Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Boren Tian and Version 2 by Rita Xu.
Osteosarcoma (OS) is a malignancy that is becoming increasingly common in adolescents. OS stem cells (OSCs) form a dynamic subset of OS cells that are responsible for malignant progression and chemoradiotherapy resistance. The unique properties of OSCs, including self-renewal, multilineage differentiation and metastatic potential, 149 depend closely on their tumor microenvironment.
- osteosarcoma
- cancer stem cell
- cell plastic
1. Introduction
Osteosarcoma (OS) is a malignancy that most commonly occurs in children and adolescents and is the second highest cause of cancer-related mortality in these groups [1][2][3][1,2,3]. There has been a rise in the annual incidence rate of OS to three cases per million individuals [4]. The majority of OS cases arises in the metaphyseal regions adjacent to the physis, including the distal femur, proximal tibia and the proximal humerus, with a strong capacity for proliferation [5]. Over the past 30 years, the treatment of OS has improved little, such that surgery accompanied with chemoradiotherapy remain as the main method of treatment [6]. Although novel clinical strategies such as gene editing, individualized treatment and novel molecular-targeted therapies, e.g., angiogenesis inhibitors, tyrosine kinase inhibitors and monoclonal antibodies, have all been deployed against OS, the outcomes for patients are poor, particularly those with more aggressive forms of the cancers [3][4][5][6][7][3,4,5,6,7]. Therefore, novel treatment strategies are in demand in clinical practice. In addition, the molecular mechanism underlying tumorigenesis and malignant metastasis needs to be studied in detail.
Based on the present research, OS is speculated to have two main origins, bone mesenchymal stem cells (BMSCs) and osteoblasts [8][9][10][8,9,10]. p53 as a classic cancer suppressor gene plays a key role in OS progression. The deficiency of p53 is an important reason leading to primary OS. In addition, retinoblastoma gene (Rb), cyclin dependent kinase inhibitor 2 (CDKN2), KRAS and c-Met also participate in the regulation of OS progression [8][9][10][11][8,9,10,11]. Within cancer tissues, there exist several dynamic subsets of cancer cells considered to be cancer stem cells (CSCs) or stem cell-like cancer cells [12][13][12,13]. CSCs have been frequently reported to exhibit stem cell properties and capabilities of long-term clonal proliferation, tumorigenicity, facilitating metastasis and promoting resistance to chemotherapy and radiotherapy [14][15][14,15]. Therefore, exploring the origins of cancer initiation and metastasis will likely facilitate the development of future therapies. In 1994, Lapidot et.al first reported that, in human acute myeloid leukemia, a rare population of CSCs exists [16]. Subsequently, an accumulating number of studies have also reported the existence of CSCs in other solid tumors, including prostate, glioblastoma, hepatoma, breast cancer and OS [17][18][19][20][21][22][17,18,19,20,21,22]. In fact, all types of malignant tumors consist of different subpopulations of tumor cells, leading to high degrees of heterogeneity.
The niche in which CSCs reside is the tumor microenvironment, where they co-exist with adjacent supporting cells, micro-vessels and the extracellular matrix [19][20][19,20]. In addition, the tumor microenvironment can contain soluble factors, such as chemokines and cytokines, whilst being under the influence of various mechanical factors, including matrix stiffness, solid stress and fluid stress [23][24][23,24]. In the OS microenvironment, OS stem cells (OSCs) are contained in a specialized niche that contains a unique bone microenvironment, which consists of various types of bone cells, such as osteoblasts or osteoclasts. OSCs are similar to other CSCs, in that they account for a proportion of cancer cells with tumorigenic and self-renewal capabilities. The existence of OSCs was first confirmed by Gibbs et al., who found that when primary human OS cells or the OS cell line MG63 were suspended in a serum-free medium with defined growth factors, 0.1% of the cells could form spheres with self-renewal capacity [17][18][19][20][21][22][23][24][25][17,18,19,20,21,22,23,24,25]. Subsequently, a series of studies have proven the existence of OSCs, in addition to revealing the phenotype and possible marker profile of OSCs. The recent studies on possible OSC markers and phenotypes (Table 1).
