1. Introduction
Hemoglobin, a highly conserved protein, due to its ability to reversibly bind oxygen, is involved in the processes that underlie the aerobic life on planet Earth. The primary role of this protein is reflected in the maintenance of cellular homeostasis, by supporting its energy requirements. However, due to nearly 200 years of research into hemoglobin, it is now known that this protein also plays important roles in cellular signaling and modulation of the inflammatory response. Hemoglobin performs these functions directly, by binding gas molecules (NO, CO and CO
2), or indirectly, by acting as their source. In vertebrates, hemoglobin performs most of its functions while inside the erythrocytes
[1][2][3][1,2,3]. However, once hemoglobin reaches the extracellular environment, in conditions such as trauma, inflammation or infection, hemoglobin exerts other potentially harmful effects on cells and tissues and may be involved in the etiology and pathophysiology of various diseases
[4][5][6][4,5,6]. This dichotomy in function makes hemoglobin the so-called “two-edged sword”, a friend in physiological conditions, and an enemy or potential danger in stress conditions
[1][6][7][8][1,6,7,8]. Furthermore, through applied biomedical research which implies application of exogenous hemoglobin originating from different sources, novel insights into the modulatory capacities of this macromolecule are obtained.
2. Extracellular Hemoglobin in Mammals
2.1. Endogeous Extracellular Hemoglobin Removal and Degradation
Under physiological conditions, about 80–90% of erythrocytes are destroyed without releasing hemoglobin into plasma, in a process termed extravascular hemolysis. The remaining erythrocytes are removed under physiological conditions by a process designated as intravascular hemolysis—hemolysis within blood vessels
[8][9][8,10]. In intravascular hemolysis, hemoglobin is released directly into the circulation, where this molecule and its degradation products can cause cell and tissue damage
[10][11][11,12] if they exceed the capacity of the mechanisms involved in their removal. Free hemoglobin in plasma is rapidly oxidized to methemoglobin, which readily and non-enzymatically dissociates into heme and αβ dimers. At low plasma hemoglobin release, haptoglobin (Hp) irreversibly binds all αβ-globin dimers present
[12][13][13,14]. Hp binding sites are located on α-globin chains
[14][15]. Hp is synthesized in parenchymal cells of the liver, and the half-life of this glycoprotein in the circulation is 3.5–5 days. However, if it binds αβ-globin dimers, the half-life of such a complex becomes only 9 to 30 min. These complexes are rapidly removed from the circulation by phagocytosis by monocytes and tissue macrophages after binding to their CD163 receptor
[15][16]. Since there is no Hp recycling, once the reserves from circulation are depleted it takes 5–7 days for the pool to be recovered because a reduced Hp concentration does not lead to an increase in the synthesis of this molecule.
In addition to Hp, both hemopexin and albumin act as mechanisms that limit the effects of extracellular hemoglobin
[3][13][3,14]. These two molecules can bind free heme, maintain it in soluble form and thus prevent it from exhibiting its oxidative and proinflammatory effects
[16][17]. Hemopexin is a plasma heme-binding glycoprotein with an affinity for heme higher than all known heme-binding proteins. Moreover, hemopexin mediates the intake of heme in hepatocytes in which this prosthetic group is removed. Hemopexin is synthesized in the liver and in healthy individuals has a half-life of an average of seven days, while in the complex with heme, its half-life is reduced to 7–8 h
[17][18]. The heme-hemopexin complex enters hepatocytes through receptor-mediated endocytosis via a lipoprotein receptor-related protein-1 receptor (LRP), also known as CD91. After the endocytosis, the heme-hemopexin complex dissociates, and the hemopexin is released and returned to the plasma as an intact protein. Transport of heme in the cytoplasm occurs by an internal heme-binding membrane protein, and iron is rapidly removed by the action of heme oxygenase
[18][19][19,20].
