Aflatoxins are natural toxicants produced mainly by species of the Aspergillus genus, which contaminate virtually all feeds and foods. Apart from their deleterious health effects on humans and animals, they can be secreted unmodified or carried over into the milk of lactating females, thereby posing health risks to suckling babies. Aflatoxin M1 (AFM1) is the major and most toxic aflatoxin type after aflatoxin B1 (AFB1). It contaminates human breast milk upon direct ingestion from dairy products or by carry-over from the parent molecule (AFB1), which is hydroxylated in the liver and possibly in the mammary glands by cytochrome oxidase enzymes and then excreted into breast milk as AFM1 during lactation via the mammary alveolar epithelial cells.
- human breast milk
- aflatoxin M1
- child growth and development
- health risk
- influx/efflux transport
1. Routes of Breast Milk Contamination with AFM1
1.1. Diet
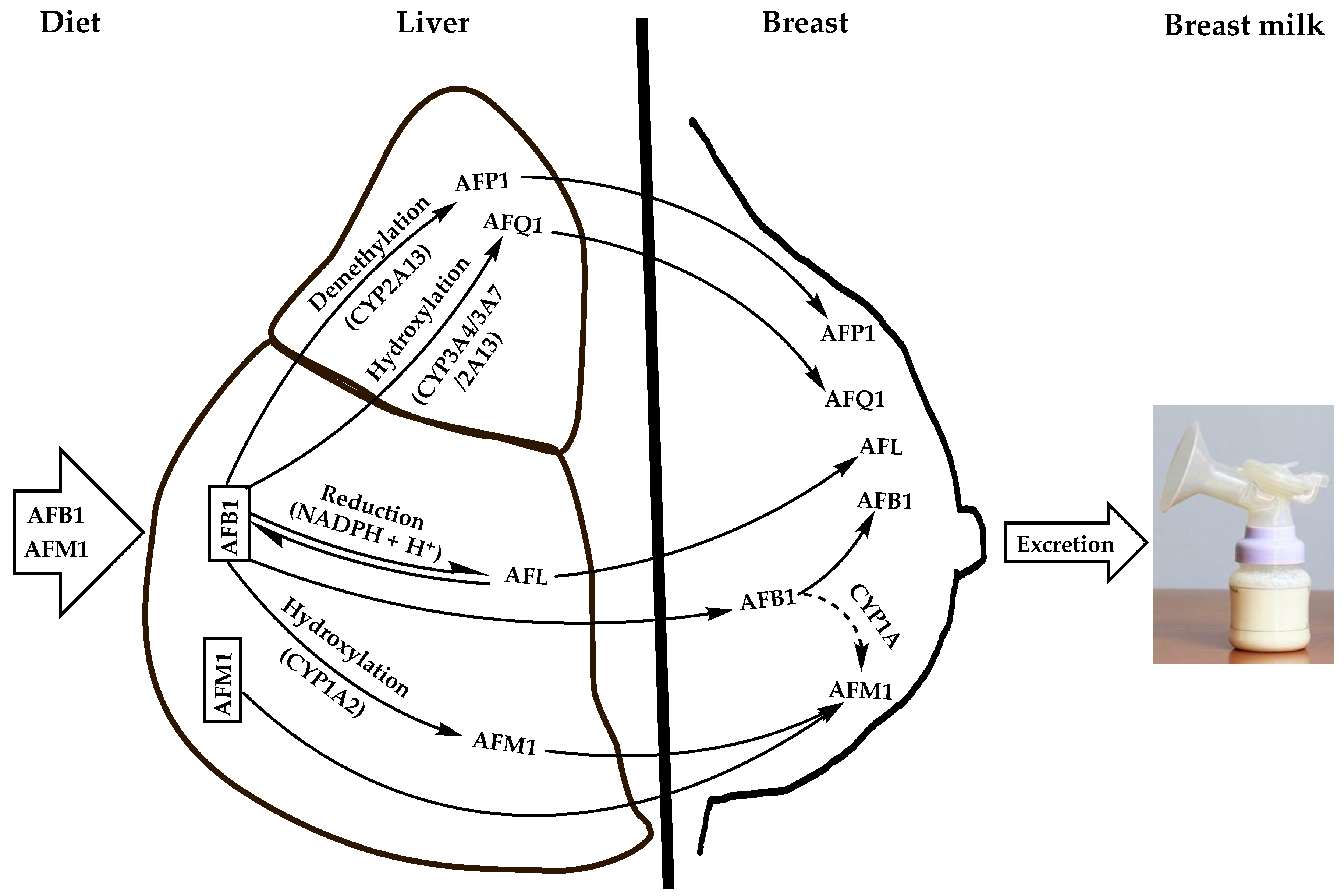
1.2. Feed as an Indirect Source of Human Breast Milk Contamination with AFM1
2. Adverse Health Effects of AFM1 on Infants and Young Children
Infants can be exposed to AFM1 from their in utero life throughout the nursing period where breastfeeding can be the main or exclusive food source [47][48][49]. Under these conditions, the extent of exposure is highly dependent on AFM1 intake by the mother as well as the rate of its transfer to the baby through the umbilical cord, as a fetus, and then through breast milk, as a suckling newborn and infant. Such continuous exposure can cause teratogenicity, stillbirth, or miscarriage during pregnancy [49][50] or lead to physiological and neurological disorders that the child would suffer the consequences for the rest of his/her life, such as stunting, malnutrition diseases (e.g., kwashiorkor and marasmus), autism, nodding syndrome, and related cognitive disorders [51]. The highest incidence of these diseases is recorded in low-income and middle-income countries of the endemic regions, especially those of Asia and Africa under subtropical climate conditions [32][47][52]. Breast milk contamination with AFM1 is of serious concern to public health for three main reasons: (i) the potential high intake of AFM1 by neonates and infants who are fed mainly or exclusively on breast milk in case of a high contamination; (ii) the weaker detoxifying capacity due to their immature organs, mainly the liver, and higher metabolic activity; and (iii) the possible continued exposure after infancy and childhood to aflatoxins through various foods [53][54], which increases the risk for the onset of severe chronic endpoints, such as cancer, at younger ages. These conditions result in an overall increase in the susceptibility of infants and young children to aflatoxins by about three-fold more than adults [53][55]. Therefore, high exposure of pregnant and nursing women to AFM1 can be anticipated to increase the rates of disabilities, morbidities, and mortalities within a society resulting in a heavy economic and social burden.2.1. Growth Impairment
Stunting, underweight, or wasting of children are the most documented growth impairments associated with the exposure to AFM1 during fetal life and infancy through umbilical cord blood and breast milk, respectively [50][56][57]. In the year 2020, 149.2 million (22%) and 45 million (6.7%) of the world children under 5 of age were affected by stunting (low height for age ratio) and wasting (low weight for height ratio), respectively [58]. In the sub-Saharan countries of Africa, notorious for the incidence of aflatoxins in their foods and feeds [59], the prevalence of stunting and underweight in children below 5 years of age were reported to be as high as 38% and 22%, respectively in 2015/2016 [60]. Although aflatoxin contamination is not the only etiology that would explain such a high incidence of growth impairment, its contribution cannot be overlooked. From the public health standpoint, these growth disorders have life threatening or life lasting consequences on affected children. Stunting alone was estimated to cause the death of children by 14–17% and was considered to be an underlaying cause of poor cognitive, motor development, and educational performances; it was even suggested to be congenitally transmitted to the offspring [33][58]. Wasting, on the other hand, can be treated and weight gain can resume normally, but it increases the death risk depending on the severity or leads to stunting upon prolonged exposure or after recurrent episodes [47][58]. Despite the prevailing belief that under-nutrition, inadequate dietary intake, and gastrointestinal illness are the primary etiologies of growth impairment [61], it is now well established that exposure to mycotoxins, e.g., aflatoxins and fumonisins, in utero and throughout infancy and early childhood hinders the linear growth and weight gain [33][47][49][53][56][57][62][63][64]. This was further corroborated by the failure of interventions, such as the provision of appropriate education on nutrition and complementary feeding in addition to the implementation of proper water, sanitation, and hygiene (WASH) to improve significantly the linear growth and normal weight gain in undernourished children [65][66][67][68][69][70][71]. One of the earliest studies that demonstrated the causal relationship between growth impairment and exposure of children to aflatoxins was conducted in Benin and Togo of Western Africa [62]. Herein, the Z scores of the height-for-age (HAZ), weight-for-age (WAZ), and height-for-weight (HWZ) were determined in children below 5 years of age whose diet consisted of breast milk only (exclusive breast feeding), breast milk and weaning foods, breast milk and household foods, and