1. Chemical Cue Interaction Leads to Plasticity in Host Plant Recognition
Olfactory cues indicating host plants, similar to cues used for intraspecific communication, do not occur alone in a natural environment but within a background of multiple odors emitted from biotic and abiotic sources
[1][21]. Concerning plant volatile emissions, different mechanisms of interaction for host- and non-host volatiles have been discovered. Non-host volatiles can mask host volatiles without being repellent themselves or alternatively have a repulsive effect, which can reduce or override the attractive effect of host plant volatiles. Thus, insects must integrate olfactory information on attractive and repulsive volatiles and weigh opposing inputs to take an adaptive behavioral decision. Non-host plant volatiles can also indirectly affect host plant recognition by influencing volatile production in the host plant.
An influence of non-host plant volatiles on the emission of volatiles by host plants has been observed in some cases. An example for such interactions has been discovered in host plants of the Asian citrus psyllid
An influence of non-host plant volatiles on the emission of volatiles by host plants has been observed in some cases. An example for such interactions has been discovered in host plants of the Asian citrus psyllid
D. citri. Citrus host plants exposed to guava plants or to guava shoot odors for at least two hours were less attractive to the citrus psyllid than un-exposed plants [7]. Several studies have also examined the effect of companion plants on the attraction of aphids to their host plants. Certain weeds, such as . Citrus host plants exposed to guava plants or to guava shoot odors for at least two hours were less attractive to the citrus psyllid than un-exposed plants [28]. Several studies have also examined the effect of companion plants on the attraction of aphids to their host plants. Certain weeds, such as , reduce the acceptance of barley by
Rhopalosiphum padi [8]. Volatiles of flowering Marigold ( [29]. Volatiles of flowering Marigold ( spp.) also decrease the attractivity of pepper plants (
) for the aphid
2. Chemical Host Plant Recognition Is Modulated by Visual Cues
In many insect species, chemical cues, especially volatiles, seem to be sufficient to localize and identify a suitable host plant. In other insects, attraction to a host plant is strongly enhanced when visual cues are added to olfactory cues. For example, visual and olfactory cues act synergistically in ambrosia beetles during localization of their conifer host trees
[10][33]. Synergistic attraction of visual cues (i.e., shapes and colors) combined with host plant odors have also been found in female melon flies,
Bactrocera cucurbitae [11][34]. Highly plastic interactions between colors and volatiles playing a role in the attraction of
Drosophila suzukii have also been found recently. Depending on the volatiles or volatile blends present, females may choose preferentially different colors, but synergistic effects between odors and colors depend on the origin of the presented odors
[12][35]. In the Asian citrus psyllid,
D. citri, a combination of citrus odors with yellow-colored paper positioned in the corresponding olfactometer fields increased attractivity
[13][36]. In the same species, visual cues, i.e., a green LED to simulate a green plant, enhanced attraction to volatile cues
[14][37].
3. Experience-Dependent Plasticity in Host Plant Responses
3.1. Plasticity Due to Olfactory Experience
Experience has been suggested to modulate host plant choice in various ways, especially in generalist species. Larval odor experience during feeding can influence adult host choice, but adult experience during hatching, early adult life, successful mating on a host plant, and egg-laying may also modify subsequent host choice in insects. Several studies described these phenomena at the behavioral level and investigated underlying mechanisms. In the noctuid moth
Spodoptera littoralis for example, both larval experience during feeding at different developmental stages on a plant, early adult experience, and mating on a specific host plant influence the preferences in the choice of an oviposition site in males and females
[15][16][17][18][41,42,43,44].
Odor learning in larvae is often only possible during a specific sensitive period. In
S. littoralis for example, host plant preference is modified by experience during late larval stages, but not when early larval stages feed on a plant and are subsequently fed on an artificial diet
[19][45]. The hypothesis that experience might also influence behavioral choice in the following generation has also been tested in this species. However, even if larval development was improved when rearing offspring on a matching high-quality host plant, no trans-generational behavioral effects on host choice in young larvae and on female oviposition preferences were found
[20][46]. In another moth species, the European grapevine moth
L. botrana, larval odor experience during feeding on different grapevine cultivars led to female oviposition preference for the experienced cultivar, whereas early adult experience during hatching did not induce any preference
[21][47].