Heme can also bind to albumin in the circulation and form methemalbumin. When added to human serum, heme is initially primarily bound to albumin, probably due to its high concentration relative to hemopexin. Removing methemalbumin from circulation is a kinetically complex process
[16][17]. Previous research shows that plasma heme can also bind α1-microglobulin (A1M), a 26 kDa glycoprotein synthesized in the liver and secreted into the blood. This protein has reductase activity, prevents intracellular oxidation, and reduces the expression of heme-induced heme oxygenase-1 (HO-1) and reactive oxygen species (ROS) production due to the presence of hemoglobin in the extracellular environment
[20][21][22][21,22,23]. The newest data suggest the use of apohemoglobin or apohemoglobin-Hp as a novel therapeutic strategy in scavenging and clearing excess heme through the monocyte/macrophage CD163 surface receptor
[23][24]. Subramanian et al.
[24][25] identified a “shortcut” for detoxification of extracellular hemoglobin within plasma, which functions Hp-independently via the capture and quench mechanism and involves the CD163 monocyte/macrophage receptor. Following hemoglobin recruitment, membrane CD163 (mCD163) directly suppresses the pseudoperoxidase activity of hemoglobin in situ on the monocyte membrane. Hemoglobin induces the release of mCD163 into plasma, and the resulting soluble CD163 (sCD163) further captures and quenches residual redox-reactive hemoglobin. These authors showed that sCD163 and immunoglobulin G (IgG) interact with extracellular hemoglobin in plasma. The resulting sCD163-Hb-IgG complex then triggers an autocrine loop of endocytosis via Fcγ receptors on monocytes and consequent recycling of internalized sCD163 via endosomes to establish mCD163 homeostasis, while internalized hemoglobin is catabolized by HO-1. Additionally, this complex induces paracrine transactivation of vascular endothelial cells, stimulates HO-1 expression in them, and secretes cytokines that will trigger a systemic defense response directed toward extracellular hemoglobin
[24][25].
2.2. Endogenous Extracellular Hemoglobin Mode of Action
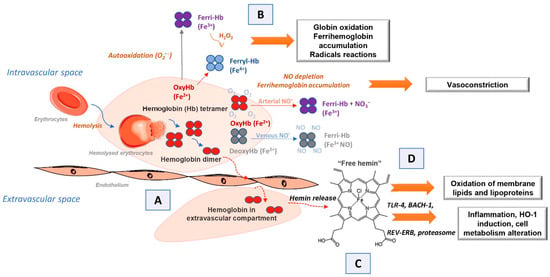
If extracellular hemoglobin exceeds the homeostatic mechanisms for its removal during intravascular hemolysis, it can affect surrounding cells and tissues. Various clinical aspects associated with circulating extracellular hemoglobin excess have been attributed to hemoglobin molecule-specific structural and biochemical characteristics through four proposed interacting mechanisms
[1][10][1,11]. The first of these is the extravascular translocation of hemoglobin (
Figure 1A)
[10][11]. After hemolysis, hemoglobin exists in the dynamic equilibrium of tetramers and αβ heterodimers, with a predominant dimer state at low plasma hemoglobin concentrations. Heterodimers are relatively small and capable of translocation and access to vulnerable anatomical sites (for example, glomeruli in the kidneys or vascular wall). Tissue exposure to hemoglobin most often accompanies cases of excessive hemoglobinuria after massive intravascular hemolysis, but hemoglobin can also translocate through endothelial barriers and thus enter the subendothelial and perivascular spaces and lymph
[25][26][26,27]. The hemoglobin dimer/oligomer is considered a damage/danger-associated molecular pattern (DAMP), while heme is independently recognized as an “alarmin”
[27][28][29][28,29,30].
Figure 1. Scheme illustrating the mechanisms of action of endogenous extracellular hemoglobin: (A) extravascular translocation of hemoglobin, (B) prooxidative reactivity of hemoglobin in plasma or within tissues after extravasation, (C) release of hemin from Hb-Fe3+ as the main product of oxidative reactions and (D) hemin induced changes in cell activation, gene expression, and metabolism.