weaning and household foods. When matched with aflatoxin-albumin (AF-Alb) adduct levels in the blood, the outcome revealed that the levels of AF-Alb were inversely related to HAZ and WAZ scores, i.e., the higher these levels were, the more severe the stunting and the underweight occurred in children. A subsequent longitudinal study on Gambian children showed that high exposure to aflatoxins from the perinatal period to the age of one year, as evidenced by high levels of AF-Alb adducts in the maternal, umbilical cord, and children blood, reduced significantly the height and weight gains in children from 6 months to one year of age [57]. Conversely, it is demonstrated that a reduction of AF-Alb levels in the maternal blood from 110 pg/mg to 10 pg/mg during pregnancy resulted in an increased height and weight gains of 0.8 kg and 2 cm, respectively by the children during the first year of growth. According to the authors' understanding, this effect stems from the maternal exposure to aflatoxins during pregnancy and extends throughout the first year of the children’s life, which has established the association between aflatoxin exposure during infancy and growth impairment. However, in the first 16 weeks after birth where the children were essentially breastfed, the serum AF-Alb was detected at low levels in only few infants (13 out of 115 babies: 11%), suggesting that were residual aflatoxin adducts from the maternal exposure [57]. Moreover, no direct causal effect between the growth impairment and the presence of AFM1 in the breast milk of the surveyed mothers was specifically demonstrated here, as the exposure to aflatoxins was measured indirectly by the blood levels of AF-Alb, which typically evokes AFB1 exposure [72]. However, since AFM1 can form albumin adducts, albeit at lower extents than does AFB1, AF-Alb can also indicate exposure to AFM1 [73]. In addition, AF-Alb levels in the children’s blood were shown to correlate highly with AFM1 levels in their urine [74], suggesting that the urinary AFM1 most likely originates from the diet (mother’s milk and/or complementary food) or from the maternal and cord blood in the case of neonates [50] rather than being an AFB1 metabolite due to the low AFB1-detoxifying activity in the young children [75]. The inverse relationship between AFM1 in breast milk and growth impairment in nursing children has been reported in different countries around the world. A study on lactating women from Tehran (Iran) showed that breast milk contamination at levels varying between 0.3 and 26.7 ng/kg (median of 8.2 ng/kg) was inversely correlated with height at birth but not with weight [76]. Another study conducted in the same country on exclusively breastfed children of 90–120 days old from the city of Tabriz showed that their exposure to an average AFM1-concentration of 6.96 ng/L (range of 5.1 to 8.1 ng/L) retarded both height and weight gain compared with children fed on AFM1-free breast milk [77]. The growth impairment of suckling children despite the generally low levels of AFM1 in breast milk was attributed to a chronic exposure starting in utero from the carry-over of AFB1 and/or AFM1 from the mother’s diet, as substantiated by the under-height at birth [76], and continuing exposure after birth through the maternal milk and/or weaning and household foods [47][76][78]. The association of exposure to AFM1 from mothers’ milk with growth impairment in children from fetal to early life is well documented [62][76][77][79][80]. However, most of the studies were observational (cohort or cross-sectional) with less power as evidence and each of them has its own pitfalls and limitations. To the best, they may make the causal link circumstantial rather than direct. In addition, no study has demonstrated the mechanism of action to elucidate how do aflatoxins, including AFM1, act to cause stunting, weight gain, or wasting in children. The demonstration of the mechanism of action is required by the IARC to consider the causal link being direct [81]. Therefore, there is a need for specifically designed studies to provide unequivocal clinical evidence for the association between AFM1 exposure and growth impairment. Longitudinal studies using cluster randomized controlled clinical trials on children intentionally given low doses of AFM1 appear to be best fit for such a purpose, but they are challenged with the dilemma of the ethical considerations for the use of human subjects. Prior to performing these trials, the experimental design should be described in detail and submitted for review and approval to national and international organizations concerned by the ethical questions of scientific research. Once approved, the research team should be committed to provide updates on a regular basis and to communicate to the authorizing body any amendments introduced into the protocol in due course. This procedure is intended to ensure that the study meets strictly the ethical principles, e.g., maintaining a favorable balance of risk/benefit and respecting participants among other provisions, while making a significant contribution to the advance of the scientific knowledge in the field. So far, only two studies have been done in this framework, and they have used AFB1 [71] and total aflatoxins [32] on partially breastfed children. None of them has specifically investigated the effect of AFM1 on breastfed children. The first of these studies was conducted in Kenya on infants who have been recruited before birth (starting from the fifth month of pregnancy) and followed until the age of 22 months for linear growth and the concentrations of serum AFB1-Alb [71]. Recruited mothers and their children were split into two cluster-randomized