Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Esraa Mahmoud and Version 2 by Conner Chen.
Hepatocellular carcinoma (HCC) is a global healthcare challenge, which affects more than 815,000 new cases every year. Activated hepatic stellate cells (aHSCs) remain the principal cells that drive HCC onset and growth. aHSCs suppress the anti-tumor immune response through interaction with different immune cells. They also increase the deposition of the extracellular matrix proteins, challenging the reversion of fibrosis and increasing HCC growth and metastasis.
- hepatocellular carcinoma
- hepatic stellate cells
- fibrosis regression
1. Introduction
Liver cancer is a global health problem, with an estimated increase of 32% by 2040 [1]. Representing 90% of liver cancers, hepatocellular carcinoma (HCC) causes 700,000 deaths annually.
Although HCC pathogenesis is complex and varies depending on underlying etiology, the usual background setting for HCC is liver injury, chronic inflammation, irreversible fibrosis, and cirrhosis [2]. In fact, 80–90% of HCC develops in the fibrotic or cirrhotic liver [3]. Hepatic stellate cells (HSCs) play a key role in this sequence of events, contributing mainly to liver fibrosis and cirrhosis. They are liver-specific mesenchymal cells, which are located in the perisinusoidal space in contact with different cell types [4]. In a healthy liver, HSCs exist in a quiescent non-proliferative state as an important source of paracrine, autocrine, and chemoattractant factors to maintain hepatic homeostasis [5]. Quiescent HSCs are very sensitive to extracellular pro-fibrotic signals [6] and contain numerous vitamin A lipid droplets, which are essential for the proper function of the immune system [7]. When toxins or viruses injure the liver, damaged hepatocytes and immune cells secrete signals, which could activate HSCs into myofibroblast-like cells [8]. Activated HSCs produce an extracellular matrix (ECM) at the site of injury as a temporary protective scar to prevent further damage, initiating the first steps of fibrosis [8][9][8,9]. Long-acting agents maintain the activation of HSCs, increasing their capabilities for proliferation and migration [10]. Activated HSCs produce more ECM, leading to chronic fibrosis and cirrhosis and eventually to HCC (Figure 1) [6]. Despite significant advances in the treatment of HCC, drug-resistance is a critical obstacle [11], and the 5-year survival rate is low (5–14%) [12], but survival rates of greater than 20% have been reached in some regions [13]. Therefore, there is a potential to increase survival rates. Given the fact that available therapies can activate HSCs, synchronous targeting activated hepatic stellate cells (aHSCs) may be beneficial for patients [14].
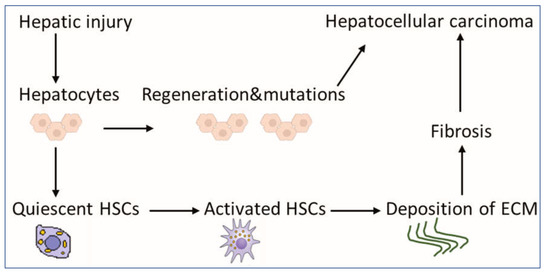
Figure 1. Scheme for the contribution of HSCs to liver pathology. HSCs exist in a quiescent state, containing numerous vitamin A lipid droplets. When the liver is injured, damaged hepatocytes mediate HSC activation, which could produce a large amount of ECM, leading to fibrosis as an indirect mechanism of HCC. Mutations during the regeneration of hepatocytes may lead directly to the development of HCC. Abbreviations: HSCs: hepatic stellate cells, HCC: hepatocellular carcinoma and ECM: extracellular matrix.
2. The Role of aHSCs in HCC
2.1. The Suppression of the Antitumor Immune Response by aHSCs
In HCC, aHSCs receive signals from individual immune cells, and, in turn, they produce soluble mediators, acting on surrounding immune cells [15]. HSC mediators could orchestrate both innate and adaptive immunity, resulting in an immunosuppressive tumor microenvironment [7]. Reduction of antitumor responses was shown in immunocompetent mice after co-transplantation of HSCs and HCC cells [16]. Such cotransplantation of HSCs inhibited systemically lymphocyte infiltration, which promoted tumor cell proliferation and, therefore, HCC growth; the size of the tumors was HSC dose-dependent [16]. Previous experiments addressing HCC–HSC interactions were performed on immunodeficient mice, and, therefore, the effect of HSC on the immune system was not investigated. Using immunocompetent mice, the authors were able to define HSC-immune interactions in HCC [16]. The antigen-directed cytotoxicity of T lymphocytes (TLs) boosts the immune response against cancer [17]. Activation and proliferation of TL in tumor tissue, predominantly CD8+ and CD4+ T lymphocytes, can control HCC progression [18]. HSCs can exert their immunomodulatory activities by downregulating the number and function of CD4+ and CD8+ TLs [19]. Contrary to quiescent HSCs, aHSCs in mice and humans expressed programmed death-ligand 1 (PD-L1) to inhibit TL responses [7]. PD-L1 expressed by HSCs can induce TL apoptosis, attenuate TL infiltration, and suppress TL-mediated cytotoxicity, therefore inhibiting TL responses and enabling tumor cells to escape the host immune response [20]. In addition, HSCs may prevent the local stimulation of naive TLs [21]. In Hepa1–6 cells, activated HSCs induced the death of activated TLs and reduced