Secretoneurin (SN) is a 33 amino-acid evolutionary conserved neuropeptide from the chromogranin peptide family. SN’s main effects may be cardioprotective and are believed to be mediated through its inhibition of calmodulin-dependent kinase II (CaMKII), which influences intracellular calcium handling. SN inhibition of CaMKII suppresses calcium leakage from the sarcoplasmic reticulum through the ryanodine receptor. This action may reduce the risk of ventricular arrhythmias and calcium-dependent remodelling in heart failure. SN is also involved in reducing the intracellular reactive oxygen species concentration, modulating the immune response, and regulating the cell cycle, including apoptosis. SN can predict mortality in different disease states, beyond the classical risk factors and markers of myocardial injury. Plasma SN levels are elevated soon after an arrhythmogenic episode. In summary, SN is a novel biomarker with potential in cardiovascular medicine, and probably beyond.
1. Basic Properties of Secretoneurin
S
ecretoneurin (SN
) ranges from 31 (zebrafish) to 43 (lamprey) amino acids in length. SNs are evolutionarily conserved neuropeptides, mostly 33 (human, chicken, and frog) amino acids long, which arose in tetrapods, and they vary in very few positions
[1][5]. All known SN peptides contain the YTPQ-X-LA-X(7)-EL sequence, which constitutes its central core
[1][5]. In addition, SNa and SNb paralogs have been identified in teleosts, and the N-terminus and the middle of teleost SNa are identical to the mammalian sequence
[2][6]. In contrast, teleost SNb, as compared to mammalian SNb, are only moderately conserved
[2][6].
SN belongs to the secretogranin/chromogranin peptide family
[1][2][5,6]. SN is produced by the proteolytic cleavage of a precursor protein, secretogranin II (SgII, also known as chromogranin C, 617 amino acids), which occurs in several tissues, including cardiomyocytes and neuroendocrine tissues
[3][7]. SgII processing is known to be species- and tissue-specific; thus, sometimes larger SN immunoreactive fragments are generated, in addition to free SN. Most studies have supported the notion that the proprotein convertase subtilisin kexin-1 plays a central role in the generation of SN from SgII
[4][8]. Exact sequence analysis of SgII can be found at
https://www.uniprot.org/uniprotkb/P13521/entry#sequences (accessed on 1 December 2022).
Granin proteins are essential for protein sorting and hormone packaging in the regulated secretory pathway, where proteins cross the Golgi apparatus to the trans-Golgi network. Under acidic, high-calcium conditions, SgII may be processed by prohormone convertases in the secretory vesicle. Secretory vesicles that carry SN fuse with the cell membrane, and SN is released through calcium-dependent exocytosis. Once released from the cell, it binds to the SN receptor in an autocrine, paracrine, or neuroendocrine manner
[5][9].
2. Biological Effects of Secretoneurin
In general, granins are predominantly known for their calcium-binding abilities. Granins have been extensively studied as neuroprotective factors and as co-factors of local immune and inflammatory reactions
[6][10]. Before SN was identified as a potential biomarker of heart failure, acute coronary syndrome, and arrhythmogenesis, the prognostic values of chromogranin A and B were studied in similar conditions with variable results. In particular, chromogranin A is an independent predictor of long-term mortality and heart failure-related hospitalization in patients with acute coronary syndromes
[7][11]. However, in patients with heart failure, chromogranin A did not show any additional value over contemporary markers
[8][12]. In contrast, chromogranin B is up-regulated in chronic heart failure, and its plasma levels are correlated with disease severity
[9][13].
SN has been detected in non-negligible amounts (range: 20–1500 pmol/L) in human serum, cerebrospinal fluid, and urine
[10][14]. SN has also been detected in human dental pulp and afferent nerve fibres that are frequently associated with blood vessels
[6][11][10,15]. SN immunoreactivity was also detected in the unmyelinated C-fibers which transmit predominantly nociceptive impulses
[11][15]. All the biological effects of SN are not completely understood.
ThWe
researchers ccan only speculate that the effects of SN may be exerted by SN receptors on the cell membrane. There is evidence that the SN receptor is a G-coupled membrane receptor
[12][16]. In addition, SN may also act directly at different subcellular levels. The biological effects of SN are summarized in
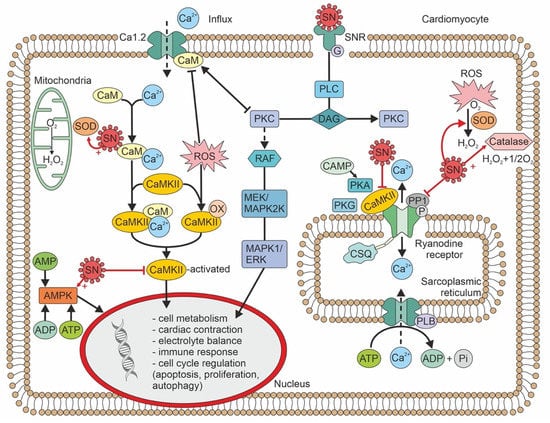
Figure 1.