groups consisting of an intervention group, receiving aflatoxin-safe maize (<10 ng/g of AFB1) as complementary food, and a control group receiving a regular household maize known to be usually contaminated with higher levels of AFB1. The clinical trials of study were approved in 2013 by Institutional Review Board for Research of the International Food Policy Research Institute (https://www.socialscienceregistry.org/trials/105 (accessed on 10 June 2022)) and ended in 2016 revealing the absence of causal link between AFB1 intake and stunting at 22 months of age [71]. These results should be interpreted with caution, as the study suffered many limitations, the most prominent of which was the high rate of follow-up loss and incomplete data collection in both the intervention and the control groups. Additionally, at the midline of the experiment (13 months of age), a-7% decrease in the stunting rate was observed but the exposure (serum AF-Alb) did not decrease. Conversely, at the endline (22 months of age), no improvement in linear growth was observed despite a significant decrease in serum AF-Alb. These results, which remain unexplained, suggest that interfering factors not considered in the design of the study, such as seasonal variation, environmental enteric dysfunction, immunomodulation, and hepatic metabolism of micronutrients, may have affected its outcome. The second study of the kind (Cluster randomized controlled trial) being conducted on Tanzanian children was approved for clinical trials in 2019 by the Institutional Review Board (IRB) and the Tanzanian National Institute for Medical Research (NIMR) (https://www.clinicaltrials.gov/ct2/show/NCT03940547 (accessed on 12 June 2022)). Herein, children were recruited at birth and followed for linear growth and serum AF-Alb for one year, including 6 months of exclusive breastfeeding followed by mixed feeding (breastfeeding was not interrupted). Recruited children were split into two-group clusters: the intervention group were intentionally exposed to low doses of total aflatoxins (up to 5 ng/g) after the sixth month of age via complementary food on a continuous basis for 18 additional months. The research team has published yearly updates in scientific journals [32][33][34] and in the ClinicalTrials.gov website [82]; the final outcome is as yet to be disclosed. Although the latter study did not address AFM1 in breast milk, it may serve as a model for future studies on exclusively breastfed children in areas where nursing mothers are fed on staple foods highly contaminated with aflatoxins, hence likely to secrete AFM1-contaminated breast milk for the control cluster group. The intervention cluster randomized group should comprise mothers fed on aflatoxin-safe foods and their breast milk be tested for the absence or safe levels of AFM1.2.2. Other AFM1-Related Health Issues
Several adverse health effects of aflatoxins on infants and young children have been proposed as standalone diseases or as possible underlying mechanisms of action for growth impairment. These include immunomodulation causing chronic immune activation; gastrointestinal diseases; nutrient maldigestion and malabsorption; and impaired bone growth and remodeling [83][84][85][86]. The common feature to all these diseases is that they are related to the disruption of the small intestine functions mediated by damaging its lining epithelium. Additionally, the pathological and clinical features associated with aflatoxin intoxications have been attributed to a sub-clinical condition known as environmental enteric dysfunction (EED) being an underlying cause of stunting and anemia [84][86][87]. Although EED is primarily associated with the ingestion of high load of fecal bacteria under poor water, sanitation, and hygiene conditions that characterize developing countries [88][89], it was reported to share overlapping pathways with aflatoxin-mediated diseases [85]. For example, like EED, aflatoxins were hypothesized to impair protein synthesis thereby promoting gastrointestinal infections and liver toxicity, ultimately leading to growth impairment [84][90]. Few and fragmentary studies have been done on the immunomodulatory effects of aflatoxins on children and their impact on liver toxicity and protein synthesis. Turner et al. [83] first reported that a high exposure of Gambian children to aflatoxins suppresses selectively their humoral immunity. A drastic reduction in salivary IgA titers of these children with a concomitant increase in the levels of serum AF-Lys biomarkers was observed. It is concluded that a high exposure of children to aflatoxins compromises their immunity, which explains the frequency of their gastrointestinal infections and hence the burden of infant infection-related mortality in West African countries. However, the same study showed that the levels of serum AF-Lys did not correlate with the Cell Mediated Immunity (CMI) response of to test antigens (tetanus, diphtheria, Streptococcus, tuberculin, candida, Trichophyton, and Proteus), nor did it with the antibody response to rabies vaccines. Further studies are thus needed to substantiate the relationship between exposure of young children to aflatoxins and the immunomodulatory effects on one hand and to gastrointestinal infection-related mortality on the other hand. A recent longitudinal study investigated the hypothesis that exposure to aflatoxins impairs protein synthesis in children with an emphasis on the proteins used as biomarkers of inflammatory reactions (C-reactive protein, α-1-glycoprotein) as well as other serum proteins directly or indirectly involved in growth development, such as transthyretin, lysine, tryptophan, and Insulin-like growth factor-1 (IGF-1) [85]. The study was conducted on 102 Ethiopian children (6–35 months of age), including 50 stunted children, living in an aeras with highly contaminated staple foods (>10 mg/g). The study aiming to relate chronic exposure to aflatoxins (AFB1, AFB2, AFG1, AFG2, and AFM1) to linear growth impairment showed no clear correlation between exposure to aflatoxins, separately or in combination, to the protein status, inflammation, or linear growth. Exposure of children to aflatoxins has also been suggested as a possible etiology for the onset or the aggravation of the clinical manifestations of neurological disorders, such as autism and nodding. A cross-sectional study conducted in Italy on autistic children (n = 172) revealed a significant difference between the levels of AFM1 in their blood compared with control group of non-autistic children or those at a risk (genetic relatedness to autistic parents). Nevertheless, other studies suggested that, like EED, the neurological disorders associated with exposure to aflatoxins relate to intestinal lining damage and to microbiota–gut–brain axis imbalance (dysbiosis) [91]. This microbiota is known to play a central role in the regulation of the metabolism and homeostasis as well as in controlling the CNS functions via neural, endocrine, and immune pathways. Therefore, its disturbance causes inflammatory bowel diseases and systemic inflammation leading to the alteration of the central nervous system (CNS) functions as is the case in some neuropsychiatric conditions, including autism [92]. Malnutrition-related diseases, such as kwashiorkor, marasmic kwashiorkor and marasmus, faltering, stunting, nodding, organomegaly, and retarded mental and physical activities have been ascribed to chronic exposure of children below 5 years of age to aflatoxins since their in utero life [47][93][94][95][96][97]. However, the exact relationship between the exposure of children to aflatoxins and the occurrence of these diseases as well as the specific contribution of breast milk AFM1 to their onset has been poorly investigated and remain to be substantiated. The occurrence of AFM1 in breast milk to different extents depending on the countries and the agroclimatic zones within the same country as well as the socio-economic conditions is well documented [36][98][99]. Due to the well-established toxicity of AFM1 in humans and animals [47], its occurrence in breast milk is of serious concern to the public health, as it affects individuals in their early life causing either immediate or delayed death or inducing lifetime disabilities. Therefore, there is an urgent need to address this issue at national level and globally regardless of the paucity of scientific evidence for a direct causal link between exposure to this toxicant and the claimed health effects it may cause to children in utero, during infancy, childhood, and even at later stages of their lives.References
- Deng, J.; Zhao, L.; Zhang, N.Y.; Karrow, N.A.; Krumm, C.S.; Qi, D.S.; Sun, L.H. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res. 2018, 778, 79–89.
- Bertero, A.; Augustyniak, J.; Buzanska, L.; Caloni, F. Species-specific models in toxicology: In vitro epithelial barriers. Environ. Toxicol. Pharmacol. 2019, 70, 103203.
- Benkerroum, N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int. J. Environ. Res. Public Health 2020, 17, 423.
- Yiannikouris, A.; Jouany, J.-P. Mycotoxins in feeds and their fate in animals: A review. Anim. Res. 2002, 51, 81–99.
- Ghadiri, S.; Spalenza, V.; Dellafiora, L.; Badino, P.; Barbarossa, A.; Dall’Asta, C.; Nebbia, C.; Girolami, F. Modulation of aflatoxin B1 cytotoxicity and aflatoxin M1 synthesis by natural antioxidants in a bovine mammary epithelial cell line. Toxicol. Vitr. 2019, 57, 174–183.
- Smith, J.W.; Groopman, J.D. Aflatoxins. In Encyclopedia of Cancer, 3rd ed.; Boffetta, P., Hainaut, P., Eds.; Academic Press: Oxford, UK, 2019; pp. 30–43.
- Loveland, P.M.; Wilcox, J.S.; Pawlowski, N.E.; Bailey, G.S. Metabolism and DNA binding of aflatoxicol and aflatoxin B1 in vivo and in isolated hepatocytes from rainbow trout (Salmo gairdneri). Carcinogenesis 1987, 8, 1065–1070.
- Bailey, G.S.; Loveland, P.M.; Pereira, C.; Pierce, D.; Hendricks, J.D.; Groopman, J.D. Quantitative carcinogenesis and dosimetry in rainbow trout for aflatoxin B1 and aflatoxicol, two aflatoxins that form the same DNA adduct. Mutat. Res. 1994, 313, 25–38.
- Carvajal-Moreno, M.; Vargas-Ortiz, M.; Hernández-Camarillo, E.; Ruiz-Velasco, S.; Rojo-Callejas, F. Presence of unreported carcinogens, Aflatoxins and their hydroxylated metabolites, in industrialized Oaxaca cheese from Mexico City. Food Chem. Toxicol. 2019, 124, 128–138.
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7.
- Massey, T.E.; Guindon, K.A. Aflatoxins. In Encyclopedia of Cancer; Schwab, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; p. 101.
- Dohnal, V.; Wu, Q.; Kuča, K. Metabolism of aflatoxins: Key enzymes and interindividual as well as interspecies differences. Arch. Toxicol. 2014, 88, 1635–1644.
- Mykkänen, H.; Zhu, H.; Salminen, E.; Juvonen, R.O.; Ling, W.; Ma, J.; Polychronaki, N.; Kemiläinen, H.; Mykkänen, O.; Salminen, S.; et al. Fecal and urinary excretion of aflatoxin B1 metabolites (AFQ1, AFM1 and AFB-N7-guanine) in young Chinese males. Int. J. Cancer 2005, 115, 879–884.
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological properties and their involvement in cancer development. Toxins 2018, 10, 214.
- Bailey, G.S.; Dashwood, R.; Loveland, P.M.; Pereira, C.; Hendricks, J.D. Molecular dosimetry in fish: Quantitative target organ DNA adduction and hepatocarcinogenicity for four aflatoxins by two exposure routes in rainbow trout. Mutat. Res. 1998, 399, 233–244.