3.2. Learning of Odors through Association with Gustatory Signals
Odors experienced during feeding both by larvae and adults can influence subsequent orientation behavior. This is the case in the citrus psyllid
D. citri, where adults learn to associate certain volatile compounds, such as limonene and benzaldehyde (and also colors), with a sucrose feeding reward and will more likely probe a surface in presence of the conditioned stimulus. Psyllids were able to associate blue color, which is not attractive to unconditioned psyllids, with a food reward when it was scented with citrus odors
[22][52]. In the same species, experience with another compound, eugenol, leads to an aversive response in subsequent behavioral tests
[22][52].
Feeding experience also influences odor response in locusts: starved individuals learn to associate odors (and also visual cues) with high reward food sources in a Y-maze experiment
[23][53]. Odor learning can also be used to avoid ingesting toxic food in this species
[24][54]. Food quality can play a role in learning effects. This is the case for example in
S. littoralis where larval exposure to host plant volatiles influences larval host choice and oviposition preferences in adult females. A poor diet proposed with the host plant volatiles leads to a decrease in attraction of the associated plant, whereas association with a high-quality diet leads to increased attractivity of the corresponding plant
[25][55].
3.3. Plasticity in Contact Compound Responses through Gustatory Experience
Feeding experience with different types of taste compounds has been shown to influence subsequent plant acceptation and feeding behavior. Larvae of different Lepidoptera change their food acceptance behavior after experience with host plants containing antifeedants. In larvae of
Pieris rapae, rearing on a plant containing antifeedants leads to a lack of deterrence in response to different deterrent compounds (strychnine, naringin, and chlorogenic acid) present if larvae are reared on cabbage. At the same time, taste neurons on the maxillary palps reduce their sensitivity to these compounds, thus desensitization of neurons could be at the origin of behavioral desensitization
[26][59]. Experience-dependent adaptation of gustatory neuron sensitivity also occurs for feeding stimulants. In larvae of
Helicoverpa armigera, rearing on high-concentration sucrose diet leads to a transgenerational desensitization of sucrose-sensitive neurons. However, introducing larvae of any subsequent generation to a low-sucrose diet establishes re-sensitization very rapidly within the same generation
[27][62]. Another example of experience-dependent changes in response to gustatory signals is the cockroach adaptation to experience with toxic baits using sucrose as a phagostimulant. The occurrence of sucrose-averse cockroaches seems to be mediated through mutations in gustatory bitter receptors, which respond to glucose, and this stimulation suppresses activation of sucrose (sweet) receptors (activated in both wild type and mutated cockroaches)
[28][64].
4. Influence of Physiological State on Host Plant Responses
Behavioral responses to host-plants or food sources depend on a variety of parameters, including feeding state, mating state, age, period within reproductive cycle, sex, and morph for polymorphic species. The changes in these state-dependent responses very often have similar underlying mechanisms reaching from state-dependent variations in gene-expression at the peripheral and central nervous level, over receptor neuron and central neuron function, as well as neuromodulator action, up to anatomical changes in peripheral and central chemosensory structures.