Another mechanism by which extracellular hemoglobin exerts its effects is the prooxidative reactivity of hemoglobin in plasma or within tissues after extravasation (
Figure 1B)
[10][11]. The reactions of hemoglobin with NO and physiological oxidants (hydrogen peroxide and lipid peroxides) are best studied. NO consumption and consequent oxidation of hemoglobin occur through two reactions: (1) NO dioxygenation of oxyhemoglobin in which nitrates (NO
3−) and ferrihemoglobin (Hb-Fe
3+) are generated (Equations (1) and (2)) through nitrosylation of iron deoxyhemoglobin which occurs in direct binding of NO to iron of ferrohemoglobin (Hb-Fe
2+)
[10][30][31][11,31,32]. The binding of NO by cell-free hemoglobin leads to depletion of this critical vasodilator produced in the vascular endothelium; it is reported that this reaction is the fundamental cause of hypertension
[31][32], since even the recombinant hemoglobin, with diverse chemistries
[32][33], exerts a similar rate of reaction (Equation (1)). The reaction of extracellular hemoglobin and NO also results in the generation of Hb-Fe
3+ within the tissue parenchyma. The accumulation of Hb-Fe
3+ can stimulate the release and/or transfer of hemin to other proteins and lipids, thus manifesting the secondary toxicity of free hemin. Although hemoglobin and peroxide reactions have been the subject of numerous studies for over 40 years, the pathophysiology underlying these reactions is still not completely clarified
[2][10][18][33][2,11,19,34].
It has been shown that in vitro, the reaction of hemoglobin and peroxide leads to the formation of Hb-Fe
3+ and also to chemical species in which iron is in the ferryl form (Hb-Fe
4+) and its radical forms (Equations (2) and (3))
[4][31][34][35][4,32,35,36].
The outcome of such reactions is hemoglobin degradation, hemin loss, cross-linking, or precipitation of globin chains, which lead to tissue damage. It is still unknown how much Hb-Fe
4+ and radicals are formed in vivo during hemolysis and to what extent they contribute to the development of the disease. The only oxidized type of hemoglobin that can be consistently quantified in vivo is Hb-Fe
3+. In other words, it should be taken into account that the disparity observed in vitro and in vivo may be due to a shifted balance between oxidation and reduction reactions in vivo
[33][36][34,37]. The third proposed mechanism by which extracellular hemoglobin exerts its effects is the release of hemin from Hb-Fe
3+ as the main product of oxidative reactions (
Figure 1C)
[10][11]. The release of hemin allows the transfer of reactive porphyrin to the cell membrane or soluble plasma proteins and lipids and provides free hemin as a ligand for various signaling pathways. As hemin is a hydrophobic molecule, it is unlikely that more significant amounts of free hemin will be found in plasma; it binds rapidly to albumin or lipids and forms complexes. Depending on what it binds to, hemin can transform a molecule from a complex into a reactive product, such as oxidized low-density lipoprotein, further damaging the vasculature
[8][37][38][8,38,39]. The fourth mechanism refers to the effects achieved by hemin (
Figure 1D)
[10][11]. Hemin can selectively bind to several receptors and transcription factors and lead to changes in the state of cell activation, gene expression, and metabolism. The best-characterized interaction is hemin binding to transcriptional repressor 1, which shows homology to BTB and CNC homology 1, Bach-1, which regulates the transcription of HO-1 and other antioxidant enzymes necessary for an adaptive response to increased intracellular hemin levels
[39][40]. Hemin is also a ligand for the nuclear receptor of the hormone REV-ERB, which regulates circadian rhythm, glucose metabolism, and adipogenesis
[40][41].