- Karabulut, S.; Paytakov, G.; Leszczynski, J. Reduction of aflatoxin B1 to aflatoxicol: A comprehensive DFT study provides clues to its toxicity. J. Sci. Food Agric. 2014, 94, 3134–3140.
- Caruso, M.; Mariotti, A.; Zizzadoro, C.; Zaghini, A.; Ormas, P.; Altafini, A.; Belloli, C. A clonal cell line (BME-UV1) as a possible model to study bovine mammary epithelial metabolism: Metabolism and cytotoxicity of aflatoxin B1. Toxicon 2009, 53, 400–408.
- IARC. Monographs on the Evaluation of Carcinogenic Risks to Humans: Chemical Agents and Related Occupations; A Review of Human Carcinogens, 100 F; IARC: Lyon, France, 2012.
- Polychronaki, N.; Turner, P.C.; Mykkänen, H.; Gong, Y.Y.; Amra, H.; Abdel-Wahhab, M.; El-Nezami, H. Determinants of aflatoxin M1 in breast milk in a selected group of Egyptian mothers. Food Addit. Contam. 2006, 23, 700–708.
- Carvajal, M.; Rojo, F.; Méndez, I.; Bolaños, A. Aflatoxin B1 and its interconverting metabolite aflatoxicol in milk: The situation in Mexico. Food Addit. Contam. 2003, 20, 1077–1086.
- Warth, B.; Braun, D.; Ezekiel, C.N.; Turner, P.C.; Degen, G.H.; Marko, D. Biomonitoring of Mycotoxins in Human Breast Milk: Current State and Future Perspectives. Chem. Res. Toxicol. 2016, 29, 1087–1097.
- Braun, D.; Abia, W.A.; Šarkanj, B.; Sulyok, M.; Waldhoer, T.; Erber, A.C.; Krska, R.; Turner, P.C.; Marko, D.; Ezekiel, C.N.; et al. Mycotoxin-mixture assessment in mother-infant pairs in Nigeria: From mothers’ meal to infants’ urine. Chemosphere 2022, 287, 132226.
- García-Lino, A.M.; Álvarez-Fernández, I.; Blanco-Paniagua, E.; Merino, G.; Álvarez, A.I. Transporters in the Mammary Gland—Contribution to Presence of Nutrients and Drugs into Milk. Nutrients 2019, 11, 2372.
- Williams, J.A.; Phillips, D.H. Mammary Expression of Xenobiotic Metabolizing Enzymes and Their Potential Role in Breast Cancer. Cancer Res. 2000, 60, 4667–4677.
- Williams, J.A.; Stone, E.M.; Millar, B.C.; Gusterson, B.A.; Grover, P.L.; Phillips, D.H. Determination of the enzymes responsible for activation of the heterocyclic amine 2-amino-3-methylimidazoquinoline in the human breast. Pharmacogenetics 1998, 8, 519–528.
- Kuilman, M.E.M.; Maas, R.F.M.; Fink-Gremmels, J. Cytochrome P450-mediated metabolism and cytotoxicity of aflatoxin B1 in bovine hepatocytes. Toxicol. Vitr. 2000, 14, 321–327.
- Héraud, F.; Barraj, L.M.; Moy, G.G. GEMS/Food Consumption Cluster Diets. In Total Diet Studies; Moy, G.G., Vannoort, R.W., Eds.; Springer: New York, NY, USA, 2013; pp. 427–434.
- Britzi, M.; Friedman, S.; Miron, J.; Solomon, R.; Cuneah, O.; Shimshoni, J.A.; Soback, S.; Ashkenazi, R.; Armer, S.; Shlosberg, A. Carry-over of aflatoxin B1 to aflatoxin M1 in high yielding Israeli cows in mid- and late-lactation. Toxins 2013, 5, 173–183.
- Ayar, A.; Sert, D.; Con, A.H. A study on the occurrence of aflatoxin in raw milk due to feeds. J. Food Safety 2007, 27, 199–207.
- Costamagna, D.; Gaggiotti, M.; Chiericatti, C.A.; Costabel, L.; Audero, G.M.L.; Taverna, M.; Signorini, M.L. Quantification of aflatoxin M1 carry-over rate from feed to soft cheese. Toxicol. Rep. 2019, 6, 782–787.
- Veldman, A.; Meijs, J.A.C.; Borggreve, G.J.; Heeres-van der Tol, J.J. Carry-over of aflatoxin from cows’ food to milk. Anim. Sci. 1992, 55, 163–168.
- Phillips, E.; Turner, P.C.; Ngure, F.M.; Kassim, N.; Makule, E.; Smith, L.E.; Nelson, R.J.; Stoltzfus, R.J. Ethical considerations in the design and conduct of a cluster-randomised mycotoxin mitigation trial in Tanzania. World Mycotoxin J. 2022, 15, 213–222.
- Phillips, E.; Ngure, F.; Smith, L.E.; Makule, E.; Turner, P.C.; Nelson, R.; Kimanya, M.; Stoltzfus, R.; Kassim, N. Protocol for the trial to establish a causal linkage between mycotoxin exposure and child stunting: A cluster randomized trial. BMC Public Health 2020, 20, 598.
- Phillips, E.; Turner, P.C.; Kassim, N.; Makule, E.; Nelson, R.; Ngure, F.; Smith, L.; Stoltzfus, R. Ethical considerations of the trial to establish a causal linkage between mycotoxin exposure and child stunting. Curr. Dev. Nutr. 2021, 5 (Suppl. 2), 678.
- El-Nezami, H.S.; Nicoletti, G.; Neal, G.E.; Donohue, D.C.; Ahokas, J.T. Aflatoxin M1 in human breast milk samples from Victoria, Australia and Thailand. Food Chem. Toxicol. 1995, 33, 173–179.
- Adejumo, O.; Atanda, O.O.; Raiola, A.; Somorin, Y.; Bandyopadhyay, R.; Ritieni, A. Correlation between aflatoxin M1 content of breast milk, dietary exposure to aflatoxin B1 and socioeconomic status of lactating mothers in Ogun State, Nigeria. Food Chem. Toxicol. 2013, 56, 171–177.
