Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by Alfonsina Gattuso.
Hypoxia imposes conflicting demands on cardio-respiratory function. Being systemically O2 supply-dependent on cardiac output (CO) and arterial O2 concentration, fish can respond to and cope with hypoxia through cardio-respiratory adjustments to preserve systemic O2 delivery, thus maintaining aerobic metabolism, or by reducing O2 demands via anaerobic metabolism or metabolic depression. In 1986, Hochachka firstly proposed metabolic arrest, i.e., a simultaneous reduction in metabolic rate and metabolic demands, as a key adaptation to O2 deprivation in organisms capable of long-term anoxic survival.
- fish heart
- cyprinids
- contractility
- metabolism
1. Introduction
Oxygen fluctuations are a common experience for species living in water environments, since they naturally occur over a diurnal/seasonal rate and are exacerbated by eventual anthropic manipulation that challenge the O2 budget. Exposure to natural O2 variations potently shaped the evolution of a number of adaptive strategies that, in fish, require behavioral, morphological and functional modifications. Reaching the surface to breathe the uppermost layer of water in contact with air, increasing the activity to avoid the hypoxic area, or decreasing the activity to reduce O2 demand, are amongst the most common behavioral responses to low O2 [1,2][1][2]. Beyond them, changes in ventilation and hemoglobin–O2 binding [3] contribute to ameliorate O2 extraction from the environment in order to maintain aerobic ATP production.
Fundamental for fish adaptation to restricted O2 is a proper availability of metabolic fuels, obtained either by reducing energy consumption or increasing substrates extraction from energy stores, or both. This is accompanied by an appropriate blood supply for cells and tissues, provided by the compensatory adaptation of both the heart and the circulatory system. All the above responses are supported by cellular and molecular adaptive rearrangements that contribute to an orchestrated framework of events allowing to preserve body functions, while at the same time protecting from the risk of metabolic impairment.
At the extremes of the large spectrum of adaptation to O2 availability, several fish species evolved the ability to survive even in the presence of O2 below the critical tension (Pcrit), thus tolerating prolonged hypoxia and/or anoxia [3,4,5][3][4][5]. An example is represented by the teleost belonging to cyprinids, which are champions of hypoxia/anoxia tolerance. For this peculiarity, there is a continuously growing interest to analyze the physiological mechanisms that, at a different degree of biological organization, make cyprinid species able to cope with restrictions in O2.
 The cardiac response of fish to low O2 requires an analysis in relation to fuel substrates. Carbohydrates represent the energy source of choice for the heart of several fish species [54][20]. In the isolated and perfused eel heart, glucose supply maintains the cardiac performance during acute anoxia (see references in [55][21]) while, in the American eel Anguilla rostrata, characterized by a marked anoxic endurance, the inhibition of oxidative phosphorylation with NaCN activates glycogen stores degradation regardless of glucose levels in the medium [56][22]. Prolonged survival under anoxia requires large stores of fermentable substrate (normally glycogen), whose conservation is facilitated by a strong metabolic depression [44][7]. In the anoxic crucian carp, cardiac glycogen stores are quickly mobilized during the first week of anoxia with little further degradation when anoxia is protracted to 3 and 6 weeks [57][23]. This suggests that, after the first week of anoxia, the heart performance relies on exogenous glucose. Of note, glycogen depletion is not paralleled by increased glucose or lactate concentration, the latter even reduced if compared to the normoxic control [57][23]. It has been proposed that the early mobilization of glycogen (and other glycolytic intermediates) is not an advantage to the fish in relation to its anoxia tolerance, but it is crucial for adapting body fluids osmolarity, which is perturbed in the presence of an increase in body mass (6.2%) occurring under anoxia [57][23]. An accumulation of glycolytic intermediates has also been detected in the cardiac and skeletal muscle of anoxic goldfish [58][24]. Protracted hypoxia (2.1 kPa for 4 weeks) suppresses goldfish metabolic rate by 74% [59][25] in the whole animal with no direct effects on the heart, which retains a normal mitochondrial respiration rate [60][26]; carbohydrates represent the election fuel in maintaining mitochondrial respiration [60][26]. This apparently contrasts with in vitro data showing that, on ventricular strips from goldfish, hypoxia depresses myocardial contractility and O2 consumption rate [61][27]. However, this does not significantly change O2 utilization capacity (i.e., the ratio of twitch force to O2 consumption), and this may reflect a lower activation of anaerobic energy production. This suggests that the goldfish heart is able to maintain a higher degree of aerobic metabolism at low O2 tensions without increasing anaerobic energy production [61][27].