3. Exogenous Administration of Extracellular Hemoglobin
The very first beginnings of hemoglobin application in biomedicine are related to its use as a blood substitute, i.e., more precisely as oxygen carriers (hemoglobin-based oxygen carriers, HBOCs)
[31][41][42][43][32,42,43,44] (
Figure 2). The first HBOC was hemoglobin isolated from lysed erythrocytes given to patients intravenously. However, it has been observed that soon after applying hemoglobin solution in this way, besides its short intravascular persistence, kidney damage, high blood pressure, and cardiovascular complications occur
[44][45][46][45,46,47] due to the effects of extracellular hemoglobin. These features and mechanisms of the act of extracellular hemoglobin are well known today and explained in detail in several comprehensive reviews and in the previous sections. Stroma-free hemoglobin rapidly dissociates into dimers and monomers, activates liver and spleen macrophages, and has the ability to bind the potent vasodilator, NO, resulting in vasoconstriction
[31][32]. Due to hemoglobin dimer precipitation in proximal tubules, nephrotoxic action occurs. Additionally, outside of erythrocytes, hemoglobin quickly oxidizes to methemoglobin, which cannot bind oxygen; instead, it releases heme, which participates in the creation of free radicals that exert harmful effects on surrounding cells
[42][43][47][48][43,44,48,49]. After such reports on outcomes of extracellular hemoglobin administration, researchers have focused on obtaining recombinant hemoglobin by mutagenesis with desired features and reduced side effects. With the use of recombinant technology, a low oxygen affinity human hemoglobin variant was produced in
Escherichia coli by Somatogen Inc. by introducing the substitution of amino acid Lys for Asn at the β108 position
[31][32]. To further stabilize this hemoglobin in tetrameric form, a site-specific mutation was introduced by inserting glycine into the region where two α subunits almost touch each other, fusing them (rHb1.0)
[49][50]. The second generation of recombinant hemoglobin (rHb2.0) included a change of heme pocket chemistry by introducing site-directed mutagenesis followed by polymerization
[50][51], such as providing HbA mutants with lower NO reactivity. A comprehensive summary of the impressive work done by biochemists in this field of recombinant hemoglobin is disclosed in the review of Varnado et al.
[51][52]. Progress in the development of recombinant hemoglobin enabled (i) adjustment of oxygen (O
2) affinity over a 100-fold range, (ii) reduction of nitric oxide (NO) scavenging activity over 30-fold without compromising dioxygen binding, (iii) decrease of antioxidant activity, (iv) reduction in hemin loss rate, (v) regulation of subunit dissociation rate, and (vi) decrease of irreversible subunit denaturation
[51][52]. Since the relatively high cost of large-quantity production of these proteins for emergency management of trauma patients is the main obstacle for their further progress, more cost-effective chemical modifications of the hemoglobin molecules have been launched
[42][52][53][54][55][56][57][58][59][60][61][62][43,53,54,55,56,57,58,59,60,61,62,63]. The procedure includes hemoglobin cross-linking/polymerization with succinyldisalicylic acid
[53][54], glycine
[54][55], glutaraldehyde
[55][56],
O-raffinose
[56][57][57,58] or modification of the surface area of hemoglobin molecules with polyethylene glycol (PEG)
[58][59][59,60] or human serum albumin with bound platinum nanoparticles
[60][61][61,62]. Another successful approach towards overcoming the side effects of a “naked hemoglobin molecule” administration involves the use of encapsulation as a method. Hemoglobin encapsulated in different biomaterials revealed longer circulatory lifetime, decreased hypertensive response and phagocytic uptake
[42][43]. From the pioneering steps in this approach by Chang et al.
[62][63], encapsulating material for hemoglobin varied from cellulose nitrate or PEG-polylactide to poly(ε-caprolactone)/poly(
l-lactic acid) and poly(
l-lysine), poly(lactic-coglycolicacid)/PEG copolymers and various lipid vesicles
[42][43]. Encapsulation of hemoglobin within emulsion with hydrophobic nanodrops (where hydrophobicity features of the nanoemulsion structures influence the binding of the hydrophobic niche of hemoglobin and thus partially act as a “stunt” -2, 3-DPG moderator) is patented demonstrating very promising results in the model of hemorrhagic rats
[63][64]. Recent studies on hemoglobin encapsulation in lipid vesicles demonstrated remarkable progress by means of larger-scale production without affecting the biophysical properties of hemoglobin molecules
[64][65] or introduction to clinical trials
[65][66].