the cytotoxicity of cancer-specific TLs, which resulted in the increased proliferation and migration of cancer cells [22]. More investigation of the role of aHSCs in the apoptosis of TLs in HCC patients is needed. aHSCs also induce expansion of two suppressive immune cell populations; myeloid-derived suppressor cells (MDSCs) [14] and T helper 17 (Th17) cells, a subset of CD4+ effector T cells [23]. MDSCs play a pivotal negative role in the immune response through the inhibition of cytotoxic T cells and recruitment of regulatory T cells, which results in tumor progression [24]. HSCs induce MDSC accumulation in the tumor tissue by the stimulation of the COX2–PGE2–EP4 pathway [25]. Inhibition of this pathway in murine orthotopic HCC models downregulated MDSCs and HCC growth [25]. Immunosuppressive functions of Th17 cells may contribute to HCC progression [26]. IL-17A produced by Th17 could increase cancer cell motility via the activation of the nuclear factor-kB (NF-kB) transcript factor, increasing HCC metastasis [27]. Culturing CD4+ cells with HSCs (extracted from hepatitis B virus-related fibrotic liver tissue) increased the percentages of Th17 cells [23]. HSCs may secrete high levels of interleukin-6 as a critical initiator of Th17 expansion and tumor necrosis factor-α as a key regulator of Th17 differentiation [28]. Interestingly, previous data indicated suppression of Th17 differentiation by mouse HSCs [29]. Critical evaluation of Th17-HSCs interactions could be addressed in appropriate mouse models of HCC. Macrophages polarize in the liver with strong plasticity into pro-inflammatory M1 or anti-inflammatory M2 in response to local signals from the tumor microenvironment [30][31][30,31]. M1 macrophages are thought to be tumoricidal, while M2 macrophages are usually believed to promote tumorigenesis and tumor progression [32]. M2 macrophages in HCC promote the invasion and migration of tumor cells [33]. M2 macrophage-derived CCL22 was proven to enhance tumor migration through the activation of epithelial–mesenchymal transition [34]. aHSCs recruited CCL2/CCR2 pathway in HCC cell lines to stimulate M2 phenotypic transformation [35]. M2 macrophage polarization could lead to the progression of HCC [36]. Natural killer (NK) cells defend the body against tumors by engaging death-inducing receptors to stimulate cancer cell apoptosis. HCC patients with low intratumoral NK cells infiltration have shorter disease-free survival [37]. In animal models of fibrosis, transforming growth factor-β secreted by HSC could inhibit NK cell function [38]. On the other hand, NK cells could induce apoptosis of aHSCs in hepatitis C virus-infected patients [39] and mouse models of fibrosis [40]. Studies on the interaction between aHSCs and NK cells in HCC models should be investigated. Dendritic cells (DCs) can activate antitumor immunity by priming TL against cancer-progression-associated antigens. HSCs induce the expression of dendritic-cell-derived immunoglobulin receptor 2 (DIgR2), which inhibits DC-induced antigen-specific TL responses [41]. DIgR2 was shown to bind to the receptor in TLs, suppressing TL proliferation, cytokine production, and cytotoxic TL activity [41]. Co-culturing of tumor-HSCs (isolated from the tumor) to DCs induced the expression of DIgR2, in contrast to quiescent HSCs, which had no significant effect on DIgR2 expression. Considering quiescent HSCs in such studies boosts the role of activated HSCs in HCC [41]. Although the role of immune system in liver cancer is complex [42], the overall role of aHSCs in immune regulation is pro-oncogenic [16]. Exploring the interaction between different immune cells and HSCs in established HCC models could highlight the main targets to improve immune surveillance against HCC.2.2. HSCs Upregulate the Deposition of ECM for the Development of Fibrosis and HCC
Under normal conditions, the rate of ECM production in the liver equals that of its degradation, resulting in no net accumulation of the matrix. Fibrogenesis occurs when there is an imbalance between ECM production and degradation [43], resulting in the impairment of liver functions, which may eventually lead to cirrhosis and HCC [44]. Fibrosis and cirrhosis are reported clinically as reversible processes [45]. Reversibility of liver fibrosis depends mainly on the degradation of ECM [46]. aHSCs are able to increase matrix protein synthesis, which might lead to the irreversibility of fibrosis and favor progress and metastasis of HCC (Figure 2).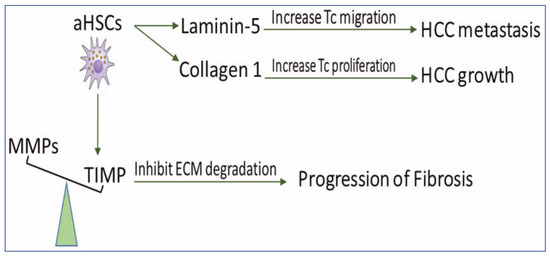
Figure 2. The scheme demonstrates the role of HSCs in disturbing ECM balance for the development of fibrosis and HCC. aHSCs express TIMP glycoproteins which inhibit MMP-mediated degradation of ECM and, therefore, prevent fibrosis regression. They also secrete collagen I, which increases tumor cell proliferation and produces Ln-5, which promotes tumor cell migration. MMPs: matrix metalloproteinases, TIMPs: tissue inhibitors of MMP, aHSCs: activated hepatic stellate cells, HCC: hepatocellular carcinoma, ECM: extracellular matrix, and TC: tumor cell.