Potassium channels are widely distributed integral proteins responsible for the effective and selective transport of K+ ions through the biological membranes. According to the existing structural and mechanistic differences, they are divided into several groups. All of them are considered important molecular drug targets due to their physiological roles, including the regulation of membrane potential or cell signaling. Among the pharmaceuticals of plant origin, which are potassium channel modulators, flavonoids appear as a powerful group of biologically active substances. It is caused by their well-documented anti-oxidative, anti-inflammatory, anti-mutagenic, anti-carcinogenic, and antidiabetic effects on human health.
- potassium channels
- flavonoids
- potassium channel modulators
1. Introduction
ions per second) and selective transport of K+ ions across the biological membranes down their electrochemical gradient [1]. Although the
channels are related members of one protein family, they can be divided into several groups according to the structural and mechanistic differences [2,3,4,5]. First, one can discern the voltage-regulated channels (Kv) and the
-regulated K+ channels (KCa), being structurally distinguished by six transmembrane domains (TMDs). Another group is formed by the “leak”, which is the double pore K+ channels (K2P) composed of four TMDs. The last group is constituted by the inward rectifier potassium channels (Kir), having two TMDs. The activity of the K+ channels plays a pivotal role in a myriad of cellular processes, for instance, volume regulation, proliferation, hormone secretion, neurotransmitter release, and modulation of potential in electrically excitable and non-excitable cells [1]. Due to their fundamental physiological roles, they appear promising drug targets for many diseases ranging from cardiovascular, metabolic, and neurodegenerative disorders to cancers [6,7,8,9,10,11]. The potential therapeutic benefits of the potassium channels targeting by pharmacological agents depend on the efficacy, strength, and selectivity of the interaction between an active compound and a particular ion channel subtype. It is also challenging to introduce such a channel modulator, which would be delivered or act only within a pathological tissue and would not exhibit any harmful side effects.
Flavonoids comprise a wide group of polyphenolic compounds of plant origin, which exhibit anti-oxidative, anti-inflammatory, anti-mutagenic, and anti-carcinogenic properties, as well as the capability to regulate the functioning of key cellular enzymes [12][13][21,22]. Flavonoids can be divided into subclasses such as flavones, flavonols, flavanols, flavanones, isoflavones, anthocyanidins, and chalcones. Due to their ability to modulate cell physiology, they have a clinically proven positive impact in counteracting a plethora of health problems such as cardiovascular and metabolic diseases, or other inflammation-related pathologies, including cancers.
2. Kv Channels
2.1. Kv1.3 Channel
Kv1.3 channels are activated, as all the other voltage-gated potassium channels, by a change of the membrane potential (membrane depolarization). Although they are expressed in many different tissues of the human body, they play the most prominent role in the T lymphocytes [16][17][50,51] where they are responsible for cell activation. Accordingly, the inhibition of these ion channels results in the inactivation of the T cells, which promotes immune suppression. It makes the Kv1.3 channel a therapeutic target for the treatment of such diseases as sclerosis, type 1 diabetes, and rheumatoid arthritis [6][16][18][6,50,52]. The recent findings suggest that an increased expression of these channels is required to induce apoptosis in the cancer cells. Therefore, the Kv1.3 channels are a new molecular target in both the diagnostics and therapy of some oncological diseases (e.g., breast cancer, colon cancer) [19][53]. Throughout the years, the group of Teisseyre investigated the impact of a large number of different flavonoids on activation of the Kv1.3 channels, both in normal and cancer T cells [19][53]. They discovered the inhibitory effects of genistein, a popular tyrosine kinase inhibitor, which suppresses the activation of the Kv1.3 channel in a concentration-dependent manner with the half-blocking range of IC50 = 30–60 μM [20][25]. These results are consistent with the previous study presented in [21][54]. Moreover, Teisseyre et al. observed that under