Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Dean Liu and Version 1 by Michael Fossel.
Interventions aimed at the cellular origin of the pathology would have a better chance of preventing and treating cardiovascular disease, lowering healthcare costs, and incurring less risk.
- cell senescence
- aging
- cardiovascular disease
- telomerase
- telomere
1. The Role of Cell Aging in Age-Related Cardiovascular Disease
Cell aging has been theorized [53,54,56,60][1][2][3][4] and implicated in age-related disease in animals [61,62][5][6] and humans [50,63][7][8]. The unified systems model (Figure 21) suggests that cell aging mediates the cascade of events between upstream clinical risk factors and downstream clinical outcomes. Traumatic injury may accelerate cell aging in joints, UV exposure may accelerate cell aging in the skin, and having two APOE4 alleles may accelerate microaggregate formation, even in the context of relatively mild microglial cell aging, resulting in earlier and more severe Alzheimer’s disease. The downstream heterogeneity of clinical outcomes (specific diagnosis, age of onset, course of disease, individual clinical findings, etc.) results from the upstream heterogeneity of genetic, epigenetic, and behavioral history. However, pathology plays out in the unified “stage” of cell aging (Figure 21).
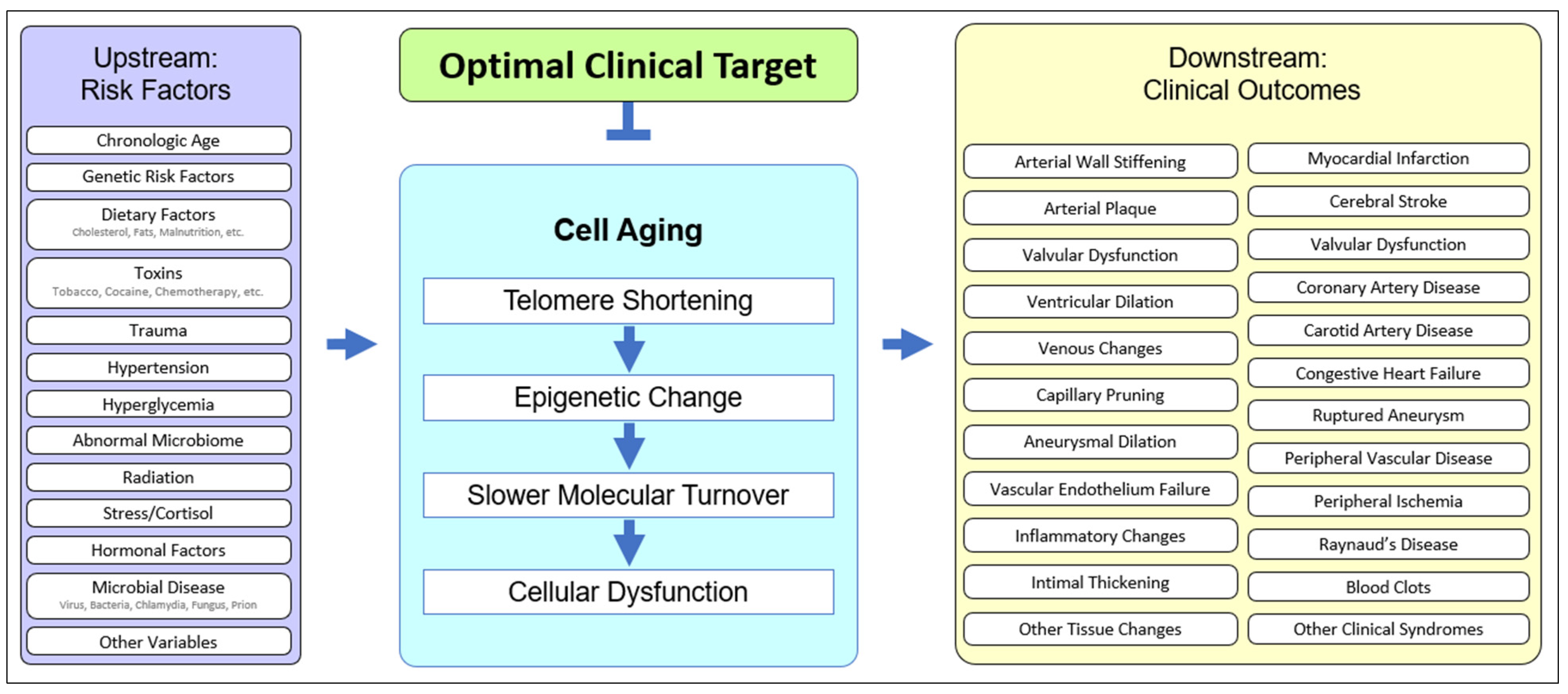
Figure 21. The unified model of cell aging being central to age-related cardiovascular disease. Upstream risk factors provide variable starting points, affect cell aging, and result in variable downstream clinical outcomes, suggesting an optimal point of intervention.
Cell aging is typified by a deceleration in molecular turnover, whether intracellular, extracellular, or intranuclear, and whether protein, lipid, or other molecular moieties, resulting in a gradual increase in the percentage of dysfunctional molecules. Slower molecular turnover results in an increased proportion of damage, but the rate of damage can also increase as a result of genetic or environmental effects acting synergistically with cell aging. Upstream risk factors can not only increase the rate of cell aging [64,65,66,67][9][10][11][12] but can also exacerbate the effects of cell aging. For example, rheologic trauma (e.g., at bifurcations, the aortic arch, etc.) and barotrauma (e.g., hypertension) may increase the rate of cell aging and thereby hasten the onset of pathology (e.g., aortic aneurysm), but genetic risks are also operative [68,69,70,71][13][14][15][16].
Cardiovascular disease is the result of vascular endothelial cell aging [72,73][17][18], as well as aging in smooth muscle cells [74[19][20],75], cardiomyocytes [76][21], cardiac fibroblasts [77][22], and immune cells [78][23]. In aging endothelial cells, there is a decreased endothelial nitric oxide synthase (eNOS) and nitric oxide, critical in atherogenesis and hypertension. Endothelial cell aging increases monocyte adhesion, implicated in atherogenesis, and the induction of endothelial cell aging results in typical atherogenic changes. Importantly, these processes are linked with telomerase activity. In the case of laminar fluid stress, telomerase is required for normal endothelial cell response [79][24], while telomere shortening is associated with increasing arterial stiffness [80][25]. This vascular dysfunction is ameliorated or normalized by telomerase, causing reversion to a pattern of gene expression typical of younger endothelial cells [81[26][27],82], indicting endothelial cell aging in atherogenesis [83][28]. Upstream risk factors leading to cell aging and downstream outcomes predisposing to cardiovascular disease are important to integrate into the Unified Model.
2. Upstream Risk Factors in Cardiovascular Disease
Upstream risk factors are clinical starting points that operate within the context of cell aging and cause age-related disease. Pathology can result from normal cell aging, from accelerated cell aging (e.g., hypertension, diabetes, vascular infections, tobacco use, poor diet, etc.), or from accelerated molecular damage (e.g., genetically abnormal free radical scavengers, membrane lipids, mitochondrial enzymes, etc.). Upstream risks may be categorized as genetic, epigenetic, or behavioral.
Genetic risks may be relatively asymptomatic in early life, as in patients with aberrant elastin or collagen genes [84][29], but increasingly symptomatic in later life as molecular pools are recycled far more slowly and the percentage of damaged molecules rises. Risks may be genetic, as with abnormal cytochrome C oxidase or mitochondrial genes [85,86][30][31], but interactive processes may also occur, for example in endothelial cell aging [67,87,88][12][32][33] and aortic smooth muscle cells [89][34], where oxidative damage [66][11] and cell aging work in tandem [90][35]. Epigenetic risks may include regulatory elements (which greatly outnumber protein-expressing genes), including variability in inherited telomere lengths [91][36]. Epigenetic differences manifest as regulatory idiosyncrasies: patients with identical genes may have different patterns of expression and resulting differences in age-related disease.
