Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Muhammad Shahid Javaid and Version 2 by Vivi Li.
The challenges in making animal models of complex human epilepsy phenotypes with varied aetiology highlights the need to develop alternative disease models that can address the limitations of animal models by effectively recapitulating human pathophysiology. The advances in stem cell technology provide an opportunity to use human iPSCs to make disease-in-a-dish models.
- stem cell lines
- epilepsy
- patient-specific cell models
- neuronal differentiation
- personalised disease models
- personalised medicine
1. Introduction
Epilepsy affects 70 million people worldwide [1][2][1,2], causing significant morbidity and mortality. with more than half of the affected people living in countries with poor medical resources and little or no access to treatment [3]. It is characterised by unprovoked, recurrent seizures which result from the disruption in the balance between neuronal excitation and inhibition in the brain [1]. In most cases the cause of epilepsy is unknown but both genetic and environmental factors are understood to contribute to its aetiology.
Genetic epilepsy is characterised by seizures that are the result of known genetic variance in one or many genes associated with epilepsy [2]. Mutations in several genes encoding ion channels and proteins have been reported to be most commonly associated with epilepsy. These genes include, but are not limited to mutation in SCN1A (encode sodium channel protein), which is associated with Dravet syndrome; KCNQ2 or KCNQ3 (both encoding potassium channel protein), which are associated with benign neuronal familial seizures [3][4][5][3,4,5]; the CHRNA4 gene (20q13), which is associated with Autosomal Dominant Nocturnal Frontal Lobe Epilepsy (ADNFLE), characterised by hypermotor nocturnal seizures [6]; and the mammalian target of rapamycin (mTOR) pathway, in particular DEPDC5 gene of this pathway, which are associated with Focal Cortical Dysplasia (FCD) type IIa and IIb [7][8][9][10][11][12][13][14][15][7,8,9,10,11,12,13,14,15].
Anti-seizure medications (ASMs) are the mainstay of treatments for epilepsy. Despite multiple newly developed drug treatments being introduced to clinical practice, more than 30% of epileptic patients remain drug-resistant [16]. Genetic causes account for almost 20% of drug-resistant epilepsy cases in children. Surgery may be the only curative treatment option for these refractory epilepsy patients. However, ASMs and surgery are not always successful due to an incomplete understanding of epilepsy aetiology and pathogenesis resulting in non-targeted treatments [16][17][18][19][16,17,18,19]. There is, therefore, a need for in-depth investigations to gain a better understanding of the pathological mechanisms, an understanding which would inform the development of new treatments.
Currently, potential ASMs are validated using acute-seizure animal models prior to clinical development. However, animal models of genetic epilepsy have several limitations [20][21][22][23][20,21,22,23]. The major concern is the species-specific differences [23] leading to differences in physiological development and lack of human-specific receptors and drug targets. Hence, there is a demanding need for human-based disease models to develop new therapeutic strategies to achieve seizure freedom for these patients.
The development of human-based in vitro disease models [24] is an active area of research. One potential source of human in vitro models is the use of human pluripotent stem cells. These models can either be derived from human embryonic stem cells (hESCs) or induced pluripotent stem cells (iPSCs). The iPSCs derived from healthy individuals or patients are a promising approach to developing regenerative therapies as well as in vitro models of pathophysiological mechanisms of diseases (Figure 1) [25].
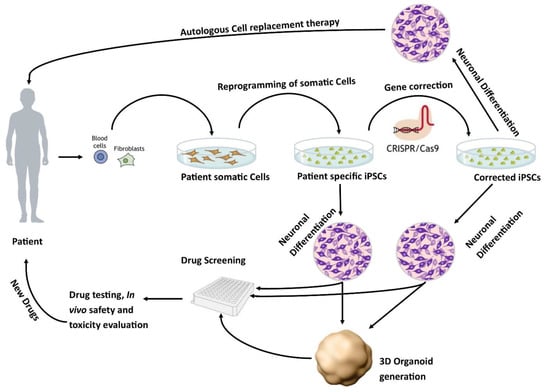
Figure 1. Patient-specific stem cell lines for cell replacement therapy and new drug development. Somatic cells from the patient’s skin or blood can be isolated and reprogrammed into iPSCs. These cells have the potential to differentiate into different neuronal populations and can be used to either further study the pathophysiology of the disease or as a drug screening system to test new potential therapies. In addition, these lines can be corrected using CRISPR/Cas9 to generate isogenic controls which provide ideal controls.