Because of their ability to detect changes in O2 availability, mitochondria are known for their role in coordinating the responses to low O2 [62][28]. Mitochondrial respiration is differently affected by hypoxia acclimation depending on metabolic fuels, species, and tissue. In fish, many studies on the effects on mitochondrial function under hypoxia and/or anoxia-reoxygenation have been mainly performed on muscle and liver tissues [63,64,65][29][30][31]. Only a few investigations examined the effects of chronic hypoxia on cardiac mitochondrial function, providing conflicting information. By using permeabilized cardiac fibers and isolated mitochondria, Cook et al. [66][32] showed no effect on complex I and II respiration in juvenile snapper (Pagrus auratus) acclimated to 10.2–12.1 kPa for 6 weeks. In contrast, oxidative phosphorylation decreased in permeabilized ventricle fibers from the hypoxia-intolerant shovelnose ray (Aptychotrema rostrata) following a 2 h in vivo hypoxic insult, while it was preserved in the hypoxia-tolerant epaulette shark (Hemiscyllum ocellatum) [67][33]. In the hypoxia-tolerant sablefish (Anoplopoma fimbria Pallas), cardiac mitochondrial respiration was maintained following exposure to chronic hypoxia (8 ± 1 kPa for 6 months) and similar P50 values between normoxic and hypoxic groups suggest that the O2 dependence of complex IV, the primary site of O2 consumption in the mitochondrion, is not affected by low O2 acclimation [68][34]. In addition, the cardiac activity of citrate synthase, a marker of oxidative capacity, increases in both ventricular homogenates and mitochondrial suspension of hypoxia-acclimated sablefish [68][34], suggesting that a sustained cardiac mitochondrial capacity primarily involves changes in the intrinsic properties of the mitochondria and not in the abundance of these organelles [69][35]. In the goldfish, chronic hypoxia decreased COX activity in different tissues, except for the heart [60][26].
An increased activity of the oxidative phosphorylation enzymes often coincides with an enhancement of the percentage of cell volume displaced by mitochondria [70][36]. Different stimuli may affect the mitochondrial compartment in fish. Examples are the increased mitochondrial density documented in response to cold acclimation in the oxidative muscle fibers of European eel (Anguilla anguilla), striped bass (Morone saxatilis), crucian carp (C. carassius), goldfish (C. auratus) and stickleback (Gasterosteus aculeatus) [70][36], as well as in the hypertrophic zebrafish heart in response to humoral stimulation by angiotensin II [71][37], and in the ventricle of the European eel during ontogenetic growth [72][38]. Of note, in the hypoxia-acclimated goldfish heart, a modulation of transcripts coding for mitochondrial fission (fis1) and fusion (mfn1 and mfn2) proteins has been reported [73][39]. Specifically, 1-week exposure to hypoxia elicits a significant reduction in the relative transcript abundance of mitofusin mfn1 and an increase in the mitochondrial fission factor fis1 [73][39], calling for activation of the fission process. On the contrary, protracting hypoxia to 4 weeks, the relative transcript abundance of mitochondrial fusion and fission proteins is restored at normoxic values [73][39]. This modulation of mitochondria dynamics allows for the hypothesis that in the early phase of hypoxia adaptation, an augmented mitochondrial density, due to the activation of fission events, may maximize energy delivery to the contractile apparatus needed to sustain the enhanced pumping behavior of the heart [31][10]. In contrast, if hypoxia is protracted, mitochondrial fusion occurs and promotes mitochondrial membrane stability to protect mitochondria from damage, mitophagy and the induction of cellular apoptosis [73][39] (Figure 1).