Telomere length varies among newborns, while it is similar in different organs of the human fetus [92][37]. Genetic mechanisms involved in the regulation of TERT expression and telomerase activity are described in recent reviews [93][38]. Germline mutations in TERT and other genes that control chromosome termini result in the shortening of telomeres transmitted to the progeny. This leads to progressive telomere length changes over successive generations. Somatic mutations in telomerase machinery, as well as the epigenetic downregulation of TERT and its co-activators, account for tissue-specific differences in telomere attrition and senescence onset during aging. Signaling pathways including WNT, converging on KLF4, as well as c-MYC, GA-binding proteins (GABP), and other E26 transformation-specific (ETS) family transcription factors have been found to regulate TERT transcription [94][39]. Epigenetic mechanisms also include promoter methylation, histone deacetylase inhibitors, and miRNAs [95][40]. The post-translational regulation of TERT activity is also important to consider [96][41]. The age of telomerase inactivation predetermines the rate of telomere attrition, which varies among individuals [91][36]. In combination with somatic genetic and epigenetic changes in the oxidative damage protection pathways, these variables in turn predetermine senescence onset in distinct cell populations and the resulting aging of individual tissues.
Behavioral risks (e.g., smoking [97,98,99][42][43][44], air pollution [11[45][46][47][48],100,101,102], diet [103][49], alcohol, etc.) may either increase the rate of cell aging or exacerbate the effects of cell aging. An example from the central nervous system may clarify. Head trauma or CNS infections would increase the risk of Alzheimer’s disease by increasing the rate of cell aging, while a patient with no history of trauma or infection but who is biallelic for APOE4 is prone to earlier microaggregate formation, exacerbating the effects of cell aging. There is a growing body of evidence that the rate of telomere shortening is also a function of environment and behavior.
Cell aging is universal, while the risk, timing, and severity of the resulting age-related cardiovascular diseases are defined by genetic, epigenetic, and behavioral starting points. Specific downstream clinical outcomes—myocardial infarction, stroke, varicosities, cardiomyopathy, etc.—depend on the specific upstream starting points.
3. Downstream Outcomes in Cardiovascular Disease
Downstream outcomes—biomarkers, clinical syndromes, or pathology—vary because upstream risk factors vary, even in the face of a unified fundamental process, that of cell aging. In atherosclerosis, for example, the initial formation of a fatty streak, composed of lipid-laden macrophages (foam cells) within the intima, is followed by vascular smooth muscle cell proliferation, fibrous plaques, cholesterol deposits, and inflammatory cell recruitment [104][50]. The observable fatty streak formation is, however, preceded by a less obvious inflammatory response within the endothelium, which has been linked to a number of insults, including hypertension, diabetes, tobacco use, and the inflammatory secretory pattern directly associated with cell aging [105][51]. This results in the recruitment of inflammatory cells (e.g., monocytes) and the loss of adhesion in endothelial cell junctions. The failure of cell junction integrity causes the leakage of plasma components, including LDL cholesterol, into the subendothelial compartment. Inflammation further contributes to endothelial damage and predisposes to LDL phagocytosis. Recruited monocytes transform into macrophages, phagocytosing oxidized LDL and forming atherosclerotic foam cells [104][50]. They also secrete inflammatory factors, induce smooth muscle cell proliferation and often form fibrotic tissue.
The earliest fatty streaks in children and adolescents lack this fibrous component and other pathological features that characterize atherosclerosis in elderly patients where cell aging is prominent [105,106,107][51][52][53]. Pathology can occur without cell senescence, for example in the presence of significant physical or oxidative damage to the endothelium, as evidenced by arterial fatty streaks in many patients by age twenty. However, the arterial remodeling typical of the young [105,106,107,108][51][52][53][54] is slower or absent as cell aging progresses and other changes, such as mitochondrial dysfunction, occur [76,109,110][21][55][56]. Cell aging drives the progression of atherosclerosis even without classical risk factors [111,112][57][58] and is critical to the progression of significant pathology [107,108][53][54]. Early lesions may regress with effective clinical intervention in upstream risk factors, but advanced lesions, in which cell aging is prominent, are not [106,111,112,113][52][57][58][59]. Even aggressive intervention in risk factors (e.g., diabetes, hypertension, and cholesterol) are insufficient, as the cell aging of the arterial cells previously induced by such factors continues apace [78,83,104,106,111,114,115][23][28][50][52][57][60][61]. The vascular endothelial cell aging continues, and pathology accelerates [78,83,111,114,115][23][28][57][60][61].
Hutchinson-Gilford progeria provides an example of the role of cell aging and telomere shortening in atherosclerosis. In these children, their lamin-A mutation results in telomere lengths typical of 70-year-old patients. Although lacking classical risk factors—diabetes, tobacco use, hypertension, or elevated cholesterol—they develop atherosclerotic pathology [116][62] and usually die (average lifespan 12.7 years) of atherosclerotic disease [117,118][63][64]. Mouse models of H g progeria likewise show an impairment of mechano-signaling in endothelial cells, suggesting that cell aging and their resulting senescence contributes to excessive fibrosis and cardiovascular disease [119][65], and studies on human progeric cells suggest that endothelial-targeted therapy may be effective [120][66]. Cardiovascular disease occurs in the context of accelerated cell aging, even when other risk factors are absent.
Cardiovascular risk factors such as diabetes, hypertension, obesity, and dyslipidemia may be promoted by cell aging and senescence, but may also accelerate cell aging, resulting in a vicious cycle of pathology. Ongoing damage accelerates cell aging, leading to subendothelial exposure, persistent and irreversible inflammation, the progression of atherosclerotic plaque, reduced fibrous plaque coverage, and eventual rupture [78,83,114,115][23][28][60][61]. This is all the more serious with LDL elevation, as high cholesterol leads to increased plaque and the oxidation of LDL within the intima, with increased inflammatory responses [106,111,112,113][52][57][58][59].
A similar causal cascade occurs in aneurysms, as the cell aging of the vascular walls results in structural deterioration. The vascular mesenchymal stromal cells (VMSCs) from aortic aneurysms are more senescent than those from healthy aortas, and medial smooth muscle cells taken from aneurysms are likewise more senescent than cells from normal arteries in the same individual [121,122,123][67][68][69].
Cardiac failure is also linked to cell aging. Cardiomyocytes [48,124][70][71] and cardiac fibroblasts [77][22] display the hallmarks of cell aging, including dysfunctional DNA repair, mitochondrial dysfunction, impaired contractility, and increased fibrosis, with the subsequent impairment of ejection fraction and heart failure [76,124,125,126][21][71][72][73]. Curiously, experimentally induced transient [127][74] or premature senescence [128][75] may reduce perivascular fibrosis and inflammation, perhaps by converting actively dysfunctional (though aged) cells into far less active, replicative senescent cells. Age-related increases in arrhythmias may also be linked to cell aging [129][76].
4. Heterogeneity of Clinical Presentations in Cardiovascular Disease
Age-related cardiovascular disease is clinically heterogenous, encompassing a spectrum of diseases that share a temporal predilection (aging) and a tissue of origin (the cardiovascular system) but often little else, at least superficially. There may appear to be little in common between strokes, myocardial infarctions, varicose veins, and capillary pruning. However, such diseases all demonstrate vascular endothelial cell aging and secondary organ pathology [130][77].
In the case of CNS arterial disease, both hemorrhagic and thrombotic stroke can result from the cell aging of the vascular endothelial cells, the former from arterial rupture and the latter from arterial blockage. In the case of coronary disease, myocardial infarction results from insufficient flow secondary to vascular endothelial cell aging. Other presentations typical of aging patients [131][78]—varicose veins, chronic venous insufficiency, deep vein thrombosis, capillary pruning, etc.—can be the result of vascular endothelial cell aging, with the consequent loss of integrity and structural resilience, whether in arterial, venous, or capillary walls.
Clinical heterogeneity is common. Patients of identical age may vary in disease, vascular localization, onset, or course, and this heterogeneity likely results from the heterogeneity of the initial risk factors. Individuals differ genetically, epigenetically, and behaviorally. Genetic differences may include alleles which impact cholesterol metabolism, inflammatory processes, immune responses, the efficacy of hepatic detoxification, renal physiology, collagen strength, elastin production, or a myriad of other genetic “starting points” prior to the changes incurred by cell aging. Epigenetic differences may be equally profound: rather than an abnormal gene, there is an abnormal pattern of gene expression, with subtly different responses to physiologic stimuli. Behavioral history—alcohol use, diet, exercise, infectious disease, trauma, etc.—also generates clinical heterogeneity in cardiovascular disease.