Figure 1. Intracellular signaling pathways of secretoneurin, and biological effects. ADP—adenosine diphosphate; AMP—adenosine monophosphate; AMPK—adenosine monophosphate kinase; ATP—adenosine triphosphate; cAMP—cyclic adenosine monophosphate; CaM—calmodulin; CaMKII—calmodulin-dependent kinase II; CSQ—calsequestrin; DAG—diacylglycerol; ERK—extracellular-regulated kinase; MAPK—mitogen-associated protein kinase; MEK—MAPK/ERK kinase; OX—oxidized; SOD—superoxide dismutase; PLB—phospholamban; P—phosphorus; PKA—protein kinase A; PKC—protein kinase C; PKG—protein kinase G; PLC -phospholipase C; RAF—serine-threonine protein kinase; ROS—reactive oxygen species; SN—secretoneurin; SNR—secretoneurin receptor.
In concentrations 10x the plasma concentration, SN inhibits CaMKII by 20–30%. SN exerts its effect by binding to calmodulin
[13][3]. A direct effect, without binding to calmodulin, is thus again highly speculative. SN reduces ryanodine receptor (RyR)-mediated Ca
2+ leakage by inhibiting RyR phosphorylation. This activity leads to a reduced risk of ventricular arrhythmia
[14][4]. Furthermore, sarcoplasmic Ca
2+ leakage leads to calmodulin calcification, which binds and activates CaMKII; thus, SN may also indirectly inhibit CaMKII at plasma concentrations that correspond to the levels observed in heart disease
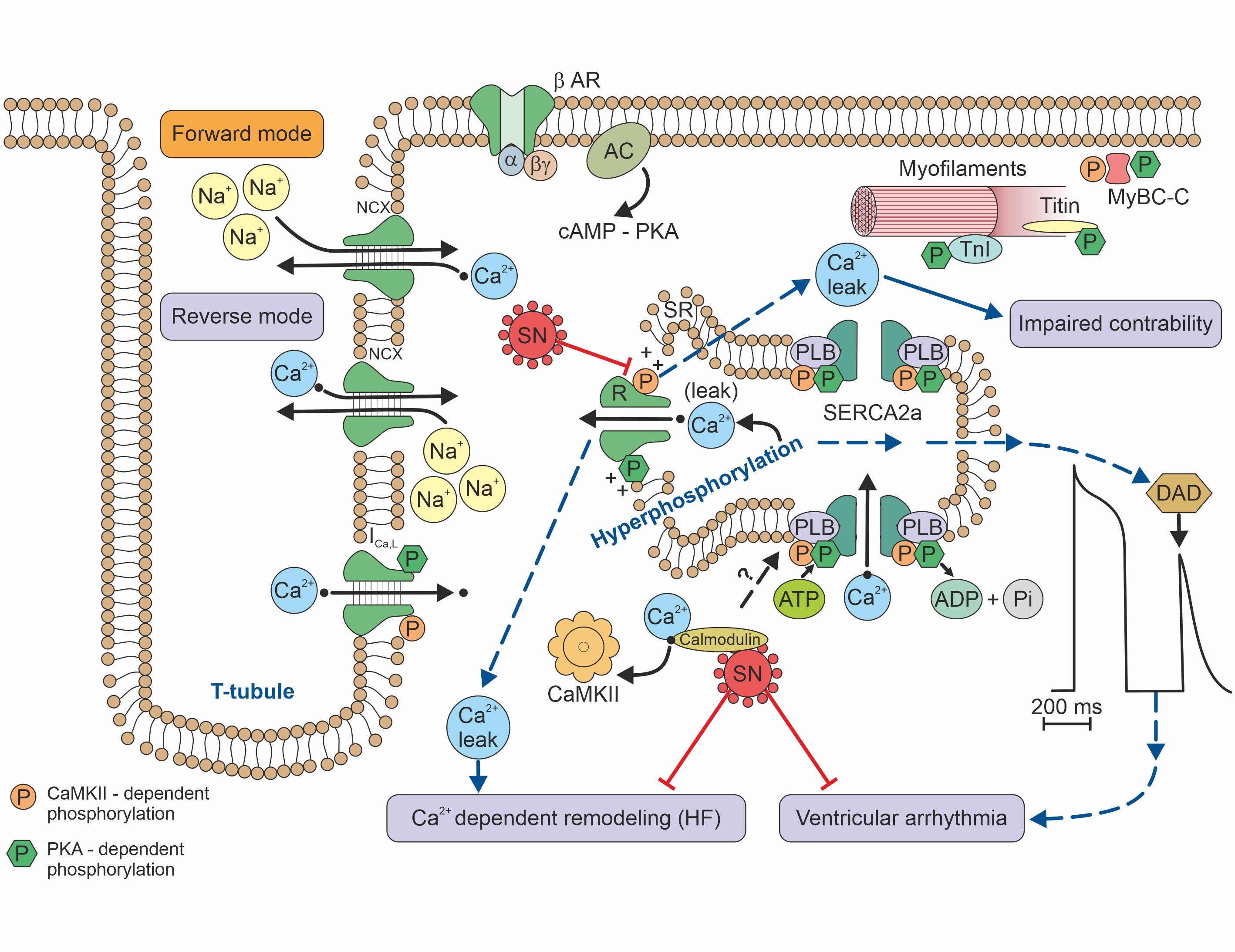
[14][4].For association of RyR function and risk of arrhythmia see Figure 2.