The cardiac response of fish to low O2 requires an analysis in relation to fuel substrates. Carbohydrates represent the energy source of choice for the heart of several fish species [54][20]. In the isolated and perfused eel heart, glucose supply maintains the cardiac performance during acute anoxia (see references in [55][21]) while, in the American eel Anguilla rostrata, characterized by a marked anoxic endurance, the inhibition of oxidative phosphorylation with NaCN activates glycogen stores degradation regardless of glucose levels in the medium [56][22]. Prolonged survival under anoxia requires large stores of fermentable substrate (normally glycogen), whose conservation is facilitated by a strong metabolic depression [44][7]. In the anoxic crucian carp, cardiac glycogen stores are quickly mobilized during the first week of anoxia with little further degradation when anoxia is protracted to 3 and 6 weeks [57][23]. This suggests that, after the first week of anoxia, the heart performance relies on exogenous glucose. Of note, glycogen depletion is not paralleled by increased glucose or lactate concentration, the latter even reduced if compared to the normoxic control [57][23]. It has been proposed that the early mobilization of glycogen (and other glycolytic intermediates) is not an advantage to the fish in relation to its anoxia tolerance, but it is crucial for adapting body fluids osmolarity, which is perturbed in the presence of an increase in body mass (6.2%) occurring under anoxia [57][23]. An accumulation of glycolytic intermediates has also been detected in the cardiac and skeletal muscle of anoxic goldfish [58][24]. Protracted hypoxia (2.1 kPa for 4 weeks) suppresses goldfish metabolic rate by 74% [59][25] in the whole animal with no direct effects on the heart, which retains a normal mitochondrial respiration rate [60][26]; carbohydrates represent the election fuel in maintaining mitochondrial respiration [60][26]. This apparently contrasts with in vitro data showing that, on ventricular strips from goldfish, hypoxia depresses myocardial contractility and O2 consumption rate [61][27]. However, this does not significantly change O2 utilization capacity (i.e., the ratio of twitch force to O2 consumption), and this may reflect a lower activation of anaerobic energy production. This suggests that the goldfish heart is able to maintain a higher degree of aerobic metabolism at low O2 tensions without increasing anaerobic energy production [61][27].
Because of their ability to detect changes in O2 availability, mitochondria are known for their role in coordinating the responses to low O2 [62][28]. Mitochondrial respiration is differently affected by hypoxia acclimation depending on metabolic fuels, species, and tissue. In fish, many studies on the effects on mitochondrial function under hypoxia and/or anoxia-reoxygenation have been mainly performed on muscle and liver tissues [63,64,65][29][30][31]. Only a few investigations examined the effects of chronic hypoxia on cardiac mitochondrial function, providing conflicting information. By using permeabilized cardiac fibers and isolated mitochondria, Cook et al. [66][32] showed no effect on complex I and II respiration in juvenile snapper (Pagrus auratus) acclimated to 10.2–12.1 kPa for 6 weeks. In contrast, oxidative phosphorylation decreased in permeabilized ventricle fibers from the hypoxia-intolerant shovelnose ray (Aptychotrema rostrata) following a 2 h in vivo hypoxic insult, while it was preserved in the hypoxia-tolerant epaulette shark (Hemiscyllum ocellatum) [67][33]. In the hypoxia-tolerant sablefish (Anoplopoma fimbria Pallas), cardiac mitochondrial respiration was maintained following exposure to chronic hypoxia (8 ± 1 kPa for 6 months) and similar P50 values between normoxic and hypoxic groups suggest that the O2 dependence of complex IV, the primary site of O2 consumption in the mitochondrion, is not affected by low O2 acclimation [68][34]. In addition, the cardiac activity of citrate synthase, a marker of oxidative capacity, increases in both ventricular homogenates and mitochondrial suspension of hypoxia-acclimated sablefish [68][34], suggesting that a sustained cardiac mitochondrial capacity primarily involves changes in the intrinsic properties of the mitochondria and not in the abundance of these organelles [69][35]. In the goldfish, chronic hypoxia decreased COX activity in different tissues, except for the heart [60][26].