the influence of daidzein, which is a structural analog of genistein, the ion channel’s activity remains unaffected. They further demonstrated that resveratrol (a non-flavonoid polyphenol) is able to decrease the activity of the Kv1.3. channel. This effect is slowly reversible, and it is exerted in a concentration-dependent manner. At the same time, it turned out that the co-application of this polyphenol with genistein did not significantly change the suppression effect of resveratrol. Since in the previous study [20][25], it was demonstrated that genistein has a similar impact on the Kv1.3 channel as resveratrol, the scholars suggested that the inhibitory effects of these compounds are independent of each other, and they interact with different binding sites of the channel. In the following experiments [22][30], the influence of four flavonoids: aromadendrin, naringenin and its two derivatives, naringenin-4′,7-dimethylether and naringenin-7-methyl-ether, on the Kv1.3 channel activity was investigated in human T lymphocytes isolated from peripheral blood. The single-channel patch-clamp traces revealed that naringenin and aromadendrin did not reduce the Kv1.3 current at the concentration of c = 30 μM. On the contrary, the two investigated derivatives of naringenin enabled us to reduce the ionic currents at the same concentrations, and the most effective inhibitory effect was observed for the naringenin-4′,7-dimethylether. In contrast, in [23][32], it was discovered that 8-prenylnaringenin blocks completely the Kv1.3 channel at c = 10 μM. The presence of the prenyl group is anticipated to promote the inhibitory abilities of this compound. Indeed, a few years later, it was demonstrated that other compounds of such structural characteristics, xanthohumol and isoxanthohumol, are able to effectively suppress the activity of the Kv1.3 channel [24][31]. Although xanthohumol turned out to be slightly more effective in decreasing the ion current, the administration of neither of the investigated compounds led to a complete channel inhibition at the concentration c = 30 μM. Even though xanthohumol and isoxanthohumol are less potent in inhibition of the Kv1.3 channel than 8-prenylnaringenin, they are much more effective than the other natural plant-derived compounds such as genistein and resveratrol. Therefore, these results confirm the hypothesis that prenylated flavonoids are much more effective in blocking the Kv1.3 channel due to the presence of the prenyl group, which facilitates the non-conducting state of the channel.2.2. Kv1.5 Channel
The channels belonging to the Kv1.5 family are expressed in many tissues of the human body [25][26][58,59]. The greatest attention of the scientists is, however, focused on its expression in the heart. It has been demonstrated [27][28][60,61] that the Kv1.5 channel conducts the ultra-rapid delayed rectifier current IKur, which plays an important role in shaping the atrial action potential (AP) repolarization [29][62]. The studies suggest that the inhibition of this channel can contribute to the prolongation of the AP duration and, by this, stop the atrial fibrillation (AF) [30][63]. It is important to note that although the Kv1.5 channel is present in the atria, it is not expressed in the ventricular muscle in the heart. These two factors, i.e., provision of the current driving the AP and selective expression of this ion channel (its presence in the atria and absence in the ventricle), consider the Kv1.5 channels as a potential target for the treatment of the cardiac arrhythmia [31][32][33][64,65,66]. For this reason, it would be useful to find new efficient inhibitors of the Kv1.5 channel. Wang et al. studied the inhibitory effects of the hesperetin on the IKur through the Kv1.5 channels expressed in the HEK 293 cells. It turned out that although externally applied hesperetin can significantly suppress the ultra-rapid delayed rectifier K+ current in the concentration-dependent manner with IC50=23.15μM, the presence of this flavonoid in pipette solution yielded no effect. The scholars concluded that hesperetin interacts with a channel protein only from the exterior. They also observed inhibition of the IKur along with the suppression of the Kv1.5 channel. However, the study does not provide any information about the action of this flavonoid on the atrial action potential. Thus, it remains unclear whether hesperetin can induce the termination of atrial fibrillation. Another study [34][38] investigated the impact of hesperetin on the expression of the Kv1.5 channels in coronary arteries of diabetic and non-diabetic rats. The known fact is that diabetes downregulates the expression of these channels in arterial myocytes. Therefore, it would be beneficial to find a biochemical agent which enhances its expression to the proper level.2.3. Kv2.1 Channel