4.1. Nutritional State and Symbiotic Bacteria can Modify Responses to Food- and Host-Related Volatiles
Whereas effects of the nutritional state on host-seeking behavior and underlying mechanisms have been broadly investigated in blood-feeding insects (
[29][66] and references therein) and parasitoids
[30][67], very little is known about such effects in herbivorous insects. However, decisions between the search for food sources or oviposition sites depend on previous food intake history in female herbivores as well. Nutritional state-dependent variations in behavior and its underlying neural mechanisms have been studied extensively in
D. melanogaster (for review see
[31][68]). The feeding state can influence foraging behavior directly but is also dependent on gut bacteria. In
B. dorsalis, foraging in protein-deprived flies is more goal-oriented in aposymbiotic flies than in flies with intact intestinal bacteria
[32][70]. Similarly, olfactory preferences for cues emitted by food sources and associated microbes depends on gut microbiota in
D. melanogaster [33][71]. However, facultative symbionts in several highly specialized biotypes of the pea aphid,
Acyrthosiphon pisum, seem not to influence host plant selection
[34][72].
4.2. Pronounced Effects of Age and Mating State on Female Responses to Host-Related Volatiles
In most insects, attraction to host- or food-related volatiles depends on mating state and age. In female herbivores, egg maturation and egg production rate can also modulate attraction to host plant volatiles, often with an initial increase during early adult life and, in certain long-living insects, a decrease with advanced age. For example, females of the Queensland fruit fly,
Bactrocera tryoni, were highly attracted to guava fruit juice in early adult life, but attraction declined with advanced adult age. However, this decline was correlated with a decline in locomotor activity (potentially due to decreasing oviposition rates) and once insects started to move, they still oriented towards the fruit odor
[35][76]. In another study, mated females of the same species were more strongly attracted by fruit stimuli than virgin females and mated females spent more time on a fruit when reaching it
[36][77]. In different psyllids, host choice is more selective in mated individuals. In the citrus psyllid
D. citri, host plant volatiles attracted only mated males and females
[14][37]. In the carrot psyllid,
Trioza apicalis, host plant volatiles were overriding light intensity cues in mated females when competing visual and olfactory host cues were tested, whereas virgin females chose volatiles from a non-host plant in strong light conditions over a host plant under weak light
[37][78].
4.3. Sex-Dependent Responses to Host Plant Volatiles
Various differences in host plant volatile responses have been observed between male and female insects depending on their biology. In herbivorous insects where adult males do not feed, host plant volatiles alone often do not attract males as in the grapevine moth
L. botrana [38][82]. In some cases, such as
Trichoplusia ni, host plant volatiles can attract males on their own, however
[39][83]. More commonly, host plant odors act on males in synergism with pheromones as an additional cue to localize females (reviewed in
[40][84]). In several moth species, such as
Cydia pomonella and
G. molesta, host plant volatiles synergize responses to female-emitted sex pheromones
[41][42][85,86]. In male
Helicoverpa zea, host plant volatiles were even found to synergize responses to the sex pheromone in pheromone-specific olfactory receptor neurons
[43][87]. Cases of synergistic attraction of host plant volatiles with the female produced sex pheromone were also reported in the scarab beetle
Holotrichia parallela [44][88]. In aphids, such as
Sitobion avenae and
S. fragariae, winged females under autumn conditions are strongly attracted by their respective host plant odors, whereas males in these two species do not respond to host plant odors, but only to the female-emitted sex pheromone. In another aphid species,
R. padi, however, even males respond to the odor of their primary host (reviewed in
[45][90]).
4.4. Morph-Dependent Responses to Host Plant-Related Volatiles
Different morphs of the same insect (with identical genomes) can also vary in behavioral and physiological responses to host plant volatiles. In certain aphid species, high population densities lead to the formation of winged morphs, to facilitate dispersal
[46][91]. In addition to the increased number of olfactory organs on the antennae, evidenced in winged morphs of different species
[47][48][92,93], stronger preferences to host plant odors have also been identified, for example in
M. persicae [49][30]. In the same aphid species, responses to the host plant potato were shown to be influenced by volatile emissions caused by a companion plant (onion) but the level of response was morph dependent: winged aphids were repelled by high doses of onion-induced terpenoids (nerolidiol, TMTT) emitted from potato, whereas wingless morphs were only repelled by low doses of the same compounds
[49][30]. To
aou
thor'sr knowledge no studies comparing responses to contact cues have been conducted between morphs so far.