An increased activity of the oxidative phosphorylation enzymes often coincides with an enhancement of the percentage of cell volume displaced by mitochondria [70][36]. Different stimuli may affect the mitochondrial compartment in fish. Examples are the increased mitochondrial density documented in response to cold acclimation in the oxidative muscle fibers of European eel (Anguilla anguilla), striped bass (Morone saxatilis), crucian carp (C. carassius), goldfish (C. auratus) and stickleback (Gasterosteus aculeatus) [70][36], as well as in the hypertrophic zebrafish heart in response to humoral stimulation by angiotensin II [71][37], and in the ventricle of the European eel during ontogenetic growth [72][38]. Of note, in the hypoxia-acclimated goldfish heart, a modulation of transcripts coding for mitochondrial fission (fis1) and fusion (mfn1 and mfn2) proteins has been reported [73][39]. Specifically, 1-week exposure to hypoxia elicits a significant reduction in the relative transcript abundance of mitofusin mfn1 and an increase in the mitochondrial fission factor fis1 [73][39], calling for activation of the fission process. On the contrary, protracting hypoxia to 4 weeks, the relative transcript abundance of mitochondrial fusion and fission proteins is restored at normoxic values [73][39]. This modulation of mitochondria dynamics allows for the hypothesis that in the early phase of hypoxia adaptation, an augmented mitochondrial density, due to the activation of fission events, may maximize energy delivery to the contractile apparatus needed to sustain the enhanced pumping behavior of the heart [31][10]. In contrast, if hypoxia is protracted, mitochondrial fusion occurs and promotes mitochondrial membrane stability to protect mitochondria from damage, mitophagy and the induction of cellular apoptosis [73][39] (Figure 1).
2. Hypoxia-Related Metabolic Responses of the Fish Heart
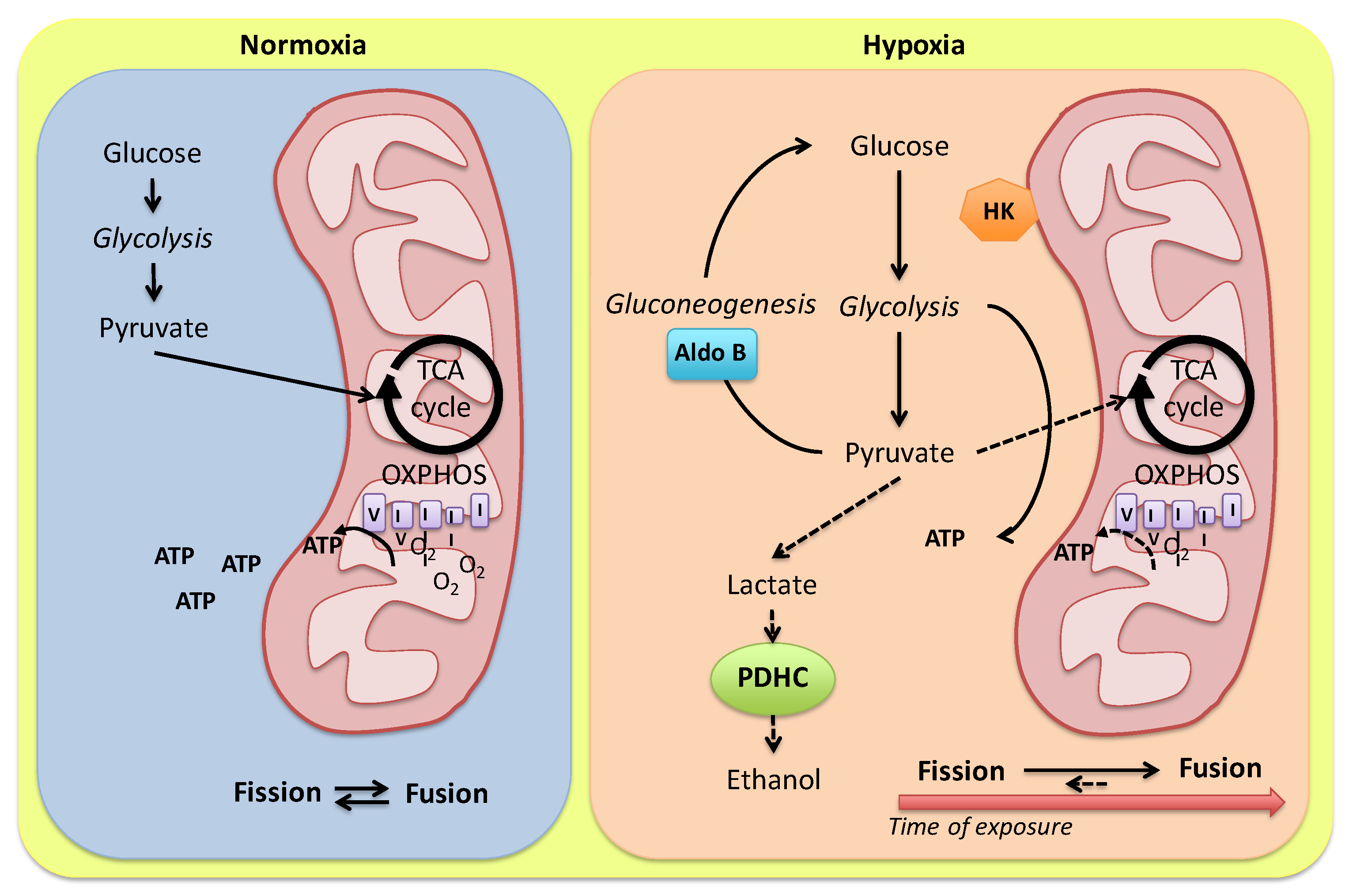
In 1986, Hochachka firstly proposed metabolic arrest, i.e., a simultaneous reduction in metabolic rate and metabolic demands, as a key adaptation to O2 deprivation in organisms capable of long-term anoxic survival [43][6]. Contrary to the activation of the anaerobic pathway to sustain ATP production and maintain aerobic respiratory rates, the reduction in energetic demand, which clues to an overall reduction in ATP turnover, preserves glycogen stores and avoids the accumulation of waste products (i.e., acid lactic production) which may rapidly lead to a Pasteur effect. Thus, organisms tolerating long-term anoxia lack a Pasteur effect [44][7]. In fact, they do not increase glycolytic ATP production to maintain aerobic respiratory rates. This general concept has endured over time and is corroborated by new findings related to the identification of alternative metabolic pathways which allow a switch to anaerobic metabolism keeping low waste product accumulation. The extreme is exemplified by the capacity of