Genetic risk factors have become increasingly well-established over the past decade with the increased use of genome-wide association studies (GWAS) to identify disease-susceptibility loci [132,133,134][79][80][81]. However, epigenetic diagnostic markers and therapeutic targets, independent of genetic inheritance, have also been identified [135,136,137][82][83][84]. While associations between the risk factors and the clinical outcomes are commonly acknowledged, the cascade of events proceeding via cell aging, presenting as overt changes in tissue and organ function, and ultimately as clinical disease, are less well appreciated.
Contributing to the confusion, many studies analyze aging in cells that are not responsible for the disease [138,139][85][86]. Because of sample collection simplicity, circulating leukocytes rather than vascular endothelial cells are commonly analyzed for telomere attrition [140,141,142,143,144,145,146,147,148,149][87][88][89][90][91][92][93][94][95][96]. As might be expected, there is a rough correlation in telomere lengths between different tissues since the entire organism is undergoing progressive cell aging over time. However, the same might be said of the correlation between cardiovascular disease, CNS neurodegenerative disease, osteoarthritis, osteoporosis, and other age-related diseases as the patient ages. The correlation reflects the age of the patient but is not sufficiently specific to the tissue in question. Although too often measured in patients with cholesterol abnormalities [150][97] or cardiovascular disease [151,152,153,154[98][99][100][101][102][103],155,156], leukocyte telomere lengths can change for independent reasons, such as infections or autoimmune reactions. Relying upon leukocyte measures in assessing cardiovascular disease results in unwarranted conclusions based on inappropriate correlations and irrelevant cell choices. In assessing risk factors for age-related cardiovascular disease, such as diabetes [157,158][104][105] or vascular aging [159][106] wresearche rs see the same error in measuring telomeres in leukocytes [160,161][107][108] rather than in relevant cells. Because vascular dysfunction in cardiovascular disease is largely attributable to endothelial cells [90][35] and/or vascular smooth muscle cells [75][20], telomere lengths in such cells should be used as a gauge of cell aging and atherosclerosis [162][109]. Studies that focus on the measurement of telomeres and cell aging of the appropriate cells find a clearer picture [163,164,165][110][111][112]. The data show (and reviews have emphasized) that telomere shortening and cell aging are not only implicated in age-related cardiovascular aging but precede and are more prominent in affected tissues [166,167][113][114]. These studies underline the role of cell aging in multiple age-related cardiovascular diseases [168][115], including hypertension [169[116][117],170], and atherosclerosis [83[28][118][119],171,172], predisposing to heart failure.
Identifying cell aging as a fundamental mechanism underlying cardiovascular disease offers a more complex—but perhaps a more accurate—paradigm of disease etiology. Current diagnostic approaches to cardiovascular disease test for either upstream risk factors or downstream biomarkers. High cholesterol [150,173][97][120] or hypertension, for example, are clearly upstream risk factors but may operate through the acceleration of cell aging [90,174,175,176][35][121][122][123]. Coronary calcium scans [177][124], on the other hand, are a downstream biomarker, but cell aging may be the driving force behind plaque buildup [178][125].
Upstream risk factors such as chronological age, genetic risk, diet, stress, and trauma contribute to cell aging, while downstream biomarkers of cardiovascular disease, such as venous changes, arterial wall hardening, and myocardial infarctions are the clinical consequences of cell aging. Changes in endothelial physiology, including the eNOS/NO and FGF21 pathways, as well as calcium signaling, appear to be detrimental [87][32]. Other metabolic changes, including hypertensive or hyperglycemic injury to these cells [173][120], are also important [179][126]. Cardiovascular cell aging can also cause secondary CNS disease [180][127], whether as an acute insult (e.g., stroke) or as a chronic result of vascular insufficiency and changes in the blood–brain barrier. Indeed, telomere shortening and cell aging are prominent not only in Alzheimer’s but also in vascular dementia and other age-related neurodegenerative diseases [181,182,183][128][129][130].
There is accumulating evidence that vascular endothelium aging underlies age-related cardiovascular pathology [63,178][8][125]. Endothelial aging induced by experimental telomere attrition stimulates metabolic disorders through SASP [110,184][56][131]. Healthy angiogenesis is not an active process in most mature organs, yet endothelial cell division and capillary sprouting underlie normal maintenance in muscle, adipose tissue, and likely the brain. Muscle regeneration relies on angiogenesis, and dysfunctional angiogenesis is implicated in some cardiovascular diseases. Endothelial cell aging is also likely to underlie peripheral artery disease (PAD) and capillary pruning. Although cause-and-effect may be complicated [185,186[132][133][134][135],187,188], cell aging offers a novel interventional target in both PAD and type-2 diabetes (T2D) [189,190][136][137]. While most cardiac diseases may be attributable to vascular endothelial cell aging, other cell types are also implicated. Cardiomyopathy may be the result of cardiac fibroblast aging [191][138]. Pericytes, mural cells [192][139], macrophages [193][140], and other myeloid cells (with foam cell and plaque formation) may also be involved [194][141]. Mice lacking TERT in perivascular progenitor cells [195][142] have premature metabolic disease [196][143], more so if given a high-caloric diet [197][144].
Adipose tissue, a key reservoir of senescent cells, may indirectly contribute to the aging of cardiovascular cells [198,199,200,201][145][146][147][148]. In obesity, the death of hypertrophic and hypoxic adipocytes prompts leukocyte infiltration, inflammation, fibrosis, and reduced ability to contain triglycerides, which results in steatosis and may contribute to the development of T2D [202][149]. Early in life, adipocyte progenitor cells divide continuously [203[150][151],204], replacing dysfunctional hypertrophic adipocytes [205,206,207][152][153][154]. With age, adipocyte progenitors lose replicative capacity, which is accelerated by hyperlipidemia [150[97][155][156],208,209], obesity [210,211[157][158][159][160],212,213], and oxidative stress [214,215][161][162]. Telomere length in progenitor cells defines the ability of adipose tissue to expand by hyperplasia, rather than hypertrophy [197][144], perhaps explaining the lack of T2D in “healthy obese” individuals [196][143]. In mice lacking TERT, adipocyte progenitors undergo premature replicative senescence, and T2D development is accelerated [195][142]. Obesity development also involves increased endothelial cell division (as well as leukocyte infiltration) to vascularize expanding adipose tissue [196][143]. The accelerated cell aging may aggravate adipose dysfunction, the accompanying dyslipidemia [216][163], and hypertension [217][164], independently of the cardiovascular risk factors [218][165]. All of these aging cells secrete SASP factors with systemic vascular effects. Aging monocytes from adipose tissue may also disseminate systemically and may contribute to atherosclerosis and inflammation directly. Potential long-distance effects of other types of aging cells in adipose tissue remain to be investigated [219][166].
References
- Fossel, M. Telomerase and the aging cell: Implications for human health. JAMA 1998, 279, 1732–1735.
- Banks, D.; Fossel, M. Telomeres, cancer, and aging: Altering the human lifespan. JAMA 1997, 278, 1345–1348.
- Fossel, M. Cells, Aging, and Human Disease; Oxford University Press: Oxford, UK, 2004.
- Olivieri, F.; Recchioni, R.; Marcheselli, F.; Marie Abbatecola, A.; Santini, G.; Borghetti, G.; Antonicelli, R.; Domenico Procopio, A. Cellular Senescence in Cardiovascular Diseases: Potential Age-Related Mechanisms and Implications for Treatment. Curr. Pharm. Des. 2013, 19, 1710–1719.
- Jaskelioff, M.; Muller, F.L.; Paik, J.H.; Thomas, E.; Jiang, S.; Adams, A.; Sahin, E.; Kost-Alimova, M.; Protopopov, A.; Cadiñanos, J.; et al. Telomerase reactivation reverses tissue degeneration in aged telomerase deficient mice. Nature 2011, 469, 102–106.
- de Jesus, B.B.; Schneeberger, K.; Vera, M.E.; Tejera, A.; Harley, C.B.; Blasco, M.A. The telomerase activator TA-65 elongates short telomeres and increases health span of adult old mice without increasing cancer incidence. Aging Cell 2011, 10, 604–621.
- Fossel, M. A Unified Model of Dementias and Age-Related Neurodegeneration. Alzheimer’s Dement. 2020, 16, 365–383.
- Dominic, A.; Banerjee, P.; Hamilton, D.J.; Le, N.T.; Abe, J. Time-dependent replicative senescence vs. disturbed flow-induced pre-mature aging in atherosclerosis. Redox Biol. 2020, 37, 101614.