Figure 2. Calcium handling/signalling in a ventricular cardiomyocyte illustrates the potential involvement of SN in arrhythmogenic risk, heart failure, and calcium-dependent remodelling
Interestingly, SN may also exert an anti-apoptotic effect on endothelial cells. This SN effect could be initiated by the activation of phosphoinositide-3 kinase/protein kinase B and the mitogen-associated protein kinase (MAPK) pathway
[15][17]. In human umbilical vein endothelial cells, SN promoted proliferation and chemotaxis and reduced apoptosis
[16][18]. In a myocardial infarction model, SN also acted as an endogenous stimulator of vascular endothelial growth factor and induced angiogenesis
[17][19].
Of note, in animal models SN also enhances bone regeneration by increasing blood vessel and bone marrow formation
[18][20].
In contrast to its beneficial roles, SN may also trigger the selective migration of monocytes and fibroblasts. This activity could potentially enhance inflammatory responses that lead to the acceleration of atherosclerosis
[19][21].
3. Cellular Pathophysiology of Secretoneurin
Oxidative stress is a significant component of the cellular and subcellular processes that lead to inflammation and compensatory mechanisms, such as cell hypertrophy, fibrosis, and/or apoptosis. Reactive oxygen species (ROS) regulate mitogenic signal transduction in cardiomyocytes (evidenced in an animal experiment). Consequently, ROS may modulate hypertrophic signalling through MAPK pathways
[20][22]. At the cellular and molecular levels, hypertrophic adaptions are quite complex in nature; they include paracrine, autocrine, and circulating biologically active effectors that react to many stimuli. Among these stimuli, the most important is wall stress
[21][23].
Myocardial stretch triggers a chain of events: (1) the release of angiotensin II; (2) the release/formation of endothelin; (3) the activation of NADPH oxidase and transactivation of endothelium growth factor; (4) the production of mitochondrial ROS; (5) the activation of redox-sensitive kinases; (6) an increase in the intracellular sodium concentration; and (7) a transient increase in the Ca
2+ amplitude through the Na+/Ca
2+ exchanger. SN was found to attenuate isoproterenol-induced myocardial cell hypertrophy by suppressing ROS production and inducing the activities of superoxide dismutase and catalase
[22][24]. These effects are known to be mediated by adenosine-monophosphate or the extracellular-regulated kinase (ERK)/MAPK pathway. It is less probable that SN directly affects myocardial cell hypertrophy
[18][20]. Nevertheless, SN gene therapy (in an animal model of isoproterenol-induced hypertrophy) led to significant reductions in natriuretic peptide levels, the heart-weight to body-weight ratio, and surprisingly, interstitial fibrosis
[23][25].
Other results have indicated that SN, both acutely and after prolonged exposure, induced the augmented expression of endothelial nitric oxide synthase and calmodulin
[23][25]. In this respect, SN may play a protective role in ischaemic heart disease by enhancing endothelium-dependent relaxation. Endothelium-dependent relaxation may also involve the activation of cyclooxygenase, which would lead to the production of prostacyclin
[23][25].
SN may be implicated in the inflammatory response, because it modulates leukocytes, endothelial cells, and mesenchymal cells. SN promotes transendothelial leukocyte migration and adhesion; this activity is comparable to that promoted by tumour necrosis factor alpha
[24][26]. In response to injury, SN stimulates an increase in the migration and locomotion of human monocytes
[25][27]. In endothelium, SN activates the G-protein-coupled and protein kinase C-mediated signalling pathways. As a result, the cytoplasmatic calcium concentration rises
[24][26]. SN also inhibits the tight junctions between endothelial cells, which increases their permeability
[26][28]. Plasma SN concentrations that led to endothelium-derived relaxation in vitro were higher than those detected in humans with hypoxic conditions
[27][28][29,30]. However, plasma SN concentrations may not precisely reflect the tissue (endothelium) concentration. The most common pathological conditions that lead to increases in SN expression are the growth of active, predominantly neuroendocrine tumours and hypoxia
[9][29][13,31].