cyprinid fish to tolerate prolonged O2 absence by using large glycogen stores to generate ethanol as a by-product of energy metabolism, thus avoiding acidosis [45][8]. This extraordinary capacity is due to the presence in Carassius genus (C. carassius and C. auratus) of an alternative E1 pyruvate dehydrogenase enzyme, one of the catalytic components of the pyruvate dehydrogenase complex (PDHC) which, under anoxia, functions as an acetaldehyde-producing mitochondrial pyruvate decarboxylase (PDC) analogous to the cytosolic pyruvate decarboxylase in brewer’s yeast [46][9]. This isoform derives from an additional set of paralogs for each of the E1α and E1β sub-units, originating from a cyprinid-specific paleotetraploidization event occurring approximately 8.2 million years ago in a common ancestor of the Carassius genus (anoxia tolerant) and the common carp (anoxia intolerant) [46][9]. While one pair maintained the original function (i.e., catalyzing the synthesis of acetyl-CoA during normoxia as an integral part of PDHC), the other pair has apparently evolved into an E1 enzyme physically independent of PDHC, catalyzing the formation of acetaldehyde in anoxia, which then can be effectively converted into ethanol by a muscle-specific alcohol dehydrogenase (ADH). Authors [46][9] reported a tissue-specific distribution of PDHC sub-units with E1α3, E1β2, and E2a transcripts dominating in ethanol-producing red and white skeletal muscle, and E1α1 or E1α2, E1β1, and E2b transcripts in heart, brain, and liver, with expression levels lower than muscle; this suggests a minor role for these tissues in ethanol production under anoxia. Nonetheless, the heart of these species conserve normal cardiac activity if exposed to acute hypoxia (C. auratus: [31][10]), or up to 5 days of anoxia (C. carassius: [28][11]). A preserved heart performance is the basis for improving anoxia resistance of the whole piscine organism since it ensures metabolic and functional cooperation among single organs [47][12]. In this view, an appropriate perfusion of organ and tissues guarantees lactate transport to the muscle for its conversion into ethanol and, the latter, to the gills for excretion. The Carassius ability to maintain routine cardiac activity during anoxia implies a cardiac ATP demand which is lower than their maximum glycolytic potential [29][13], thus protecting the heart from the accumulation of anaerobic waste products. In the goldfish, the enhancement of myocardial contractility in response to low O2 is associated with low amounts of cardiac lactate together with a slight reduction in pyruvate levels [48][14]. By using mass spectrometry-based proteomic analysis, reseauthorchers identified two isoforms of fructose-bisphosphate aldolase, i.e., aldolase C and aldolase B, differently expressed in homogenates of goldfish heart exposed to normoxic or hypoxic medium, with aldolase B predominantly expressed in the hypoxic heart [48][14]. Aldolase