- Elaridi, J.; Bassil, M.; Kharma, J.A.; Daou, F.; Hassan, H.F. Analysis of Aflatoxin M1 in Breast Milk and Its Association with Nutritional and Socioeconomic Status of Lactating Mothers in Lebanon. J. Food Prot. 2017, 80, 1737–1741.
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24.
- El-Tras, W.F.; El-Kady, N.N.; Tayel, A.A. Infants exposure to aflatoxin M1 as a novel foodborne zoonosis. Food Chem. Toxicol. 2011, 49, 2816–2819.
- Benkerroum, N. Mycotoxins in dairy products: A review. Int. Dairy J. 2016, 62, 63–75.
- Battacone, G.; Nudda, A.; Palomba, M.; Pascale, M.; Nicolussi, P.; Pulina, G. Transfer of aflatoxin B1 from feed to milk and from milk to curd and whey in dairy sheep fed artificially contaminated concentrates. J. Dairy Sci. 2005, 88, 3063–3069.
- van der Fels-Klerx, H.J.; Camenzuli, L. Effects of Milk Yield, Feed Composition, and Feed Contamination with Aflatoxin B1 on the Aflatoxin M1 Concentration in Dairy Cows’ Milk Investigated Using Monte Carlo Simulation Modelling. Toxins 2016, 8, 290.
- Frobish, R.A.; Bradley, B.D.; Wagner, D.D.; Long-Bradley, P.E.; Hairston, H. Aflatoxin Residues in Milk of Dairy Cows after Ingestion of Naturally Contaminated Grain. J. Food Prot. 1986, 49, 781–785.
- Masoero, F.; Gallo, A.; Moschini, M.; Piva1, G.; Diaz, D. Carryover of aflatoxin from feed to milk in dairy cows with low or high somatic cell counts. Animal 2007, 9, 1344–1350.
- van Eijkeren, J.C.; Bakker, M.I.; Zeilmaker, M.J. A simple steady-state model for carry-over of aflatoxins from feed to cow’s milk. Food Addit. Contam. 2006, 23, 833–838.
- Zarba, A.; Wild, C.P.; Hall, A.J.; Montesano, R.; Hudson, G.J.; Groopman, J.D. Aflatoxin M1 in human breast milk from The Gambia, west Africa, quantified by combined monoclonal antibody immunoaffinity chromatography and HPLC. Carcinogenesis 1992, 13, 891–894.
- Khlangwiset, P.; Shephard, G.S.; Wu, F. Aflatoxins and growth impairment: A review. Crit. Rev. Toxicol. 2011, 41, 740–755.
- da Silva, J.V.; de Oliveira, C.A.; Ramalho, L.N. Effects of Prenatal Exposure to Aflatoxin B1: A Review. Molecules 2021, 26, 7312.
- Smith, L.E.; Prendergast, A.J.; Turner, P.C.; Humphrey, J.H.; Stoltzfus, R.J. Aflatoxin exposure during pregnancy, maternal anemia, and adverse birth outcomes. Am. J. Trop. Med. Hyg. 2017, 96, 770–776.
- Abdulrazzaq, Y.M.; Osman, N.; Yousif, Z.M.; Trad, O. Morbidity in neonates of mothers who have ingested aflatoxins. Ann. Trop. Paediatr. 2004, 24, 145–151.
- Adhikari, M.; Gita, R.; Berjak, P. Aflatoxin, kwashiorkor, and morbidity. Nat. Toxins 1994, 2, 1–3.
- Black, R.E.; Victora, C.G.; Walker, S.P.; Bhutta, Z.A.; Christian, P.; de Onis, M.; Ezzati, M.; Grantham-McGregor, S.; Katz, J.; Martorell, R.; et al. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 427–451.
- Ortiz, J.; Jacxsens, L.; Astudillo, G.; Ballesteros, A.; Donoso, S.; Huybregts, L.; De Meulenaer, B. Multiple mycotoxin exposure of infants and young children via breastfeeding and complementary/weaning foods consumption in Ecuadorian highlands. Food Chem. Toxicol. 2018, 118, 541–548.
- Sherif, S.O.; Salama, E.E.; Abdel-Wahhab, M.A. Mycotoxins and child health: The need for health risk assessment. Int. J. Hyg. Environ. Health 2009, 212, 347–368.
- Hulin, M.; Bemrah, N.; Nougadère, A.; Volatier, J.L.; Sirot, V.; Leblanc, J.C. Assessment of infant exposure to food chemicals: The French Total Diet Study design. Food Addit. Contam. Part A 2014, 31, 1226–1239.
- Lauer, J.M.; Duggan, C.P.; Ausman, L.M.; Griffiths, J.K.; Webb, P.; Wang, J.S.; Xue, K.S.; Agaba, E.; Nshakira, N.; Ghosh, S. Maternal aflatoxin exposure during pregnancy and adverse birth outcomes in Uganda. Matern. Child Nutr. 2019, 15, e12701.
- Turner, P.C.; Collinson, A.C.; Cheung, Y.B.; Gong, Y.Y.; Hall, A.J.; Prentice, A.M.; Wild, C.P. Aflatoxin exposure in utero causes growth faltering in Gambian infants. Int. J. Epidemiol. 2007, 36, 1119–1125.
- JME (UNICEF/WHO/World Bank). Levels and Trends in Child Malnutrition: Key Fndings of the 2021 Edition of the Joint Child Malnutrition Estimates; World Health Organization: Geneva, Switzerland, 2021.
- Boni, S.B.; Beed, F.; Kimanya, M.E.; Koyano, E.; Mponda, O.; Mamiro, D.; Kaoneka, B.; Bandyopadhyay, R.; Korie, S.; Mahuku, G. Aflatoxin contamination in Tanzania: Quantifying the problem in maize and groundnuts from rural households. World Mycotoxin J. 2021, 14, 553–564.
