Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Sabry Nasif Soliman and Version 2 by Camila Xu.
Strigolactones (SLs) are carotenoid derivatives that occur naturally in plants and are defined as novel phytohormones that regulate plant metabolism, growth, and development. Strigolactone assists plants in the acquisition of defensive characteristics against drought stress by initiating physiological responses and mediating the interaction with soil microorganisms.
- strigolactones
- abiotic stress
- ecological microbiome
1. Introduction
Abiotic stresses are the most significant limiting factors of plant survival and growth under the increasing crisis of climatic changes. Numerous studies tried conducted to find a solution for different plant species to cope with various stressors. Plant hormones (phytohormones) are organic molecules that cause signaling effects in plant tissues. Cell elongation, phototropism, stress tolerance, apical dominance, plant growth improvement, senescence, and dormancy are a few of many processes considered as physiological functions of phytohormones. The impacts on plants are substantial despite the minimal concentration of secretion and significantly improve plant tolerance against abiotic stress [1]. In order to adapt to adverse conditions, plants have developed various responses by evoking several signals that cause metabolic and genetic pathways to be reprogrammed [2][3][2,3].
Strigolactones (SLs), was identified as plant hormones that play regulatory roles against abiotic stresses in plants, due to their essential role in regulating plant growth and development [4][5][4,5]. The first naturally occurring germination stimulant for Striga was isolated as early as 1966 from root exudates of cotton (Gossypium hirsutum L.), which is neither a host for Striga nor Orobanche [6][7][8][6,7,8]. SLs are a class of terpenoid-derived compounds that were first discovered as (+)-strigol, which stimulates seeds germination from the parasitic plant Striga [6]. Many researchers reported strigolactone (SL) as a newly identified phytohormone [9][10][11][9,10,11]. Strigolactones improve control the development patterns and interactions between nearby colonies in moss [12][13][12,13] Furthermore, SLs participate in metabolic processes acting against biotic and abiotic stress [4][14][15][4,14,15]. The production of SLs in plants is strongly controlled and influenced by the various kinds of stressors that they experience at different growth phases. Recent evidence of interactions between SLs and other phytohormones, such as abscisic acid, in plant responses to abiotic stressors, implies that SLs actively engage in phytohormone-controlled regulatory networks of plant stress adaption [16].
Several studies were focused on SLs since they strongly demonstrate many internal and external responses to plant growth and development [5][8][17][5,8,17]. They regulate lateral roots and root hairs, gravitropism, soil microbes, vasculature development, nutrient and photoassimilate capture and allocation, light responses, leaf shape, leaf senescence, drought, and salinity tolerance [18]. Crosstalk with other known hormones was discussed by several investigations, and the signaling between SLs and other plant hormones may demonstrate their physiological function. Plants regulate their growth by simultaneously sensing and responding to both the external environmental signals and the internal developmental signals [1][4][1,4]. For instance, plant phytohormones may have responses to the signaling pathways of photoreceptor phytochromes for improving the photosynthesis processes. The phytohormones also affect photoreceptor signal transduction at cellular levels [18]. The crosstalk of strigolactone with other phytohormones is widely investigated. Phenomena caused by these interactions, such as defense against abiotic stress and attraction of microorganisms, have been explored. Plant morphological adaptability in response to changes in environmental factors is largely influenced by phytohormones. Strigolactones (SLs) are carotenoid-derived hormones that affect various aspects of development and interaction with microorganisms. They have been proposed as mediators of environmental stimuli in resource allocation processes; as a result, their pathways must be responsive to environmental cues in order to contribute to adaptive adjustments [19][20][19,20]. The research studies indicate that plants make efficient use of their secondary metabolites to defend themselves and make their environment suitable. For instance, plants actively release volatile compounds to repel herbivores and, at the same time, to draw in natural enemies that are specifically adapted to fight the herbivores [21][22][21,22].