catalyzes the reversible conversion of fructose-1,6-bisphosphate to glyceraldehyde 3-phosphate (G3P) and dihydroxyacetone phosphate (DHAP). While aldolase C appears to be more effective in participating in glycolysis, aldolase B has evolved to have a role in gluconeogenesis [49,50][15][16]. This supports the possibility that in the goldfish exposed to reduced O2, a tight modulation of the aldolase enzyme isoforms may finely regulate glycolytic vs. gluconeogenic flux, thus enhancing anaerobic ATP yield and minimizing metabolic acidosis [48][14]. Of note, under O2 limitation, a number of glycolytic enzymes show increased binding to subcellular components, particularly mitochondria [51][17] or the particulate fraction, and this is proposed to finely regulate glycolytic flux rates through the modulation of enzyme-specific kinetics [52][18] (Figure 1). Examples are represented by the increased binding of hexokinase to mitochondria observed in the heart of goldfish maintained in anoxic water [53][19], as well as the increased binding capacity of phosphofructokinase, aldolase, and pyruvate kinase to the particulate fraction observed in ventricular sheets of armored catfish (Liposarcus pardalis) exposed to hypoxia [52][18].Figure 1. Model for alternative routes of pyruvate metabolism in the Carassius auratus heart. In the presence of O2, pyruvate from glycolysis is converted to acetyl-CoA and addressed to mitochondria for the oxidative phosphorylation. Under hypoxia, a tight modulation of glycolysis enzymes may finely regulate anaerobic ATP production by modulating glycolytic vs. gluconeogenic flux. Pyruvate conversion to ethanol is reduced by a low cardiac expression of alternative PDHC sub-units (see the text for details). A hypoxia-dependent modulation of mitochondria dynamics in relation to the exposure time is proposed. HK: hexokinase; Aldo B: aldolase B; PDHC: pyruvate dehydrogenase complex.
References
- Chapman, L.J.; Mckenzie, D.J. Behavioral responses and ecological consequences. In Fish Physiology; Elsevier: Amsterdam, The Netherlands, 2009; Volume 27, pp. 25–77.
- Urbina, M.A.; Forster, M.E.; Glover, C.N. Leap of faith: Voluntary emersion behaviour and physiological adaptations to aerial exposure in a non-aestivating freshwater fish in response to aquatic hypoxia. Physiol. Behav. 2011, 103, 240–247.
- Richards, J.G. Metabolic and molecular responses of fish to hypoxia. In Fish Physiology; Jeffrey, G., Richards, A.P.F., Colin, J.B., Eds.; Academic Press: Cambridge, MA, USA, 2009; Volume 27, pp. 443–485.
- Rogers, N.J.; Urbina, M.A.; Reardon, E.E.; McKenzie, D.J.; Wilson, R.W. A new analysis of hypoxia tolerance in fishes using a database of critical oxygen level (P crit). Conserv. Physiol. 2016, 4, cow012.
- Ultsch, G.R.; Regan, M.D. The utility and determination of P(crit) in fishes. J. Exp. Biol. 2019, 222, jeb203646.
- Hochachka, P.W. Defense strategies against hypoxia and hypothermia. Science 1986, 231, 234–241.