Table 1. Putative OSC markers and phenotypes.
| Marker | Cell Origin | Phenotype | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CD133 | Saos-2, MG-63, U2-OS, MNNG/HOS, 143B, HOS, Human primary cells | High stem cells gene expression, sphere formation, side population, increased cell proliferation [26][27][28][29][30]. | High stem cells gene expression, sphere formation, side population, increased cell proliferation [26,27,28,29,30]. | ||||||||||||||||||||
| CD117/Stro-1 | K7M2, KHOS/NP, MNNG/HOS, 318–1, P932, BCOS | High stem cells gene expression, sphere formation, drug resistance, in vivo tumorigenicity and metastatic potential [17][18][19][20][21][22][23][24][25][26][27][28][. | High stem cells gene expression, sphere formation, drug resistance, in vivo tumorigenicity and metastatic potential [17,18 | 29 | ,19 | ][ | ,20 | 30][ | ,21 | 31 | ,22 | ] | ,23,24,25,26,27,28,29,30,31]. | ||||||||||
| CD271 | Human primary (FFPE), MNNG/HOS, U2-OS, Saos-2 | High stem cells gene expression, sphere formation, drug resistance, in vivo tumorigenicity [32]. | |||||||||||||||||||||
| Aldehyde dehydrogenase | MG-63, OS99–1 Hu09, Saos-2 | High stem cells gene expression, sphere formation, drug resistance, increased cell proliferation [33][34]. | High stem cells gene expression, sphere formation, drug resistance, increased cell proliferation [33,34]. | ||||||||||||||||||||
| Stem cells antigen-1 | 4 Murine osteosarcoma cell lines | Sphere formation, in vivo tumorigenicity [35][36] | Sphere formation, in vivo tumorigenicity [35,36] | ||||||||||||||||||||
| Fas apoptotic inhibitory molecule 2 | MNNG/HOS, U2-OS | Sphere formation, drug resistance, in vivo tumorigenicity [37]. | |||||||||||||||||||||
| Side population | OS2000, KIKU, NY, Huo9, HOS, U2OS, Saos-2, human primary |
High stem cells gene expression, Sphere formation, in vivo tumorigenicity, self-renewal, apoptosis resistant [38][39][40]. | High stem cells gene expression, Sphere formation, in vivo tumorigenicity, self-renewal, apoptosis resistant [38,39,40]. | ||||||||||||||||||||
| Sphere formation | MG-63, MNNG/HOS, human primary | High stem cells gene expression, drug resistance, in vivo tumorigenicity [17][18][19][20][21][22][23][24][25[42]. | High stem cells gene expression, drug resistance, in vivo tumorigenicity [17,18 | ][ | ,19 | 26][ | ,20 | 27 | ,21,22,23,24,25 | ][28][29][30][31 | ,26 | ] | ,27 | [ | ,28,29 | 32 | ,30,31 | ] | ,32 | [33] | ,33 | [41] | ,41,42]. |
2. Role of the Tumor Microenvironment in Regulating OSC Stemness
OSCs can interact with their microenvironment through complex and dynamic processes, including variation of oxygen, mechanical interactions, enzymatic modification of the extracellular matrix (ECM) structure and signaling cross-talk, all of which can influence the progression and the dissemination of OS cells (Figure 1)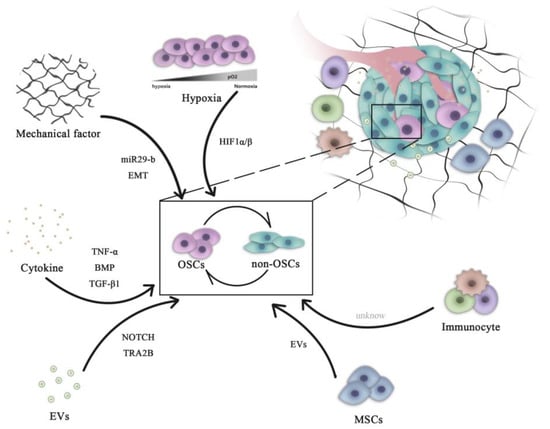
Figure 1. Role of microenvironment signaling in regulating OSC and non-OSC reversion. Complex pathways are necessary for the maintenance of the homeostasis of the OSC population. All microenvironment components, including cells (mesenchymal cells and immune cells) and non-cellular factors (hypoxia, cytokines and mechanical EVs), can influence the dynamic transition between OSCs and non-OSCs. OSCs, osteosarcoma stem cells; miR, microRNA; HIF, hypoxia-inducible factor; EVs, extracellular vesicles; EMT, epithelial-mesenchymal transition; BMP, bone morphogenetic protein; TNF, tumor necrosis factor; TGF, transforming growth factor; TRA2B, transformer 2β homolog.