Similar to aphids, locusts also occur in two morphs depending on population density. They switch from solitary to gregarious forms, which differ considerably in their sensory equipment and responses to host plant volatiles. Solitary locusts possess higher numbers of antennal chemoreceptors
[50][94] and different expression of antennal odorant binding proteins compared to gregarious locusts
[51][95]. Larger numbers of receptor neuron axons converge within the primary olfactory centers in solitary locusts, leading to larger relative sizes of the antennal lobes compared to gregarious locusts
[52][53][96,97]. Central olfactory neurons respond more frequently to plant volatiles, corresponding to a higher sensitivity and localization over larger distances of host plants, as well as more selectivity in host plant choice
[54][55][98,99].
5. Implications of chemosensory plasticity for alternative pest management
Efforts to replace classical pest management strategies using synthetic insecticides with alternative strategies have been made since many years. However, recent developments in the banning of hazardous insecticides require more research on alternative pest management strategies, and chemical ecology provides promising avenues for behavior-based control strategies using plant compounds. Such methods comprise the use of repulsive and attractive compounds in dispensers and/or companion plants and possibly their essential oils, either in combination with confined insecticides in an attract and kill approach, or the combination of repulsive and attractive elements in push–pull systems. Other approaches include masking of host plant compounds to disrupt recognition, selection of crop varieties emitting less attractive volatiles, and manipulation of volatile emission in plants by plant resistance induction through plant defense stimulators [56][57][108,109].
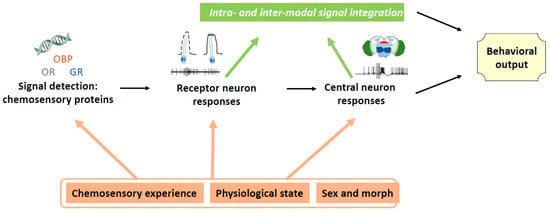
Chemical signals, alone or in concert with other sensory cues, are essential for host plant recognition and choice, but determining the identity of attractive or repulsive compounds is far from being sufficient to develop efficient and reliable crop protection methods based on these signals. First, collection of potentially active volatiles needs to be performed at the correct physiological state of the host or non-host plant. Second, the complexity of the signal and interactions between various host plant cues need to be considered. Indeed, as shown in the present review, there are multiple interaction levels between various host plant cues and their environment (Figure 1). The knowledge on chemical cue interactions is important for potential applications, because the use of attractive or repulsive plants is costly both in time and money, and therefore, as a first step, identification of individual volatile compounds, which can potentially be produced at a reasonable cost and applied relatively easily is often attempted. Nevertheless, it is difficult to reach high efficiency with this approach.

Figure 1.
Overview over the different origins of plasticity in host plant recognition.
Furthermore, as shown in the present review, insect behavior is highly plastic (Figure 1) and therefore the different forms of plasticity need to be taken into account when developing pest control methods involving the chemical senses. Insects with the same genome might not respond in the same way to chemical stimuli as a function of various factors, such as experience, physiological state, and season of the year. Powerful attractants or repellents might change their efficiency through adaptation and learning processes, as shown for the above-described case of a switch in the effect of sucrose from a phagostimulant to an aversive substance in cockroaches experienced with toxic baits [58][64]. Attractive, repellent, or stimulating compounds might not have the same effects on mature and immature insects.
HeThis re, itview shows that mechanisms of insect host plant localization and recognition are highly plastic. These mechanisms are an excellent target for the development of alternative pest control methods. Some of these are already widely used but have a significant potential to be further improved and extended to a large range of pests. In particular, their use may have been oversimplified, while it is showedwe show here that plasticity both among plant cues or in behavioral responses of insects may strongly influence their efficiency and durability of use. Based on the rich literature reviewed herein, there is a in the present paper, we hope to see consideration of plasticity in future research on next-generation, sustainable, pest insect control based on behavioral manipulation.