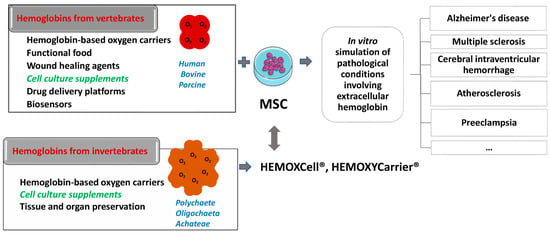
Figure 2. Schematic presentation of the potential use of vertebrates and invertebrates’ hemoglobin preparations in biomedicine and biotechnology (left side of the scheme), emphasizing already commercially available cell culture additives based on invertebrate hemoglobin (HEMOXCell®, HEMOXYCarriers®, HEMARINA SA, Morlaix, France). Up-to-date research on the impact of such hemoglobin preparation on mesenchymal stromal cell (MSC) cultures could significantly contribute to a better understanding of the onset and progression of pathological conditions involving extracellular hemoglobin as presented on the right side of the scheme.
Both allogeneic and xenogeneic hemoglobins were used as a source of extracellular hemoglobin. Among xenogeneic hemoglobins, the most studied are bovine and porcine hemoglobin, which show a high degree of homology with human hemoglobin (84.4% and 85%, respectively). Apart from the use as blood substitute, porcine hemoglobin was used as a wound healing agent in the form of spray if applied at regular intervals of 2–4 weeks. Namely, this product has been shown to reduce pain, wound size, and the necrotic tissue layer that impairs wound healing
[66][67][67,68]. Moreover, there are reports on bovine hemoglobin used as a drug platform itself, i.e., as a pH-sensitive nano-vehicles for potential cancer detection and therapy
[68][69] and as an effective glucose biosensor in vitro
[69][70]. One part of the research relies on examining rodent hemoglobin as well, aiming to simulate extracellular hemoglobin presence in vivo
[70][71]. Furthermore, hemoglobin from invertebrates has also become a significant subject of research. Thus, potential of extracellular hemoglobin as an additive for organ and tissue preservation
[71][72][72,73] and as a growth stimulator of mesenchymal stromal cells (MSC), which maintain ‘‘stemness” in vitro
[73][74], have been described. Therefore, one should be aware that under the same term “hemoglobin”, the studies on extracellular hemoglobin functions imply both vertebrate and evolutionary distant invertebrate hemoglobin.
Among invertebrate hemoglobins, the most investigated are extracellular hemoglobin of annelids (also named erythrocruorins and chlorocruorins after the nature of the porphyrin group)
[74][75][75,76], present in three classes of annelids:
Polychaete,
Oligochaeta, and
Achateae. Annelid hemoglobins are huge biopolymers with a high molecular weight of 3000–4000 kDa
[75][76][76,77]. The beginning of the application of EHb in biotechnology is related to the group led by Andre Tulmond and Franck Zal. This research group started an examination on hemoglobin from the marine polychaete
Arenicola marina (HbAm) in 1993 and described the structure of several extracellular hemoglobins in detail
[77][78]. It has been discovered that the hemoglobin of
Arenicola marina (HbAm) had all the sought traits of the universal oxygen carrier: it is naturally extracellular and polymerized, and its molecular mass is 50 times greater than the mass of human hemoglobin. It had properties of O
2-binding and release like human hemoglobin within erythrocytes. Each HbAm molecule can bind 156 oxygen molecules (human Hb can bind four) and has natural antioxidant properties. Additionally, HbAm has functional properties independent of secondary molecules such as 2,3-DPG in humans. The HbAm molecule displayed activity without any additional chemical modifications. It appears to be stable at temperatures ranging from 4 to >30 °C and does not show any vasoconstriction effects upon application
[75][76][76,77]. HbAm dissociates in human plasma from a hexagonal bilayer into globin dodecamers (although the dodecamer still appears to function as desired without extravasating)
[78][79][79,80]. It is important to note erythrocruorin of
Lumbricus terrestris (LtEc), which has been shown to be extremely stable, resistant to oxidation, and may interact with NO differently than mammalian hemoglobin molecules
[79][80].