- Hemanthakumar, K.A.; Fang, S.; Anisimov, A.; Mäyränpää, M.I.; Mervaala, E.; Kivelä, R. Cardiovascular disease risk factors induce mesenchymal features and senescence in mouse cardiac endothelial cells. eLife 2021, 10, e62678.
- Liu, Y.; Bloom, S.I.; Donato, A.J. The role of senescence, telomere dysfunction and shelterin in vascular aging. Microcirculation 2018, 26, e12487.
- Imanishi, T.; Hano, T.; Sawamura, T.; Nishio, I. Oxidized low-density lipoprotein induces endothelial progenitor cell senescence, leading to cellular dysfunction. Clin. Exp. Pharmacol. Physiol. 2004, 31, 407–413.
- Kurz, D.J.; Decary, S.; Hong, Y.; Trivier, E.; Akhmedov, A.; Erusalimsky, J.D. Chronic oxidative stress compromises telomere integrity and accelerates the onset of senescence in human endothelial cells. J. Cell Sci. 2004, 117, 2417–2426.
- Erdmann, J.; Linsel-Nitschke, P.; Schunkert, H. Genetic Causes of Myocardial Infarction. Dtsch. Arztebl. Int. 2010, 107, 694–699.
- Dai, X.; Wiernek, S.; Evans, J.P.; Runge, M.S. Genetics of coronary artery disease and myocardial infarction. World J. Cardiol. 2016, 8, 1–23.
- Katz, D.H.; Tahir, U.A.; Bick, A.G.; Pampana, A.; Ngo, D.; Benson, M.D.; Yu, Z.; Robbins, J.M.; Chen, Z.-Z.; Cruz, D.E.; et al. Whole Genome Sequence Analysis of the Plasma Proteome in Black Adults Provides Novel Insights Into Cardiovascular Disease. Circulation 2022, 145, 357–370.
- Yun, H.; Noh, N.I.; Lee, E.Y. Genetic risk scores used in cardiovascular disease prediction models: A systematic review. Rev. Cardiovasc. Med. 2022, 23, 8.
- Yegorov, Y.; Poznyak, A.V.; Nikiforov, N.G.; Starodubova, A.V.; Orekhov, A.N. Role of Telomeres Shortening in Atherogenesis: An Overview. Cells 2021, 10, 395.
- Bloom, S.I.; Islam, M.T.; Lesniewski, L.A.; Donato, A.J. Mechanisms and consequences of endothelial cell senescence. Nat. Rev. Cardiol. 2022, 1–4.
- Gorenne, I.; Kavurma, M.; Scott, S.; Bennett, M. Vascular smooth muscle cell senescence in atherosclerosis. Cardiovasc. Res. 2006, 72, 9–17.
- Matthews, C.; Gorenne, I.; Scott, S.; Figg, N.; Kirkpatrick, P.; Ritchie, A.; Goddard, M.; Bennett, M. Vascular Smooth Muscle Cells Undergo Telomere-Based Senescence in Human Atherosclerosis. Circ. Res. 2006, 99, 156–164.
- Moslehi, J.; DePinho, R.A.; Sahin, E. Telomeres and Mitochondria in the Aging Heart. Circ. Res. 2012, 110, 1226–1237.
- Travers, J.G.; Kamal, F.A.; Robbins, J.; Yutzey, K.E.; Blaxall, B.C. Cardiac Fibrosis: The Fibroblast Awakens. Circ. Res. 2016, 118, 1021–1040.
- Wu, C.-M.; Zheng, L.; Wang, Q.; Hu, Y.-W. The emerging role of cell senescence in atherosclerosis. Clin. Chem. Lab. Med. 2020, 59, 27–38.
- Kant, S.; Tran, K.V.; Kvandova, M.; Caliz, A.D.; Yoo, H.J.; Learnard, H.; Dolan, A.C.; Craige, S.M.; Hall, J.D.; Jiménez, J.M.; et al. PGC1α Regulates the Endothelial Response to Fluid Shear Stress via Telomerase Reverse Transcriptase Control of Heme Oxygenase-1. Arterioscler. Thromb. Vasc. Biol. 2021, 42, 19–34.
- Wai, K.M.; Kaori, S.; Itoh, K.; Shinya, O.; Uchikawa, Y.; Hayashi, S.; Shiraki, A.; Murashita, K.; Nakaji, S.; Ihara, K. Telomere Length and Arterial Stiffness Reflected by Brachial-Ankle Pulse Wave Velocity: A Population-Based Cross-Sectional Study. J. Pers. Med. 2021, 11, 1278.
- Matsushita, H.; Chang, E.; Glassford, A.J.; Cooke, J.P.; Chiu, C.P.; Tsao, P.S. eNOS Activity Is Reduced in Senescent Human Endothelial Cells. Preservation by hTERT Immortalization. Circ. Res. 2001, 89, 793–798.
- Mojiri, A.; Walther, B.K.; Jiang, C.; Matrone, G.; Holgate, R.; Xu, Q.; Morales, E.; Wang, G.; Gu, J.; Wang, R.; et al. Telomerase therapy reverses vascular senescence and extends lifespan in progeria mice. Eur. Heart J. 2021, 42, 4352–4369.
- Minamino, T.; Miyauchi, H.; Yoshida, T.; Ishida, Y.; Yoshida, H.; Komuro, I. Endothelial Cell Senescence in Human Atherosclerosis. Role of Telomere in Endothelial Dysfunction. Circulation 2002, 105, 1541–1544.
- Zekavat, S.M.; Chou, E.L.; Zekavat, M.; Pampana, A.; Paruchuri, K.; Lino Cardenas, C.L.; Koyama, S.; Ghazzawi, Y.; Kii, E.; Uddin, M.M.; et al. Fibrillar Collagen Variants in Spontaneous Coronary Artery Dissection. JAMA Cardiol. 2022, 7, 396–406.
- Behjati, M.; Sabri, M.R.; Far, M.E.; Nejati, M. Cardiac complications in inherited mitochondrial diseases. Heart Fail. Rev. 2020, 26, 391–403.
- Brischigliaro, M.; Zeviani, M. Cytochrome c oxidase deficiency. Biochim. Biophys. Acta (BBA) Bioenerg. 2021, 1862, 148335.
- Jia, G.; Aroor, A.R.; Jia, C.; Sowers, J.R. Endothelial cell senescence in aging-related vascular dysfunction. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1802–1809.
- Bloom, S.; Tucker, J.; Machin, D.; Liu, Y.; Thomas, T.; Abdeahad, H.; Bramwell, R.; Lesniewski, L.; Donato, A. Aging results in endothelial cell telomere uncapping that induces senescence and physiological dysfunction. FASEB J. 2022, 36 (Suppl. 1).
- Gordon, C.A.; Madamanchi, N.R.; Runge, M.S.; Jarstfer, M.B. Effect of oxidative stress on telomere maintenance in aortic smooth muscle cells. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166397.
- Voghel, G.; Thorin-Trescasesa, N.; Farhata, N.; Nguyen, A.; Villeneuvea, L.; Mamarbachi, A.M.; Fortier, A.; Perrault, L.P.; Carrier, N.; Thorin, E. Cellular senescence in endothelial cells from atherosclerotic patients is accelerated by oxidative stress associated with cardiovascular risk factors. Mech. Ageing Dev. 2007, 128, 662–671.
- Martens, D.S.; Van Der Stukken, C.; Derom, C.; Thiery, E.; Bijnens, E.M.; Nawrot, T.S. Newborn telomere length predicts later life telomere length: Tracking telomere length from birth to child- and adulthood. EBioMedicine 2021, 63, 103164.
- Okuda, K.; Bardeguez, A.; Gardner, J.P.; Rodriguez, P.; Ganesh, V.; Kimura, M.; Skurnick, J.; Awad, G.; Aviv, A. Telomere length in the newborn. Pediatr. Res. 2002, 52, 377–381.
- Lister-Shimauchi, E.H.; McCarthy, B.; Lippincott, M.; Ahmed, S. Genetic and Epigenetic Inheritance at Telomeres. Epigenomes 2022, 6, 9.
- Chakravarti, D.; LaBella, K.A.; DePinho, R.A. Telomeres: History, health, and hallmarks of aging. Cell 2021, 184, 306–322.
- Dogan, F.; Forsyth, N.R. Telomerase Regulation: A Role for Epigenetics. Cancers 2021, 13, 1213.