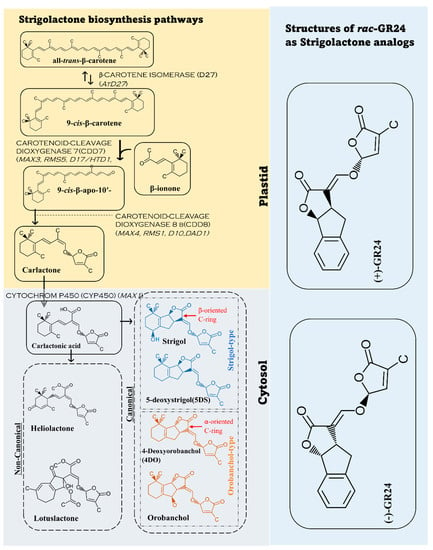
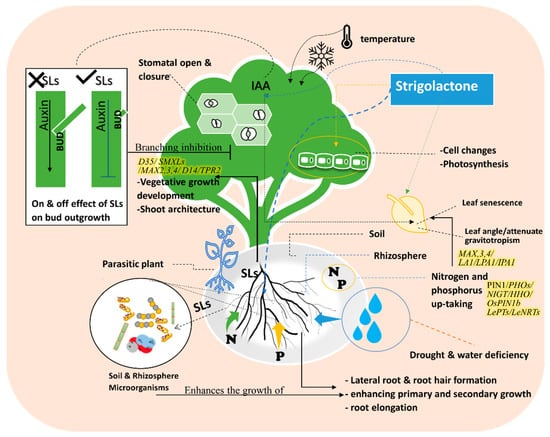
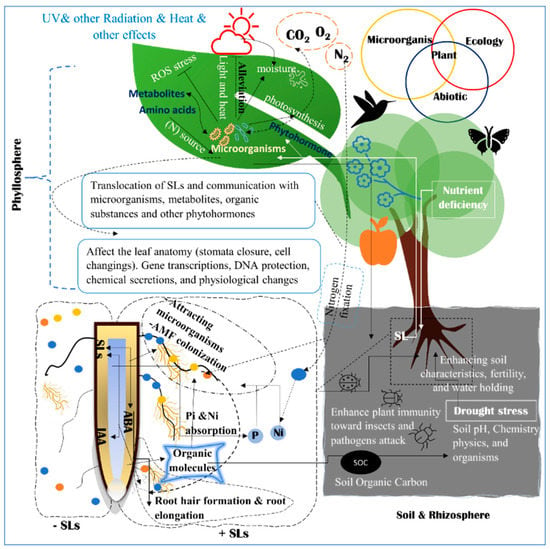
2. Strigolactone Biosynthesis, and Signaling Pathways
2.1. Biosynthesis Pathway
Biosynthesis of the strigolactone molecules and SL-like compounds is carried out by β-CAROTENE ISOMERASE (D27) and a group of other enzymes known as CAROTENOID-CLEAVAGE DIOXYGENASE (CCDs group). There are two main dioxygenases groups including CCD7 and CCD8 [23][30]. In the biosynthetic pathway, D27 has 2 ways of dissociation by converting all-trans-β-carotene to 9-cis-β-carotene and vice versa. 9-cis-β- carotene is converted to 9-cis-β-apo-10′carotenal as CCD7 attacks the 9′ and 10′ bonds of β -carotene, and CCD8 cleaves 9-cis-β-apo-10′carotene and forms carlactone. Carlactone is transformed into carlactonic acid (CLA) in the cytoplasm by sub family CYTOCHROM P450 (CYP450). Carlactonic acid leads to the formation of 5-deoxystrigol(5DS),4-deoxyorobanchol(4DO), and other forms of SLs [24][25][26][31,32,33]. Although the reactions of converting all-trans-β-carotene happen in plastids, the formation of strigolactones are carried out in the cytoplasm. This happens when carlactone (CL), the precursor of strigolactones, is transported to the cytoplasm where strigolactone and SL-like compounds are formed [24][31]. Different plants have different enzyme members from the families reswearchers mentioned. [15][26][27][28][29][15,23,33,34,35]. Many synthetic analogs such as GR24 have been developed and studied. Further studies about synthetic analogs and their characteristics and effects need to occur. The biosynthetic pathway and the common analogs GR24 are explained in Figure 1 [27][28][29][30][31][23,34,35,36,37].Figure 1. Strigolactone biosynthesis from carotenoids. In plastids all–trans–β–carotene goes through different enzymatic reactions to carlactone. Then, the carlactone transforms to form carlactonic acid in the cytosol (cytoplasm) to form various forms of strigolactones. The figure demonstrates two common examples of SLs in non-canonical form, four common ones of canonical form and the most common strigolactone analog, GR24.