The voltage-dependent potassium channels Kv2.1 are expressed both in the central and peripheral nervous system of mammals where they are predominant mediators of the delayed rectifier current [35][36][37][69,70,71]. They play a prominent role in shaping neuronal excitability [38][72] and in the glucose-stimulated insulin secretion [39][73], which makes this channel a promising target in treatment of diabetes. In [40][46], the scholars showed an inhibitory impact of genistein on the activation of the Kv2.1 channels. The analysis of the data obtained from the patch-clamp technique on HEK293 cells revealed that in the presence of this compound, the ion channel is inhibited in a concentration-dependent manner. The more profound kinetic analysis showed that genistein shifted the voltage dependence of channels’ activation and inactivation to membrane hyperpolarization. It also accelerated the closed-state inactivation and delayed the recovery from inactivation.2.4. Kv4 Channels
The Kv4 (Shal) channels are widely expressed in the neurons of different animals [41][42][43][74,75,76]. They mediate the fast A-type K+ currents and are thought to be responsible for the fundamental electrical properties of nerve cells. There is not much information about the modulation of these types of ion channels by the flavonoids. In [44][45][77,78], it was shown that pinocembrin upregulates the expression of Kv4.2 channels, which can be beneficial in the treatment of ventricular arrhythmia. A little bit more is known about Kv4.3, which according to [46][48] can be blocked by genistein and, to a smaller extent, by daidzein. The analysis of the Kv4.3 recordings obtained by the patch-clamp technique in the CHO cells revealed that genistein inhibits the current in a reversible and concentration-dependent manner with IC50 ≈ 125 μM.2.5. hERG Channels
The ether-à-go-go-related hERG ion channel (Kv11.1 channel), similarly to the already mentioned Kv1.5 channel, is responsible for the electrical activity of the heart. It regulates the cardiac action potential by mediating the repolarizing current [47][48][79,80]. One of the most important studies concerning the modulation of the hERG channel was performed by Zitron et al. [49][82] in 2005, who screened a large number of flavonoids for their inhibitory abilities. Based on the patch-clamp recordings, they found that the most potent inhibitors of the Kv11.1 channels are naringenin, morin, and hesperetin. Naringenin blocked the hERG channel expressed both in the Xenopus laevis oocytes and HEK 293 cells with half-blocking concentrations IC50 = 102 μM and IC50 = 36.5 μM, respectively. A more detailed analysis revealed that the channels are blocked in the open and inactivated states by naringenin but not in the closed states [50][83]. Moreover, the ECG examinations suggested that the blockade of the hERG channel induced by naringenin results in prolongation of the QT interval [51][45].2.6. Further Kv Channels
The flavonoids also influence the Kv7.1 channel, which contributes to the regulation of the repolarization phase of the cardiac action potential. Puerarin is an isoflavonoid found in the root of Pueraria Lobata, which is known from its anti-inflammatory, immunomodulatory, anti-cancer and cardioprotective properties [52][95]. In [53][43], it was demonstrated that the isoflavone, puerarin, effectively downregulates the channel activity via direct interaction with the channel protein. It was reported that this inhibitory effect (along with the blockade of the slow delayed rectifier current IKS) contributed to the prolongation of action potential duration, which can be beneficial in the case of treatment of cardiovascular diseases. It turns out that also naringenin exerts an inhibitory effect on this channel, with a mild impact of this flavonoid on the current [51]. Eventually, the studies performed by Kang et al. [54] revealed that also (−)-epigallocatechin-3-gallate [54] is a potent inhibitor of the Kv1.7 ion channel.IKS