- Jie, M.M.; Chang, X.; Zeng, S.; Liu, C.; Liao, G.B.; Wu, Y.R.; Liu, C.H.; Hu, C.J.; Yang, S.M.; Li, X.Z. Diverse regulatory manners of human telomerase reverse transcriptase. Cell Commun. Signal. 2019, 17, 63.
- Cardenas, A.; Ecker, S.; Fadadu, R.P.; Huen, K.; Orozco, A.; McEwen, L.M.; Engelbrecht, H.R.; Gladish, N.; Kobor, M.S.; Rosero-Bixby, L.; et al. Epigenome-wide association study and epigenetic age acceleration associated with cigarette smoking among Costa Rican adults. Sci. Rep. 2022, 12, 4277.
- Nyunoya, T.; Monick, M.M.; Klingelhutz, A.; Yarovinsky, T.O.; Cagley, J.R.; Hunninghake, G.W. Cigarette Smoke Induces Cellular Senescence. Am. J. Respir. Cell Mol. Biol. 2006, 35, 681–688.
- Farhat, N.; Thorin-Trescases, N.; Voghel, G.; Villeneuve, L.; Mamarbachi, M.; Perrault, L.P.; Carrier, M.; Thorin, E. Stress-induced senescence predominates in endothelial cells isolated from atherosclerotic chronic smokers. Can. J. Physiol. Pharmacol. 2008, 86, 761–769.
- Yusuf, S.; Joseph, P.; Rangarajan, S.; Islam, S.; Mente, A.; Hystad, P.; Brauer, M.; Kutty, V.R.; Gupta, R.; Wielgosz, A.; et al. Modifiable risk factors, cardiovascular disease and mortality in 155, 722 individuals from 21 high-, middle-, and low-income countries. Lancet 2020, 395, 795–808.
- Li, R.; Chen, G.; Liu, X.; Pan, M.; Kang, N.; Hou, X.; Liao, W.; Dong, X.; Yuchi, Y.; Mao, Z.; et al. Aging biomarkers: Potential mediators of association between long-term ozone exposure and risk of atherosclerosis. J. Intern. Med. 2022, 292, 512–522.
- Sharma, K.; Lee, H.H.; Gong, D.S.; Park, S.H.; Yi, E.; Schini-Kerth, V.; Oak, M.H. Fine air pollution particles induce endothelial senescence via redox-sensitive activation of local angiotensin system. Environ. Pollut. 2019, 252 Pt A, 317–329.
- Chang-Chiena, J.; Huang, J.L.; Tsai, H.J.; Wang, S.L.; Kuo, M.L.; Yao, T.C. Particulate matter causes telomere shortening and increase in cellular senescence markers in human lung epithelial cells. Ecotoxicol. Environ. Saf. 2021, 222, 112484.
- Leitão, C.; Mignano, A.; Estrela, M.; Fardilha, M.; Figueiras, A.; Roque, F.; Herdeiro, M.T. The Effect of Nutrition on Aging-A Systematic Review Focusing on Aging-Related Biomarkers. Nutrients 2022, 14, 554.
- Xu, S.; Bendeck, M.; Gotlieb, A.I. Vascular pathobiology: Atherosclerosis and large vessel disease. In Cardiovascular Pathology; Academic Press: Cambridge, MA, USA, 2016; pp. 85–124.
- McGill, H.C., Jr.; McMahan, C.A.; Zieske, A.W.; Tracy, R.E.; Malcom, G.T.; Herderick, E.E.; Strong, J.P. Association of Coronary Heart Disease Risk Factors with microscopic qualities of coronary atherosclerosis in youth. Circulation 2000, 102, 374–379.
- Schade, D.S.; Gonzales, K.; Eaton, R.P. Stop Stenting; Start Reversing Atherosclerosis. Am. J. Med. 2021, 134, 301–303.
- Tuzcu, E.M.; Kapadia, S.R.; Tutar, E.; Ziada, K.M.; Hobbs, R.E.; McCarthy, P.M.; Young, J.B.; Nissen, S.E. High prevalence of coronary atherosclerosis in asymptomatic teenagers and young adults: Evidence from intravascular ultrasound. Circulation 2001, 103, 2705–2710.
- Li, Y.; Gilbert, T.R.; Matsumoto, A.H.; Shi, W. Effect of aging on fatty streak formation in a diet-induced mouse model of atherosclerosis. J. Vasc. Res. 2008, 45, 205–210.
- Vecoli, C.; Borghini, A.; Andreassi, M.G. The Molecular Biomarkers of Vascular Aging and Atherosclerosis: Telomere Length and Mitochondrial DNA 4977 Common Deletion. Mutat. Res. 2020, 784, 108309.
- Ritov, V.B.; Menshikova, E.V.; He, J.; Ferrell, R.E.; Goodpaster, B.H.; Kelley, D.E. Deficiency of subsarcolemmal mitochondria in obesity and type 2 diabetes. Diabetes 2005, 54, 8–14.
- Björkegren, J.L.M.; Hägg, S.; Talukdar, H.A.; Asl, H.F.; Jain, R.K.; Cedergren, C.; Shang, M.-M.; Rossignoli, A.; Takolander, R.; Melander, O.; et al. Plasma cholesterol-induced lesion networks activated before regression of early, mature, and advanced atherosclerosis. PLoS Genet. 2014, 10, e1004201.
- Parsons, C.; Agasthi, P.; Mookadam, F.; Arsanjani, R. Reversal of coronary atherosclerosis: Role of life style and medical management. Trends Cardiovasc. Med. 2018, 28, 524–531.
- Nissen, S.E.; Tuzcu, E.M.; Schoenhagen, P.; Brown, B.G.; Ganz, P.; Vogel, R.A.; Crowe, T.; Howard, G.; Cooper, C.J.; Brodie, B.; et al. Effect of intensive compared with moderate lipid-lowering therapy on progression of coronary atherosclerosis: A randomized controlled trial. JAMA 2004, 291, 1071–1080.
- Wang, J.; Uryga, A.K.; Reinhold, J.; Figg, N.; Baker, L.; Finigan, A.J.; Gray, K.; Kumar, S.; Clarke, M.; Bennett, M.R. Vascular Smooth Muscle Cell Senescence Promotes Atherosclerosis and Features of Plaque Vulnerability. Circulation 2015, 132, 1909–1919.
- Honda, S.; Ikeda, K.; Urata, R.; Yamazaki, E.; Emoto, N.; Matoba, S. Cellular senescence promotes endothelial activation through epigenetic alteration, and consequently accelerates atherosclerosis. Sci. Rep. 2021, 11, 14608.
- Olive, M.; Harten, I.; Mitchell, R.; Beers, J.K.; Djabali, K.; Cao, K.; Erdos, M.R.; Blair, C.; Funke, B.; Smoot, L.; et al. Cardiovascular pathology in Hutchinson-Gilford progeria: Correlation with the vascular pathology of aging. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2301–2309.
- Fossel, M. Accelerated Aging: Progeria. In Genetics; Robinson, R., Ed.; Macmillan: New York, NY, USA, 2003.
- Fossel, M. Human Aging and Progeria. J. Ped. Endo. Metab. 2000, 13, 1477–1481.
- Osmanagic-Myers, S.; Kiss, A.; Manakanatas, C.; Hamza, O.; Sedlmayer, F.; Szabo, P.L.; Fischer, I.; Fichtinger, P.; Podesser, B.K.; Eriksson, M.; et al. Endothelial progerin expression causes cardiovascular pathology through an impaired mechanoresponse. J. Clin. Investig. 2019, 129, 531–545.
- Xu, Q.; Mojiri, A.; Boulahouache, L.; Morales, E.; Walther, B.K.; Cooke, J.P. Vascular senescence in progeria: Role of endothelial dysfunction. Eur. Heart J. Open 2022, 2, oeac047.
- Teti, G.; Chiarini, F.; Mazzotti, E.; Ruggeri, A.; Carano, F.; Falconi, M. Cellular senescence in vascular wall mesenchymal stromal cells, a possible contribution to the development of aortic aneurysm. Mech. Ageing Dev. 2021, 197, 111515.
- Shimizu, I.; Minamino, T. Cellular Senescence in Arterial Diseases. J. Lipid Atheroscler. 2020, 9, 79–91.
- Liao, S.; Curci, J.A.; Kelley, B.J.; Sicard, G.A.; Thompson, R.W. Accelerated replicative senescence of medial smooth muscle cells derived from abdominal aortic aneurysms compared to the adjacent inferior mesenteric artery. J. Surg Res. 2000, 92, 85–95.