- Ministry of Health, C.D. Gender, Elderly and Children (MoHCDGEC) ; Ministry of Health (MoH) ; National Bureau of Statistics (NBS); Office of the Chief Government Statistician (OCGS); ICF; Tanzania Demographic and Health Survey and Malaria Indicator Survey (TDHS-MIS) 2015–2016. Dar es Salaam Rockville: MoHCDGEC, MoH, NBS, OCGS, and ICF. 2016, p. 591. Available online: https://dhsprogram.com/pubs/pdf/fr321/fr321.pdf (accessed on 12 June 2022).
- UNICEF. The Conceptual Framework of Under-Nutrition. 2022. Available online: https://forum.susana.org/244-theme-1-making-the-link-in-theory-and-practice-where-do-we-stand/17576-the-conceptual-framework-of-under-nutrition (accessed on 12 June 2022).
- Gong, Y.Y.; Cardwell, K.; Hounsa, A.; Egal, S.; Turner, P.C.; Hall, A.J.; Wild, C.P. Dietary aflatoxin exposure and impaired growth in young children from Benin and Togo: Cross sectional study. BMJ 2002, 325, 20–21.
- Maxwell, S.M.; Familusi, J.B.; Sodeinde, O.; Chan, M.C.; Hendrickse, R.G. Detection of naphthols and aflatoxins in Nigerian cord blood. Ann. Trop. Paediatr. 1994, 14, 3–5.
- Magoha, H.; Kimanya, M.; De Meulenaer, B.; Roberfroid, D.; Lachat, C.; Kolsteren, P. Risk of dietary exposure to aflatoxins and fumonisins in infants less than 6 months of age in Rombo, Northern Tanzania. Matern. Child Nutr. 2016, 12, 516–527.
- Dewey, K.G.; Adu-Afarwuah, S. Systematic review of the efficacy and effectiveness of complementary feeding interventions in developing countries. Matern. Child Nutr. 2008, 4 (Suppl. 1), 24–85.
- Panjwani, A.; Heidkamp, R. Complementary feeding interventions have a small but significant impact on linear and ponderal growth of children in low- and middle-income countries: A systematic review and meta-analysis. J. Nutr. 2017, 147, 2169s–2178s.
- Bashiry, M.; Javanmardi, F.; Sadeghi, E.; Shokri, S.; Hossieni, H.; Oliveira, C.A.F.; Mousavi Khaneghah, A. The prevalence of aflatoxins in commercial baby food products: A global systematic review, meta-analysis, and risk assessment study. Trends Food Sci. Technol. 2021, 114, 100–115.
- Humphrey, J.H.; Mbuya, M.N.N.; Ntozini, R.; Moulton, L.H.; Stoltzfus, R.J.; Tavengwa, N.V.; Mutasa, K.; Majo, F.; Mutasa, B.; Mangwadu, G.; et al. Independent and combined effects of improved water, sanitation, and hygiene, and improved complementary feeding, on child stunting and anaemia in rural Zimbabwe: A cluster-randomised trial. Lancet Glob. Health 2019, 7, e132–e147.
- Null, C.; Stewart, C.P.; Pickering, A.J.; Dentz, H.N.; Arnold, B.F.; Arnold, C.D.; Benjamin-Chung, J.; Clasen, T.; Dewey, K.G.; Fernald, L.C.; et al. Effects of water quality, sanitation, handwashing, and nutritional interventions on diarrhoea and child growth in rural Kenya: A cluster-randomised controlled trial. Lancet Glob. Health 2018, 6, e316–e329.
- Luby, S.P.; Rahman, M.; Arnold, B.F.; Unicomb, L.; Ashraf, S.; Winch, P.J.; Stewart, C.P.; Begum, F.; Hussain, F.; Benjamin-Chung, J.; et al. Effects of water quality, sanitation, handwashing, and nutritional interventions on diarrhoea and child growth in rural Bangladesh: A cluster randomised controlled trial. Lancet Glob. Health 2018, 6, e302–e315.
- Hoffmann, V.; Jones, K.; Leroy, J.L. The impact of reducing dietary aflatoxin exposure on child linear growth: A cluster randomised controlled trial in Kenya. BMJ Glob. Health 2018, 3, e000983.
- Benkerroum, N. Retrospective and Prospective Look at Aflatoxin Research and Development from a Practical Standpoint. Int. J. Environ. Res. Public Health 2019, 16, 3633.
- Saltzmann, J.; Xu, Y.; Gong, Y.Y.; Lindahl, J.; Kersten, S.; Dänicke, S.; Routledge, M.N. Preliminary study on the relationship between aflatoxin-bovine serum albumin adducts in blood and aflatoxin M1 levels in milk of dairy cows. Mycot. Res. 2020, 36, 207–211.
- Chen, G.; Gong, Y.Y.; Kimanya, M.E.; Shirima, C.P.; Routledge, M.N. Comparison of urinary aflatoxin M1 and aflatoxin albumin adducts as biomarkers for assessing aflatoxin exposure in Tanzanian children. Biomarkers 2018, 23, 131–136.
- WHO. Principles for Evaluating Health Risks in Children Associated with Exposure to Chemicals; World Health Organization: Geneva, Switzerland, 2006.
- Sadeghi, N.; Oveisi, M.R.; Jannat, B.; Hajimahmoodi, M.; Bonyani, H.; Jannat, F. Incidence of aflatoxin M1 in human breast milk in Tehran, Iran. Food Control 2009, 20, 75–78.
- Mahdavi, R.; Nikniaz, L.; Arefhosseini, S.R.; Vahed Jabbari, M. Determination of aflatoxin M(1) in breast milk samples in Tabriz-Iran. Matern. Child Health J. 2010, 14, 141–145.
- Montesano, R.; Hainaut, P.; Wild, C.P. Hepatocellular carcinoma: From gene to public health. J. Natl. Cancer Inst. 1997, 89, 1844–1851.
