr. 2017, 118, 1070–1077.
- Conte, F.; van Buuringen, N.; Voermans, N.C.; Lefeber, D.J. Galactose in human metabolism, glycosylation and congenital metabolic diseases: Time for a closer look. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2021, 1865, 129898.
- Krycer, J.R.; Elkington, S.D.; Diaz-Vegas, A.; Cooke, K.C.; Burchfield, J.G.; Fisher-Wellman, K.H.; Cooney, G.J.; Fazakerley, D.J.; James, D.E. Mitochondrial oxidants, but not respiration, are sensitive to glucose in adipocytes. J. Biol. Chem. 2020, 295, 99–110.
- Krishna, M.S.; Revathy, V.M.; Jaleel, A. Adipocytes utilize sucrose as an energy source—Effect of different carbohydrates on adipocyte differentiation. J. Cell. Physiol. 2020, 235, 891–899.
- Remigante, A.; Morabito, R.; Spinelli, S.; Trichilo, V.; Loddo, S.; Sarikas, A.; Dossena, S.; Marino, A. D-Galactose Decreases Anion Exchange Capability through Band 3 Protein in Human Erythrocytes. Antioxidants 2020, 9, 689.
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552.
- Umbayev, B.; Askarova, S.; Almabayeva, A.; Saliev, T.; Masoud, A.-R.; Bulanin, D. Galactose-Induced Skin Aging: The Role of Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 7145656.
- Sasaoka, N.; Imamura, H.; Kakizuka, A. A Trace Amount of Galactose, a Major Component of Milk Sugar, Allows Maturation of Glycoproteins during Sugar Starvation. Iscience 2018, 10, 211–221.
- Bouwman, L.M.S.; Fernández-Calleja, J.M.S.; van der Stelt, I.; Oosting, A.; Keijer, J.; van Schothorst, E.M. Replacing Part of Glucose with Galactose in the Postweaning Diet Protects Female But Not Male Mice from High-Fat Diet–Induced Adiposity in Later Life. J. Nutr. 2019, 149, 1140–1148.
More