2.2. Signaling Pathways
2.2.1. Impact on Branching and Leaf-Stem Angle with Relatioship to Gravitropism
D53-like SMXLs regulate leaf morphology and SL-induced SMXL6 degradation requires D14 and MAX2 [32][33][38,39]. D53-like SMXLs interact with MAX2 and D14. D53-like SMXLs interact with TPR2 and exhibit transcriptional repression [33][39]. It is believed that D53 regulates the expression of genes essential for the development of secondary shoots in the nucleus [34][40]. In the signaling activation of branching, auxin regulates “MORE AXILLARY GROWTH” MAX3 and MAX4 gene expression, these two genes are also influenced by strigolactone during branching inhibition control [35][41]. MAX3 and MAX4 participate in shoot branching and architecture signaling through the gravitropism isolation of LAZY1(LA1) suppressors, which revealed the involvement of SLs in shoot gravitropism/rice tiller angle Figure 2. SLs attenuate shoot gravitropic response in rice by modifying trilling angles. SL regulation of rice shoot gravitropism is dependent on indigenous auxin levels [36][42]. SL-mediated shoot gravitropism is conserved in Arabidopsis and many other plant species. For instance, it was reported in rice that SLs is a moderator of some genes such as LAZY1(LA1), LOOSE PLANT ARCHITECTURE1(LPA1), and IDEAL PLANT ARCHITECTURE1 (IPA1), which are involved in shoot branching inhibition and shoot gravitropism. Thus, SLs regulate tiller/branch angle in different plant species, indicating that shoot gravitropism is the key component dictating the proper positioning of shoot branches (Figure 2) [36][37][42,43].Figure 2. Effects of Strigolactone analogs (e.g., GR24) on the plant’s vegetative growth and signaling pathway under abiotic stress in relation to plant biological characteristics and soil & rhizosphere microbiome. The figure shows the role of SLs on nitrogen and phosphorus uptake. Root and Shoot architecture changes identify the effects of SLs in modifying the plant structure to cope with the abiotic stress. The figure demonstrates that SLs impact shoot branching. The plant utilizes SLs to preform mechanisms that defend itself against different abiotic stresses such as drought, senescence, temperature, light, and nutrient deficiency.
2.2.2. Impact on Low-Light Stress
GR24 application increased the activity and gene expression of antioxidant enzymes, and it reduced malonaldehyde (MDA) and hydrogen peroxide (H2O2) content in Low Light stressed plants (LL-stressed plants) [38][39][44,45]. These results suggested that exogenous application of GR24 enhances plant tolerance to LL and improving photosynthesis by promoting utilization of light energy to alleviate photosystem injuries induced by excess light energy and ROS as well as enhancing photosynthesis efficiency to improve plant growth [39][40][45,46]. Exogenous GR24 application on tomato seedlings reduces the negative effects of low light exposure in at least three ways: by reducing growth inhibition, improving photosynthetic efficiency, and relieving oxidative stress [39][41][42][45,47,48]. In addition, the application of GR24 effectively alleviates the photoinhibition of photosystems I and II (PSII and PSI) under high light stress mainly by balancing excitation energy and promoting the electron transfer chain between two photosystems, thus enhancing CEF, PQ pools, and quantum yield of PSII and PSI photochemistry [39][45]. Furthermore, plants growing in LL exhibit reduced levels of the enzymes photosystem II (PS II), ATP synthase, cytochrome b/f, and ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), as well as poorer electron transport (ETR) and CO2 consumption [39][43][45,49].2.2.3. Impact on Nitrogen and Phosphorus Deficiency
SLs consider the modulating expression factor of regulatory genes as a signaling pathway for nitrogen (N) and phosphorus (P) starvation defense mechanisms [44][50]. Therefore, many regulatory genes are involved in N and P regulations as well as signaling pathways of N–P integrators PHOs family [44][45][46][50,51,52]. SLs also regulate NIGT/HHO involved in the phosphorus deficiency signaling pathway [44][50]. LePTs and LeNRTs families are responsible for improving phosphorus and nitrogen absorption efficiency [44][46][47][48][50,52,53,54]. A signaling impact of D3-dependent and SLs biosynthesis in the suppression of tiller bud outgrowth under Pi deficit was identified in several SL signaling pathways [47][49][26,53]. OsPIN1b responds to low levels of N and P and regulates the activities in the root apical meristem, which leads to the rice seminal root elongation (Figure 2 and Figure 3) [50][55].Figure 3. The complex relations between plants, ecology, and microorganisms. The rhizosphere, soil, and root system interact together to alleviate or eliminate the effects of abiotic stress. The phyllosphere ecosystem in the top portion of the figure has a different microbiome. Phyllosphere is composed of all parts above the ground. There are different mechanisms to alleviate abiotic stress with the cooperation of the surrounding ecosystems, physiological processes, and participation of phytohormones and organic substances. The relationship between plants and organisms in the rhizosphere, phyllosphere, and endosphere is demonstrated.