- Anderson, R.; Lagnado, A.; Maggiorani, D.; Walaszczyk, A.; Dookun, E.; Chapman, J.; Birch, J.; Salmonowicz, H.; Ogrodnik, M.; Jurk, D.; et al. Length-independent telomere damage drives postmitotic cardiomyocyte senescence. EMBO J. 2019, 38, e100492.
- Tang, X.; Li, P.H.; Chen, H.Z. Cardiomyocyte Senescence and Cellular Communications Within Myocardial Microenvironments. Front. Endocrinol. 2020, 11, 280.
- Mehdizadeh, M.; Aguilar, M.; Thorin, E.; Ferbeyre, G.; Nattel, S. The role of cellular senescence in cardiac disease: Basic biology and clinical relevance. Nat. Rev. Cardiol. 2022, 19, 250–264.
- Gevaert, A.B.; Shakeri, H.; Leloup, A.J.; Van Hove, C.E.; De Meyer, G.R.; Vrints, C.J.; Lemmens, K.; Van Craenenbroeck, E.M. Endothelial Senescence Contributes to Heart Failure With Preserved Ejection Fraction in an Aging Mouse Model. Circ. Heart Fail. 2017, 10, e003806.
- Sarig, R.; Rimmer, R.; Bassat, E.; Zhang, L.; Umansky, K.B.; Lendengolts, D.; Perlmoter, G.; Yaniv, K.; Tzahor, E. Transient p53-Mediated Regenerative Senescence in the Injured Heart. Circulation 2019, 139, 2491–2494.
- Meyer, K.; Hodwin, B.; Ramanujam, D.; Engelhardt, S.; Sarikas, A. Essential Role for Premature Senescence of Myofibroblasts in Myocardial Fibrosis. J. Am. Coll. Cardiol. 2016, 67, 2018–2028.
- Guo, G.; Watterson, S.; Zhang, S.-D.; Bjourson, A.; McGilligan, V.; Peace, A.; Rai, T.S. The role of senescence in the pathogenesis of atrial fibrillation: A target process for health improvement and drug development. Ageing Res. Rev. 2021, 69, 101363.
- Ferrari, S.; Pesce, M. Stiffness and Aging in Cardiovascular Diseases: The Dangerous Relationship between Force and Senescence. Int. J. Mol. Sci. 2021, 22, 3404.
- Molnár, A.A.; Nádasy, G.L.; Dörnyei, G.; Patai, B.B.; Delfavero, J.; Fülöp, G.A.; Kirkpatrick, A.C.; Ungvári, Z.; Merkely, B. The aging venous system: From varicosities to vascular cognitive impairment. GeroScience 2021, 43, 2761–2784.
- Vilne, B.; Schunkert, H. Integrating Genes Affecting Coronary Artery Disease in Functional Networks by Multi-OMICs Approach. Front. Cardiovasc. Med. 2018, 5, 89.
- McPherson, R.; Tybjaerg-Hansen, A. Genetics of Coronary Artery Disease. Circ. Res. 2016, 118, 564–578.
- Chen, Z.; Schunkert, H. Genetics of coronary artery disease in the post-GWAS era. J. Int. Med. 2021, 290, 980–992.
- Sumi, M.P.; Mahajan, B.; Sattar, R.S.A.; Nimisha; Apurva; Kumar, A.; Sharma, A.K.; Ahmad, E.; Ali, A.; Saluja, S.S. Elucidation of Epigenetic Landscape in Coronary Artery Disease: A Review on Basic Concept to Personalized Medicine. Epigenetic Insights 2021, 14.
- Dogan, M.V.; Knight, S.; Dogan, T.K.; Knowlton, K.U.; Philibert, R. External validation of integrated genetic-epigenetic biomarkers for predicting incident coronary heart disease. Epigenomics 2021, 13, 1095–1112.
- Garratt, H.; Ashburn, R.; Sopic, M.; Nogara, A.; Caporali, A.; Mitic, T. Long Non-Coding RNA Regulation of Epigenetics in Vascular Cells. Non-Coding RNA 2021, 7, 62.
- Fossel, M. Use of Telomere Length as a Biomarker for Aging and Age-Related Disease. Curr. Transl. Geriatr. Exp. Gerontol. Rep. 2012, 1, 121–127.
- Semeraro, M.D.; Almer, G.; Renner, W.; Gruber, H.J.; Herrmann, M. Telomere length in leucocytes and solid tissues of young and aged rats. Aging 2022, 14, 1713–1728.
- Bhattacharyya, J.; Mihara, K.; Bhattacharjee, D.; Mukherjee, M. Telomere length as a potential biomarker of coronary artery disease. Indian J. Med. Res. 2017, 145, 730–737.
- Schneider, C.V.; Schneider, K.M.; Teumer, A.; Rudolph, K.L.; Hartmann, D.; Rader, D.J.; Strnad, P. Association of Telomere Length with Risk of Disease and Mortality. JAMA Intern. Med. 2022, 182, 291.
- De Meyer, T.; Van Daele, C.M.; De Buyzere, M.L.; Denil, S.; De Bacquer, D.; Segers, P.; Cooman, L.; De Backer, G.G.; Gillebert, T.C.; Bekaert, S.; et al. No shorter telomeres in subjects with a family history of cardiovascular disease in the Asklepios study. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 3076–3081.
- Doroschuk, N.A.; Postnov, A.Y.; Doroschuk, A.D.; Ryzhkova, A.I.; Sinyov, V.V.; Sazonova, M.D.; Khotina, V.A.; Orekhov, A.N.; Sobenin, I.A.; Sazonova, M.A. An original biomarker for the risk of developing cardiovascular diseases and their complications: Telomere length. Tox Rep. 2021, 8, 499–504.
- Codd, V.; Denniff, M.; Swinfield, C.; Warner, S.C.; Papakonstantinou, M.; Sheth, S.; Nanus, D.E.; Budgeon, C.A.; Musicha, C.; Bountziouka, V.; et al. Measurement and initial characterization of leukocyte telomere length in 474,074 participants in UK Biobank. Nat. Aging 2022, 2, 170–179.
- Vecoli, C.; Basta, G.; Borghini, A.; Gaggini, M.; Del Turco, S.; Mercuri, A.; Gastaldelli, A.; Andreassi, M.G. Advanced glycation end products, leukocyte telomere length, and mitochondrial DNA copy number in patients with coronary artery disease and alterations of glucose homeostasis: From the GENOCOR study. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 1236–1244.
- Nordfjäll, K.; Eliasson, M.; Stegmayr, B.; Melander, O.; Nilsson, P.; Roos, G. Telomere Length Is Associated with Obesity Parameters but With a Gender Difference. Obesity 2012, 16, 2682–2689.
- Fyhrquist, F.; Saijonmaa, O.; Strandberg, T. The roles of senescence and telomere shortening in cardiovascular disease. Nat. Rev. Cardiol. 2013, 10, 274–283.
- Zimnitskaya, O.V.; Petrova, M.M.; Lareva, N.V.; Cherniaeva, M.S.; Al-Zamil, M.; Ivanova, A.E.; Shnayder, N.A. Leukocyte Telomere Length as a Molecular Biomarker of Coronary Heart Disease. Genes 2022, 13, 1234.
- Aviv, H.; Khan, M.Y.; Skurnick, J.; Okuda, K.; Kimura, M.; Gardner, J.; Priolo, L.; Aviv, A. Age dependent aneuploidy and telomere length of the human vascular endothelium. Atherosclerosis 2001, 159, 281–287.
- Shi, Q.; Hubbard, G.B.; Kushwaha, R.S.; Rainwater, D.; Thomas, C.A.; Leland, M.M.; VandeBerg, J.L.; Wang, X.L. Endothelial senescence after high-cholesterol, high-fat diet challenge in baboons. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H2913–H2920.
- De Meyer, T.; Nawrot, T.; Bekaert, S.; De Buyzere, M.L.; Rietzschel, E.R.; Andrés, V. Telomere Length as Cardiovascular Aging Biomarker. J. Am. Coll. Cardiol. 2018, 72, 805–813.
- Zhang, W.G.; Zhu, S.Y.; Bai, X.J.; Zhao, D.L.; Jiang, S.M.; Li, J.; Li, Z.X.; Fu, B.; Cai, G.Y.; Sun, X.F.; et al. Select aging biomarkers based on telomere length and chronological age to build a biological age equation. Age 2014, 36, 9639.