current [45]. Eventually, the studies performed by Kang et al. [44] revealed that also (−)-epigallocatechin-3-gallate [44] is a potent inhibitor of the Kv1.7 ion channel. Quite recently, it has been discovered that another flavonoid, procyanidin B1, a natural compound extracted from the grape seed, is a potent inhibitor of the Kv10.1 channel (IC50 = 10 μM ), which is overexpressed in some tumors [55][96]. According to this work, targeting the Kv10.1 channel by procyanidin B1 can inhibit the proliferation of cancerous cells. Consequently, this compound is a promising agent for cancer treatment.3. Calcium-Activated Channels (KCa)
3.1. BK Channel
The large-conductance voltage- and Ca2+-activated channels (BK) are ubiquitously expressed K+ channels being characterized by a large single-channel conductance (150–300 pS) [56]. They are considered important drug targets due to their important roles in many physiological processes, such as neural transmission, hearing, endocrine secretion, and smooth muscle contraction [57]. In addition, the mitochondrial BK channel variants (mitoBK) received great scientific interest in terms of the possibilities of their chemical modulation because of the involvement of these channels in the regulation of metabolism, including ATP synthesis as well as the pro-life and pro-death processes [58][59][60].+
channels being characterized by a large single-channel conductance (150–300 pS) [97]. They are considered important drug targets due to their important roles in many physiological processes, such as neural transmission, hearing, endocrine secretion, and smooth muscle contraction [98]. In addition, the mitochondrial BK channel variants (mitoBK) received great scientific interest in terms of the possibilities of their chemical modulation because of the involvement of these channels in the regulation of metabolism, including ATP synthesis as well as the pro-life and pro-death processes [99,100,101]. The impact of flavonoids on the functioning of the BK channels was extensively studied in recent years. The main inferences from those investigations are outlined in Figure 1.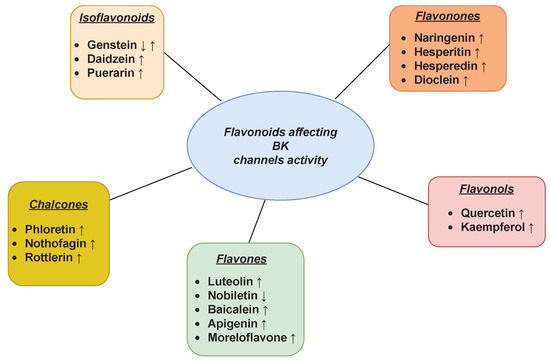
3.2. IK and SK Channels
4. Inward Rectifying Potassium Channels (Kir)
The Inward Rectifying Potassium Channels (Kir) belong to one of the structurally simplest ion channels group containing four identical subunits, each containing two membrane-spanning alpha helices. These channels allow ions to be transported more effectively into than out of the cell. They are responsible for the regulation of resting membrane potential. Thus, their function is mostly related to the modulation of cardiac and neural cells activity, insulin secretion, or epithelial K+ transport [72][155]. The Kir channels are expressed in many cell types: myocytes, neurons, blood cells, endothelial, glial cells, or oocytes [73][156]. The classification of the Kir channels family covers the groups Kir1–Kir7 together with their respective subgroups. Among the Kir channels, one can also distinguish the adenosine triphosphate (ATP)-dependent K+ channels (KATP, Kir6) and the G-protein regulated K+ channels (GIRK, Kir3). The structure of Kir channels lacks a proper voltage-sensing domain. Nevertheless, some representatives of the Kir family exert a bit stronger ‘’voltage dependence” than the others. In that aspect, Kir 2 channels, which are strongly rectifying ones (and consequently more sensitive to extracellular K+), deserve to be distinguished.
The flavonols represented by quercetin and rutin exert significant impact on Kir channels. Trezza et al. [74][159] examined the both modulators and 5-hydroxyflavone in the context of their possible impact on the ATP-sensitive Kir6.1 channel. They compared the experimental results of channels from Rat norvegicus aorta cells with molecular dynamics and docking calculations. All the compared results suggested that there was no effect on Kir6.1 caused by rutin, and significant downregulation in the case of quercetin and 5-hydroxyflavone, but only in the case of the closed channel conformation. The cardioprotective effect of flavonoids on rat myocytes through the regulation of mitochondrial ATP-sensitive potassium channels activity was also shown recently by Rameshrad et al. [75][160]. This research group studied a flavonol, morin, and postulated that its antioxidative effects are mediated by mitochondrial ATP-dependent potassium channels. The activity of Kir6.1 can be upregulated in the presence of isovitexin, which is obtained from the extract of Luehea divaricata Mart., according to [76][154]. This effect supports the regulation of mesenteric arteriolar tone.