- Bickler, P.E.; Buck, L.T. Hypoxia tolerance in reptiles, amphibians, and fishes: Life with variable oxygen availability. Annu. Rev. Physiol. 2007, 69, 145–170.
- Shoubridge, E.A.; Hochachka, P.W. Ethanol: Novel end product of vertebrate anaerobic metabolism. Science 1980, 209, 308–309.
- Fagernes, C.E.; Stenslokken, K.O.; Rohr, A.K.; Berenbrink, M.; Ellefsen, S.; Nilsson, G.E. Extreme anoxia tolerance in crucian carp and goldfish through neofunctionalization of duplicated genes creating a new ethanol-producing pyruvate decarboxylase pathway. Sci. Rep. 2017, 7, 7884.
- Imbrogno, S.; Capria, C.; Tota, B.; Jensen, F.B. Nitric oxide improves the hemodynamic performance of the hypoxic goldfish (Carassius auratus) heart. Nitric Oxide 2014, 42, 24–31.
- Stecyk, J.A.; Stenslokken, K.O.; Farrell, A.P.; Nilsson, G.E. Maintained cardiac pumping in anoxic crucian carp. Science 2004, 306, 77.
- Gattuso, A.; Garofalo, F.; Cerra, M.C.; Imbrogno, S. Hypoxia Tolerance in Teleosts: Implications of Cardiac Nitrosative Signals. Front. Physiol. 2018, 9, 366.
- Farrell, A.P.; Stecyk, J.A. The heart as a working model to explore themes and strategies for anoxic survival in ectothermic vertebrates. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 300–312.
- Imbrogno, S.; Aiello, D.; Filice, M.; Leo, S.; Mazza, R.; Cerra, M.C.; Napoli, A. MS-based proteomic analysis of cardiac response to hypoxia in the goldfish (Carassius auratus). Sci. Rep. 2019, 9, 18953.
- Penhoet, E.E.; Kochman, M.; Rutter, W.J. Molecular and catalytic properties of aldolase C. Biochemistry 1969, 8, 4396–4402.
- Penhoet, E.E.; Rutter, W.J. Catalytic and immunochemical properties of homomeric and heteromeric combinations of aldolase subunits. J. Biol. Chem. 1971, 246, 318–323.
- Brooks, S.P.J.; Storey, K.B. Is glycolytic rate controlledby the reversible binding of enzymes to subcellular structures? In Biochemistry and Molecular Biology of Fishes; Hochachka, P.W., Mommsen, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 1995; Volume 4, pp. 291–307.
- Treberg, J.R.; MacCormack, T.J.; Lewis, J.M.; Almeida-Val, V.M.; Val, A.L.; Driedzic, W.R. Intracellular glucose and binding of hexokinase and phosphofructokinase to particulate fractions increase under hypoxia in heart of the amazonian armored catfish (Liposarcus pardalis). Physiol. Biochem. Zool. 2007, 80, 542–550.
- Duncan, J.A.; Storey, K.B. Role of enzyme binding in muscle metabolism of the goldfish. Can. J. Zool. 1991, 69, 1571–1576.
- Sidell, B.D.; Stowe, D.B.; Hansen, C.A. Carbohydrate Is the Preferred Metabolic Fuel of the Hagfish (Myxine glutinosa) Heart. Physiol. Zool. 1984, 57, 266–273.
- Imbrogno, S. The eel heart: Multilevel insights into functional organ plasticity. J. Exp. Biol. 2013, 216, 3575–3586.
- Bailey, J.R.; MacDougall, R.; Clowe, S.; Driedzic, W.R. Anoxic performance of the american eel (Anguilla rostrata L.) heart requires extracellular glucose. J. Exp. Zool. 2000, 286, 699–706.
- Vornanen, M.; Haverinen, J. Glycogen dynamics of crucian carp (Carassius carassius) in prolonged anoxia. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2016, 186, 999–1007.
- Shoubridge, E.A.; Hochachka, P.W. The integrationand control of metabolism in the anoxic goldfish. Mol. Physiol. 1983, 4, 165–195.