These models better recapitulate the complexity of genetic epilepsy. In recent years, stem cell research has focused on using patient-specific hiPSC-derived neurons as in vitro models of epilepsy.
2. Gene Editing Techniques for Disease Modelling and Isogenic Control
The methodology used to induce genetic variance in healthy cells for disease modelling, or to correct patient cells for isogenic control, has evolved over time. Sixteen studies (Table S1) reported the use of gene editing techniques to create isogenic controls or to study the disease-causing genetic variation in patient-derived iPSCs. Transcription Activator-Like Effector Nucleases (TALEN) was one of the first gene-editing techniques used in 2014 to generate the SCN1A mutation in human iPSCs [26][61] and was subsequently used in two more studies [27][28][48,55]. Zinc-Finger Nucleases (ZFNs) is another gene-editing technology and was used to model TSC in 2016 [29][35]. In addition, virus- or vector-based knock-out and knock-down techniques were also used in a few studies [30][31][32][33][33,42,95,106]. These methods were subsequently replaced with the more advanced CRISPR/Cas9 method. Unlike its predecessors, TALEN and ZFNs, it is precise, robust, and site-specific with fewer off-target effects [34][113]. The first use of CRISPR/Cas9 technology in epilepsy was reported in 2016 to generate a loss of function SCN1A mutation in human iPSCs to gain insight into Dravet syndrome [35][114]. This approach has been widely adopted in recent years with eight additional studies generating disease-specific neurons and isogenic controls using CRISPR/Cas9 [35][36][37][38][39][40][41][42][37,44,68,70,74,97,112,114].
3. Epilepsy Patient-Specific iPSCs Derived Disease Models
Dravet syndrome was the most commonly studied [26][35][38][43][44][45][46][47][48][49][60,61,62,63,64,65,67,68,69,114], followed by tuberous sclerosis [29][39][50][51][52][35,38,70,71,72], focal cortical dysplasia [53][54][55][56][57][82,83,84,85,104], and a few rare epilepsy syndromes [37][40][58][59][60][61][62][63][64][65][66][67][68][69][44,45,46,47,53,73,74,76,77,79,80,81,87,115]. These models have the potential to provide useful information about the involvement of a particular gene in disease progression and its anticonvulsant response.
The first in vitro model from a Dravet patient carrying a mutation in the SCN1A gene was generated in 2013. The findings from that study showed that the loss of function in GABAergic inhibition appears to be the main driver in epileptogenesis [44][62]. Since then, there have been several studies assessing various mutations in the SCN1A gene [26][27][28][35][38][43][44][45][46][47][48][70][71][72][48,54,55,56,60,61,62,63,64,65,66,67,68,114]. Neurons derived from these patients exhibit increased sodium currents and hyperexcitability [46][64], which can be alleviated by treatment with phenytoin [43][60]. In another study, neurons derived from two patients with Dravet syndrome demonstrated that genetic alterations of SCN1A differentially impacted electrophysiological impairment. The degree of impairment corresponded with the symptomatic severity of the donor from which the iPSCs were derived [45][63]. Recently, another patient-specific iPSCs-derived neuronal study generated from individuals with SCN1A mutation indicated an imbalance in excitation and inhibition that leads to hyperactivity in the neural network. This study used homozygous and isogenic controls to show the hyperexcitability in the generated neurons [38][68]. These studies indicated that neurons could recapitulate the neuronal pathophysiology and could potentially be used for screening drugs for personalised therapies [43][46][60,64].