- Saliha, R.; Syed Tasleem, R.; Farzana, M. Telomere Length Variations in Aging and Age-Related Diseases. Curr. Aging Sci. 2014, 7, 161–167.
- Fasching, C.L. Telomere length measurement as a clinical biomarker of aging and disease. Crit. Rev. Clin. Lab. Sci. 2018, 55, 443.
- Martins, M.J.; Constância, M.; Neves, D.; Simm, A. Biomarkers of Aging: From Cellular Senescence to Age-Associated Diseases. Oxidative Med. Cell. Longev. 2017, 55, 443.
- Bekaert, S.; De Meyer, T.; Rietzschel, E.R.; De Buyzere, M.L.; De Bacquer, D.; Langlois, M.; Segers, P.; Cooman, L.; Van Damme, P.; Cassiman, P.; et al. Telomere length and cardiovascular risk factors in a middle-aged population free of overt cardiovascular disease. Aging Cell 2007, 6, 639–647.
- Opstad, T.B.; Berg, T.J.; Bech Holte, K.B.; Arnesen, H.; Solheim, S.; Seljeflot, I. Reduced leukocyte telomere lengths and sirtuin 1 gene expression in long-term survivors of type 1 diabetes: A Dialong substudy. J. Diabetes Investig. 2020, 12, 1183–1192.
- Ali, M.K.; Narayan, K.M.; Tandon, N. Diabetes & coronary heart disease: Current perspectives. Indian J. Med. Res. 2010, 132, 584–597.
- Kosmopoulos, M.; Chiriacò, M.; Stamatelopoulos, K.; Tsioufis, C.; Masci, P.G.; Kontogiannis, C.; Mengozzi, A.; Pugliese, N.R.; Taddei, S.; Virdis, A.; et al. The relationship between telomere length and putative markers of vascular ageing: A Systematic Review and Meta-analysis. Mech. Ageing Dev. 2021, 201, 111604.
- Willeit, P.; Willeit, J.; Brandstätter, A.; Ehrlenbach, S.; Mayr, A.; Gasperi, A.; Weger, S.; Oberhollenzer, F.; Reindl, M.; Kronenberg, F.; et al. Cellular Aging Reflected by Leukocyte Telomere Length Predicts Advanced Atherosclerosis and Cardiovascular Disease Risk. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1649–1656.
- Pusceddu, I.; Kleber, M.; Delgado, G.; Herrmann, W.; Marz, W.; Herrmann, M. Telomere length and mortality in the Ludwigshafen Risk and Cardiovascular Health study. PLoS ONE 2018, 13, e0198373.
- Sun, X.; Feinberg, M.W. Vascular Endothelial Senescence: Pathobiological Insights, Emerging Long Noncoding RNA Targets, Challenges and Therapeutic Opportunities. Front. Physiol. 2021, 12, 693067.
- Khan, S.; Naidoo, D.P.; Chuturgoon, A.A. Telomeres and atherosclerosis: Review. S. Afr. J. Diabetes Vasc. Dis. 2015, 12. Available online: https://hdl.handle.net/10520/EJC181963 (accessed on 15 October 2022).
- den Buijs, J.O.; Musters, M.; Verrips, T.; Post, J.P.; Braam, B.; van Riel, N. Mathematical modeling of vascular endothelial layer maintenance: The role of endothelial cell division, progenitor cell homing, and telomere shortening. Am. J. Physiol. 2004, 287, H2651–H2658.
- Xie, Y.; Lou, D.; Zhang, D. Melatonin Alleviates Age-Associated Endothelial Injury of Atherosclerosis via Regulating Telomere Function. J. Inflamm. Res. 2021, 14, 6799–6812.
- Zglinicki, T.; Martin-Ruiz, C.M. Telomeres as Biomarkers for Ageing and Age-Related Diseases. Curr. Mol. Med. 2005, 5, 197–203.
- Ogami, M.; Ikura, Y.; Ohsawa, M.; Matsuo, T.; Kayo, S.; Yoshimi, N.; Hai, E.; Shirai, N.; Ehara, S.; Komatsu, R.; et al. Telomere Shortening in Human Coronary Artery Diseases. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 546–550.
- Serrano, A.L.; Andres, V. Telomeres and Cardiovascular Disease. Does Size Matter? Circ. Res. 2004, 94, 575–584.
- Aviv, A.; Aviv, H. Reflections on Telomeres, Growth, Aging, and Essential Hypertension. Hypertension 1997, 29, 1067–1072.
- Aviv, A.; Aviv, H. Telomeres and essential hypertension. Am. J. Hypertens. 1999, 12, 427–432.
- Okuda, K.; Khan, M.Y.; Skurnick, J.; Kimura, M.; Aviv, H.; Aviv, A. Telomere attrition of the human abdominal aorta: Relationships with age and atherosclerosis. Atherosclerosis 2000, 152, 391–398.
- Edo, M.D.; Andres, V. Aging, telomeres, and atherosclerosis. Cardiovascular. Res. 2005, 66, 213–221.
- Maeda, M.; Hayashi, T.; Mizuno, N.; Hattori, Y.; Kuzuya, M. Intermittent High Glucose Implements Stress-Induced Senescence in Human Vascular Endothelial Cells: Role of Superoxide Production by NADPH Oxidase. PLoS ONE 2015, 10, e0123169.
- Roger, I.; Milara, J.; Belhadj, N.; Cortijo, J. Senescence Alterations in Pulmonary Hypertension. Cells 2021, 10, 3456.
- McCarthy, C.G.; Wenceslau, C.F.; Webb, R.C.; Joe, B. Novel Contributors and Mechanisms of Cellular Senescence in Hypertension-Associated Premature Vascular Aging. Am. J. Hypertens. 2019, 32, 709–719.
- Westhoff, J.H.; Hilgers, K.F.; Steinbach, M.P.; Hartner, A.; Klanke, B.; Amann, K.; Melk, A. Hypertension Induces Somatic Cellular Senescence in Rats and Humans by Induction of Cell Cycle Inhibitor p16INK4a. Hypertension 2008, 52, 123–129.
- Bell, K.J.L.; White, S.; Hassan, O.; Zhu, L.; Scott, A.M.; Clark, J.; Glasziou, P. Evaluation of the Incremental Value of a Coronary Artery Calcium Score Beyond Traditional Cardiovascular Risk Assessment: A Systematic Review and Meta-analysis. JAMA Intern. Med. 2022, 182, 634.
- Katsuumi, G.; Shimizu, I.; Yoshida, Y.; Minamino, T. Vascular Senescence in Cardiovascular and Metabolic Diseases. Front. Cardiovasc. Med. 2018, 5, 18.
- Draoui, N.; de Zeeuw, P.; Carmeliet, P. Angiogenesis revisited from a metabolic perspective: Role and therapeutic implications of endothelial cell metabolism. Open Biol. 2017, 7, 170219.
- Graves, S.I.; Baker, D.J. Implicating endothelial cell senescence to dysfunction in the ageing and diseased brain. Basic Clin. Pharmacol. Toxicol. 2020, 127, 102–110.
- Eitan, E.; Hutchison, E.R.; Mattson, M.P. Telomere shortening in neurological disorders: An abundance of unanswered questions. Trends Neurosci. 2014, 37, 256–263.
- Kota, L.N.; Bharath, S.; Purushottam, M.; Moily, N.S.; Sivakumar, P.T.; Varghes, M.; Pal, P.K.; Jain, S. Reduced telomere length in neurodegenerative disorders may suggest shared biology. J. Neuropsychiatry Clin. Neurosci. 2015, 27, e92–e96.
- Liu, M.Y.; Neme, A.; Zhou, Q.G. The Emerging Roles for Telomerase in the Central Nervous System. Front. Mol. Neurosci. 2018, 11, 160.
- Barinda, A.J.; Ikeda, K.; Nugroho, D.B.; Wardhana, D.A.; Sasaki, N.; Honda, S.; Urata, R.; Matoba, S.; Hirata, K.; Emoto, N. Endothelial progeria induces adipose tissue senescence and impairs insulin sensitivity through senescence associated secretory phenotype. Nat. Commun. 2020, 11, 481.
- Regulsk, M. Understanding Diabetic Induction of Cellular Senescence: A Concise Review. Wounds 2018, 30, 96–101.