- Farhat, E.; Turenne, E.D.; Choi, K.; Weber, J.M. Hypoxia-induced remodelling of goldfish membranes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 237, 110326.
- Farhat, E.; Cheng, H.; Romestaing, C.; Pamenter, M.; Weber, J.M. Goldfish Response to Chronic Hypoxia: Mitochondrial Respiration, Fuel Preference and Energy Metabolism. Metabolites 2021, 11, 187.
- Pedersen, C.L.; Faggiano, S.; Helbo, S.; Gesser, H.; Fago, A. Roles of nitric oxide, nitrite and myoglobin on myocardial efficiency in trout (Oncorhynchus mykiss) and goldfish (Carassius auratus): Implications for hypoxia tolerance. J. Exp. Biol. 2010, 213, 2755–2762.
- Pamenter, M.E. Mitochondria: A multimodal hub of hypoxia tolerance. Can. J. Zool. 2014, 92, 569–589.
- Du, S.N.N.; Mahalingam, S.; Borowiec, B.G.; Scott, G.R. Mitochondrial physiology and reactive oxygen species production are altered by hypoxia acclimation in killifish (Fundulus heteroclitus). J. Exp. Biol. 2016, 219, 1130–1138.
- Onukwufor, J.O.; Stevens, D.; Kamunde, C. Combined effects of cadmium, temperature and hypoxia-reoxygenation on mitochondrial function in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2017, 182, 129–141.
- Sappal, R.; MacDougald, M.; Fast, M.; Stevens, D.; Kibenge, F.; Siah, A.; Kamunde, C. Alterations in mitochondrial electron transport system activity in response to warm acclimation, hypoxia-reoxygenation and copper in rainbow trout, Oncorhynchus mykiss. Aquat. Toxicol. 2015, 165, 51–63.
- Cook, D.G.; Iftikar, F.I.; Baker, D.W.; Hickey, A.J.R.; Herbert, N.A. Low-O2 acclimation shifts the hypoxia avoidance behaviour of snapper (Pagrus auratus) with only subtle changes in aerobic and anaerobic function. J. Exp. Biol. 2013, 216, 369–378.
- Hickey, A.J.R.; Renshaw, G.M.C.; Speers-Roesch, B.; Richards, J.G.; Wang, Y.; Farrell, A.P.; Brauner, C.J. A radical approach to beating hypoxia: Depressed free radical release from heart fibres of the hypoxia-tolerant epaulette shark (Hemiscyllum ocellatum). J. Comp. Physiol. B 2012, 182, 91–100.
- Gerber, L.; Clow, K.A.; Katan, T.; Emam, M.; Leeuwis, R.H.J.; Parrish, C.C.; Gamperl, A.K. Cardiac mitochondrial function, nitric oxide sensitivity and lipid composition following hypoxia acclimation in sablefish. J. Exp. Biol. 2019, 222, jeb208074.
- St-Pierre, J.; Boutilier, R.G. Aerobic capacity of frog skeletal muscle during hibernation. Physiol. Biochem. Zool. 2001, 74, 390–397.
- O’Brien, K.M. Mitochondrial biogenesis in cold-bodied fishes. J. Exp. Biol. 2011, 214, 275–285.
- Filice, M.; Barca, A.; Amelio, D.; Leo, S.; Mazzei, A.; Del Vecchio, G.; Verri, T.; Cerra, M.C.; Imbrogno, S. Morpho-functional remodelling of the adult zebrafish (Danio rerio) heart in response to waterborne angiotensin II exposure. Gen. Comp. Endocrinol. 2021, 301, 113663.
- Cerra, M.C.; Imbrogno, S.; Amelio, D.; Garofalo, F.; Colvee, E.; Tota, B.; Icardo, J.M. Cardiac morphodynamic remodelling in the growing eel (Anguilla anguilla L.). J. Exp. Biol. 2004, 207, 2867–2875.
- Farhat, E.; Talarico, G.G.M.; Grégoire, M.; Weber, J.M.; Mennigen, J.A. Epigenetic and post-transcriptional repression support metabolic suppression in chronically hypoxic goldfish. Sci. Rep. 2022, 12, 5576.
More