The most commonly studied mTORopathies were tuberous sclerosis and focal cortical dysplasia (FCD). A study using neurons derived from a patient with TSC2 mutation reported hyperactivation of mTORC1 pathway [29][35]. In this model, pharmacological inhibition of mTORC1 with rapamycin reverses developmental abnormalities and synaptic dysfunction during independent developmental stages [29][35]. In another study, neuronal progenitor cells (NPCs) generated from a patient carrying a heterozygous TSC2 mutation exhibited disrupted neuronal development, potentially contributing to the disease neuropathology. Moreover, NPCs also exhibited activation of mTORC1 downstream signalling and attenuation of PI3K/AKT signalling upstream of TSC [52][72]. More recently, NPCs generated from the patient carrying TSC germline nonsense mutation in exon 15 of TSC1 showed the influence of TSC1 mutation in the early neurodevelopmental phenotypes, signalling, and gene expression in NPCs compared to the genetically matched wild-type cells [39][70]. In 2019, Sundberg et al showed that loss of one allele of TSC2 is sufficient to cause some morphological and physiological changes, elevated phosphorylation, and hyperexcitability of mTORC1 in human neurons, but biallelic mutations in TSC2 are necessary to induce gene expression dysregulation seen in cortical tubers. They also found that treatment of TSC2 patient-specific iPSCs-derived neurons with rapamycin reduced neuronal activity and partially reversed gene expression abnormalities [51][71]. In 2020, Alsaqati used commercially available TSC2 (loss of function mutation) patient-derived iPSCs and reported that the dysfunctional neuronal network behaviour in the differentiated neurons could not be rescued with rapamycin treatment [73][57]. The difference in response in these two studies is because of two different iPSCs samples carrying different mutations in the TSC2 gene [51][73][57,71].
Five studies assessed the FCD-related cortical malformation by generating iPSCs from patients with mutations in genes involved in regulating the mTOR pathway. In 2017, Marinowic et al described the generation of iPSC-based cellular models of refractory epilepsy from the fibroblasts of two refractory epilepsy patients with FCD type IIb, one a 45-year-old male and the other 12-years-old female [55][84]. Then in 2020, Marinowic published another study using these cells investigating the differences in the migration potential and the expression of genes for cell proliferation, adhesion, and apoptosis. The main finding of the study was that the gene expression was different between the neurons generated from the adult male compared to the child. They concluded that differences in the migration potential of adult cells, and differences in the expression of genes related to the fundamental brain development processes, might be associated with cortical alteration in the two patients with FCD IIb [56][85]. In 2018, Majolo et al studied the Notch signalling pathway, a pathway involved in cortical development to regulate neuronal differentiation, self-renewal, survival, and neuronal plasticity, using the iPSCs from FCD IIb patients. The study assess the expression of genes involved in Notch signalling and showed that, during embryonic neurogenesis, the neural precursor cells of FCD type IIb individuals exhibited an increase in HEY1 and NOTCH1 genes as well as a decrease in the expression of HES1 and PAX5 genes, compared to the cells from control subjects [53][82]. In the subsequent study, Majolo et al studied the migration and synaptic aspects of neurons generated using the iPSCs derived from patients with FCD type IIb. Using real-time PCR, the study presented the expression of most of the synaptic and ion channels genes ASCL1, DCX, DLG4, FGF2, NEFL, NEUROD2, NEUROD6, NRCAM, and STX1A in different groups; fibroblasts, iPSCs, differentiated neurons, and brain tissues [54][83]. This study suggested that the cells derived from FCD patients may have more sensitivity to stimuli resulting in altered cell survival, apoptosis, migration, and morphological development. In 2021, Klofas et al published a study using the FCD patient-derived neurons carrying a heterozygous loss-of-function mutation in the DEPDC5 gene and reported hyperactivation of mTORC1 and enlarged cell somas that were rescued with the inhibition of mTORC1. This study also reported that cell starvation leads to hyperactivation of the mTOR pathway [57][104] but the exact mechanism is still unclear. None of these FCD studies have performed electrophysiological functional analysis of the generated neurons.