Well-pronounced effects on the Kir channels (especially KATP channels) are also documented for a natural alkaloid, berberine (BBR), which is conditionally classified as an ’isoquinoline flavonoid’. BBR is frequently used in the Chinese and East Asian medicines [77][140]. Hua et al. confirmed the inhibition effect of BBR on ATP-sensitive channels [78][167]. The scholars postulated that the anti-arrhythmic and antidiabetic properties of berberine are related to the inhibition of potassium channels. The inhibitory effects of berberine were also investigated by Wang et al. [79][168]. The scholars characterized a similar anti-arrhythmic impact of BBR manifested by the reduction of action potential duration and the effective refractory period of ischemia. In contrast to BBR, a flavonoid from the anthocyanins-cyanidin caused the upreguation of Kir6.2 genes, which have potential implication in glucose sensitivity and its homeostasis [80][169].
Another isoflavonoid, puerarin, exerts an activating effect on the mitochondrial K+ ATP-regulated channels (mitoKATP), according to the results presented in [81][135]. That study concluded that the mitoKATP channel activation participated in the cardioprotection by puerarin. The activation of mitochondrial KATP channels also plays a crucial role in shaping the cardioprotective effects exerted by other flavonoids [82][175]. Among them, six natural compounds should be mentioned: (−)-epigallocatechin-3-gallate [83][176], theaflavin [84][177], proanthocyanidins [85][178], genistein [86][179], baicalein [87][180], and morin[75][160].
5. Two-Pore Domain Potassium Channels (K2P)
The two-pore domain potassium channels are widely distributed in excitable and non-excitable cells and are responsible for the background potassium conductance [88][89][185,186]. They are emerging drug targets in case of a.o. cardiovascular and neurological diseases [90][91][92][93][187,188,189,190].
Among the TREK subgroup of K2P channels, TREK-1 ( K2p2.1) and TRAAK (K2p4.1) are mainly expressed in the central nervous system (CNS), and TREK-2 ( K2p10.1) is expressed in both CNS and peripheral tissues [94][95][191,192]. TREK channels are activated by several stimuli, including biomolecules (e.g., riluzole, nitrous oxide, polyunsaturated fatty acids, and lysophospholipids). These modulators can contribute to the opening of TREKs under pathological conditions. Considering the effects of flavonoids’ administration, the neuroprotective properties of quercetin were demonstrated in [96][193].
6. Conclusions
Flavonoids are widely known for their beneficial health effects, which involve complex biochemical interactions with specific molecular targets, including potassium channels. Due to the fact that these transport proteins play important roles in shaping cardiac action potential as well as smooth muscle tone, their stimulation by flavonoids yields vasorelaxant and cardioprotective effects [63][82][97][98][99][100][81,148,175,198,199,200]. Nevertheless, these effects are not the only examples of the K+ channel-mediated physiological processes that are regulated by flavonoids, as presented in Figure 2.
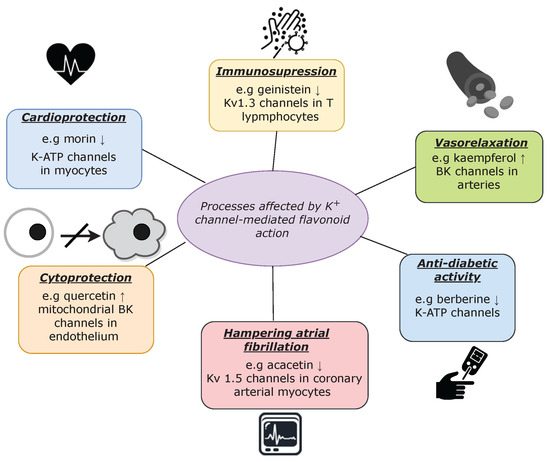
channel modulation (with appropriate examples). The ↑ represents channels’ activation and ↓ denotes channels’ inhibition.