- Liu, J.; Huang, K.; Cai, G.-Y.; Chen, X.-M.; Yang, J.-R.; Lin, L.-R.; Yang, J.; Huo, B.-G.; Zhan, J.; He, Y.-N. Receptor for advanced glycation end-products promotes premature senescence of proximal tubular epithelial cells via activation of endoplasmic reticulum stress-dependent p21 signaling. Cell Signal. 2014, 26, 110–121.
- Narasimhan, A.; Flores, R.R.; Robbins, P.D.; Niedernhofer, L.J. Role of Cellular Senescence in Type II Diabetes. Endocrinology 2021, 162, bqab136.
- Kuki, S.; Imanishi, T.; Kobayashi, K.; Matsuo, Y.; Obana, M.; Akasaka, T. Hyperglycemia accelerated endothelial progenitor cell senescence via the activation of p38 mitogen-activated protein kinase. Circ. J. 2006, 70, 1076–1081.
- Palmer, A.K.; Tchkonia, T.; LeBrasseur, N.K.; Chini, E.N.; Xu, M.; Kirkland, J.L. Cellular Senescence in Type 2 Diabetes: A Therapeutic Opportunity. Diabetes 2015, 64, 2289–2298.
- Senthil, K.K.J.; Gokila, V.M.; Wang, S.Y. Activation of Nrf2-mediated anti-oxidant genes by antrodin C prevents hyperglycemia-induced senescence and apoptosis in human endothelial cells. Oncotarget 2017, 8, 96568–96587.
- Hu, C.; Zhang, X.; Teng, T.; Ma, Z.G.; Tang, Q.Z. Cellular Senescence in Cardiovascular Diseases: A Systematic Review. Aging Dis. 2022, 13, 103–128.
- Sweeney, M.; Foldes, G. It Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and Disease. Front. Cardiovasc. Med. 2018, 5, 154.
- Behmoaras, J.; Gil, J. Similarities and interplay between senescent cells and macrophages. J. Cell Biol. 2021, 220, e202010162.
- Pamukcu, B.; Lip, G.Y.; Devitt, A.; Griffiths, H.; Shantsila, E. The role of monocytes in atherosclerotic coronary artery disease. Ann. Med. 2010, 42, 394–403.
- Gao, Z.; Daquinag, A.C.; Fussell, C.; Zhao, Z.; Dai, Y.; Rivera, A.; Snyder, B.E.; Eckel-Mahan, K.L.; Kolonin, M.G. Age-associated telomere attrition in adipocyte progenitors predisposes to metabolic disease. Nat. Metab. 2020, 2, 1482–1497.
- Eckel-Mahan, K.; Latre, A.R.; Kolonin, M.G. Adipose Stromal Cell Expansion and Exhaustion: Mechanisms and Consequences. Cells 2020, 9, 863.
- Ribas-Latre, A.; Santos, R.B.; Fekry, B.; Tamim, Y.M.; Shivshankar, S.; Mohamed, A.M.T.; Baumgartner, C.; Kwok, C.; Gebhardt, C.; Rivera, A.; et al. Cellular and physiological circadian mechanisms drive diurnal cell proliferation and expansion of white adipose tissue. Nat. Commun. 2021, 12, 3482.
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; LeBrasseur, N.K.; Childs, B.G.; van de Sluis, B.; Kirkland, J.L.; van Deursen, J.M. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature 2011, 479, 232–236.
- Palmer, A.K.; Xu, M.; Zhu, Y.; Pirtskhalava, T.; Weivoda, M.M.; Hachfeld, C.M.; Prata, L.G.; van Dijk, T.H.; Verkade, E.; Casaclang-Verzosa, G.; et al. Targeting senescent cells alleviates obesity-induced metabolic dysfunction. Aging Cell 2019, 18, e12950.
- Schafer, M.J.; White, T.A.; Evans, G.; Tonne, J.M.; Verzosa, G.C.; Stout, M.B.; Mazula, D.L.; Palmer, A.K.; Baker, D.J.; Jensen, M.D.; et al. Exercise Prevents Diet-Induced Cellular Senescence in Adipose Tissue. Diabetes 2016, 65, 1606–1615.
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nature reviews. Mol. Cell Biol. 2019, 20, 242–258.
- Kim, S.M.; Lun, M.; Wang, M.; Senyo, S.E.; Guillermier, C.; Patwari, P.; Steinhauser, M.L. Loss of white adipose hyperplastic potential is associated with enhanced susceptibility to insulin resistance. Cell Metab. 2014, 20, 1049–1058.
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787.
- Hammarstedt, A.; Gogg, S.; Hedjazifa, S.; Nerstedt, A.; Smith, U. Impaired Adipogenesis and Dysfunctional Adipose Tissue in Human Hypertrophic Obesity. Physiol. Rev. 2018, 98, 1911–1941.
- Berry, R.; Jeffery, E.; Rodeheffe, M.S. Weighing in on adipocyte precursors. Cell Metab. 2014, 19, 18–20.
- Lakowa, N.; Trieu, N.; Flehmig, G.; Lohmann, T.; Schon, M.R.; Dietrich, A.; Zeplin, P.H.; Langer, S.; Stumvoll, M.; Bluher, M.; et al. Telomere length differences between subcutaneous and visceral adipose tissue in humans. Biochem. Biophys. Res. Commun. 2015, 457, 426–432.
- Lee, Y.H.; Petkova, A.P.; Mottillo, E.P.; Granneman, J.G. In vivo identification of bipotential adipocyte progenitors recruited by beta3-adrenoceptor activation and high-fat feeding. Cell Metab. 2012, 15, 480–491.
- Liu, W.Q.; Zhang, Y.Z.; Wu, Y.; Zhang, J.J.; Li, T.B.; Jiang, T.; Xiong, X.-M.; Luo, X.-J.; Ma, Q.-L.; Peng, J. Myeloperoxidase-derived hypochlorous acid promotes ox-LDL-induced senescence of endothelial cells through a mechanism involving β-catenin signaling in hyperlipidemia. Biochem. Biophys. Res. Commun. 2015, 467, 859–865.
- Oh, S.T.; Park, H.; Yoon, H.J.; Yang, S.Y. Long-Term Treatment of Native LDL Induces Senescence of Cultured Human Endothelial Cells. Oxid Med. Cell Longev. 2017, 2017, 6487825.
- Conley, S.M.; Hickson, L.J.; Kellogg, T.A.; McKenzie, T.; Heimbach, J.K.; Taner, T.; Tang, H.; Jordan, K.L.; Saadiq, I.M.; Woollard, J.R.; et al. Human Obesity Induces Dysfunction and Early Senescence in Adipose Tissue-Derived Mesenchymal Stromal/Stem Cells. Front. Cell Dev. Biol. 2020, 8, 197.
- Ogrodnik, M.; Zhu, Y.; Langhi, L.G.; Tchkonia, T.; Krüger, P.; Fielder, E.; Victorelli, S.; Ruswhandi, R.A.; Giorgadze, N.; Pirtskhalava, T.; et al. Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis. Cell Metab. 2019, 29, 1061–1077.
- Rabhi, N.; Desevin, K.; Belkina, A.C.; Tilston-Lunel, A.; Varelas, X.; Layne, M.D.; Farmer, S.R. Obesity-induced senescent macrophages activate a fibrotic transcriptional program in adipocyte progenitors. Life Sci. Alliance 2022, 5, e202101286.
- Ahima, R. Connecting obesity, aging and diabetes. Nat. Med. 2009, 15, 996–997.
- Smith, U.; Li, Q.; Rydén, M.; Spalding, K.L. Cellular senescence and its role in white adipose tissue. Int. J. Obes. 2021, 45, 934–943.
- Burton, D.G.A.; Faragher, R.G.A. Obesity and type-2 diabetes as inducers of premature cellular senescence and ageing. Biogerontology 2018, 19, 447–459.
- Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218–1240.
- Hall, J.E.; do Carmo, J.M.; da Silva, A.A.; Wang, Z.; Hall, M.E. Obesity-Induced Hypertension—Interaction of Neurohumoral and Renal Mechanisms. Circ. Res. 2015, 116, 991–1006.
- Cercato, C.; Fonseca, F.A. Cardiovascular risk and obesity. Diabetol. Metab. Syndr. 2019, 11, 74.
- Murakami, T.; Inagaki, N.; Kondoh, H. Cellular Senescence in Diabetes Mellitus: Distinct Senotherapeutic Strategies for Adipose Tissue and Pancreatic β Cells. Front. Endocrinol 2022, 13, 869414.
More