- Gong, Y.Y.; Egal, S.; Hounsa, A.; Turner, P.C.; Hall, A.J.; Cardwell, K.F.; Wild, C.P. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: The critical role of weaning. Int. J. Epidemiol. 2003, 32, 556–562.
- Gong, Y.Y.; Hounsa, A.; Egal, S.; Turner, P.C.; Sutcliffe, A.E.; Hall, A.J.; Cardwell, K.; Wild, C.P. Postweaning exposure to aflatoxin results in impaired child growth: A longitudinal study in Benin, West Africa. Environ. Health Perspect 2004, 112, 1334–1338.
- IARC (International Agency for Research on Cancer). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; World Health Organization: Lyon, France, 1993; Volume V56, pp. 51–72.
- NIH. Mycotoxin Mitigation Trial. 2022. Available online: https://www.clinicaltrials.gov/ct2/show/NCT03940547 (accessed on 31 August 2022).
- Turner, P.C.; Moore, S.E.; Hall, A.J.; Prentice, A.M.; Wild, C.P. Modification of immune function through exposure to dietary aflatoxin in Gambian children. Environ. Health Perspect. 2003, 111, 217–220.
- Smith, L.E.; Stoltzfus, R.J.; Prendergast, A. Food chain mycotoxin exposure, gut health, and impaired growth: A conceptual framework. Adv. Nutr. 2012, 3, 526–531.
- Tessema, M.; De Groote, H.; Brouwer, I.D.; De Boevre, M.; Corominas, A.V.; Stoecker, B.J.; Stoecker, B.J.; Feskens, E.J.; Belachew, T.; Karakitsou, A.; et al. Exposure to aflatoxins and fumonisins and linear growth of children in rural Ethiopia: A longitudinal study. Public Health Nutr. 2021, 24, 3662–3673.
- Mbuya, M.N.; Humphrey, J.H. Preventing environmental enteric dysfunction through improved water, sanitation and hygiene: An opportunity for stunting reduction in developing countries. Matern. Child Nutr. 2016, 12 (Suppl. 1), 106–120.
- Harper, K.M.; Mutasa, M.; Prendergast, A.J.; Humphrey, J.H.; Manges, A.R. Environmental enteric dysfunction pathways and child stunting: A systematic review. PLoS Negl. Trop. Dis. 2018, 12, e0006205.
- Keusch, G.T.; Rosenberg, I.H.; Denno, D.M.; Duggan, C.; Guerrant, R.L.; Lavery, J.V.; Tarr, P.I.; Ward, H.D.; Black, R.E.; Nataro, J.P.; et al. Implications of acquired environmental enteric dysfunction for growth and stunting in infants and children living in low- and middle-income countries. Food Nutr. Bull. 2013, 34, 357–364.
- Humphrey, J.H.; Jones, A.D.; Manges, A.; Mangwadu, G.; Maluccio, J.A.; Mbuya, M.N.; Moulton, L.H.; Ntozini, R.; Prendergast, A.J. The sanitation hygiene infant nutrition efficacy (SHINE) trial: Rationale, design, and methods. Clin. Infect. Dis. 2015, 61 (Suppl. 7), S685–S702.
- Smith, L.E.; Prendergast, A.J.; Turner, P.C.; Mbuya, M.N.; Mutasa, K.; Kembo, G.; Stoltzfus, R.J. The Potential Role of Mycotoxins as a Contributor to Stunting in the SHINE Trial. Clin. Infect. Dis. 2015, 61 (Suppl. 7), S733–S737.
- Weber, T.K.; Polanco, I. Gastrointestinal microbiota and some children diseases: A review. Gastroenterol. Res. Pract. 2012, 2012, 676585.
- Sampson, T.R.; Mazmanian, S.K. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe 2015, 17, 565–576.
- Obuseh, F.A.; Jolly, P.E.; Kulczycki, A.; Ehiri, J.; Waterbor, J.; Desmond, R.A.; Preko, P.O.; Jiang, Y.; Piyathilake, C.J. Aflatoxin levels, plasma vitamins A and E concentrations, and their association with HIV and hepatitis B virus infections in Ghanaians: A cross-sectional study. J. Int. AIDS Soc. 2011, 14, 53.
- Onyemelukwe, G.; Ogoina, D.; Ibiam, G.E.; Ogbadu, G.H. Aflatoxins in body fluids and food of Nigerian children with protein-energy malnutrition. Afr. J. Food Agric. Nutr. Dev. 2012, 12, 6553–6566.
- Hatem, N.L.; Hassab, H.M.; Abd Al-Rahman, E.M.; El-Deeb, S.A.; El-Sayed Ahmed, R.L. Prevalence of aflatoxins in blood and urine of Egyptian infants with protein-energy malnutrition. Food Nutr. Bull. 2005, 26, 49–56.
- Coulter, J.B.; Hendrickse, R.G.; Lamplugh, S.M.; Macfarlane, S.B.; Moody, J.B.; Omer, M.I.; Suliman, G.I.; Williams, T.E.; Liverpool/Khartoum Aflatoxin Study Group. Aflatoxins and kwashiorkor: Clinical studies in Sudanese children. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 945–951.
- Hendrickse, R.G.; Coulter, J.B.S.; Lamplugh, S.M.; Macfarlane, S.B.J.; Williams, T.E.; Omer, M.I.A.; Suliman, G.I. Aflatoxins and kwashiorkor: A study In Sudanese children. Br. Med. J. (Clin. Res. Ed.) 1982, 285, 843–846.
- Ortega-Beltran, A.; Bandyopadhyay, R. Contributions of integrated aflatoxin management strategies to achieve the sustainable development goals in various African countries. Glob. Food Secur. 2021, 30, 100559.
- Fakhri, Y.; Rahmani, J.; Oliveira, C.A.F.; Franco, L.T.; Corassin, C.H.; Saba, S.; Rafique, J.; Khaneghah, A.M. Aflatoxin M1 in human breast milk: A global systematic review, meta-analysis, and risk assessment study (Monte Carlo simulation). Trends Food Sci. Technol. 2019, 88, 333–342.