Skin is the largest organ of the human body and is a great shield, as it protects it from external infections (environmental and chemical pollutants) as well as from UV irradiation. However, it is vulnerable since its degradation can occur both due to extrinsic and intrinsic factors, leading to early aging. Among all, extrinsic skin aging, called photoaging, is a remarkable result of oxidative stress caused by UV irradiation. In addition, reactive oxygen species (ROS) have also been found to contribute to skin aging, as they are produced in skin cells through UV irradiation, although at low concentrations they could be beneficial for some signaling pathways. Environmental and chemical pollutants also produce ROS, triggering a number of pathologies. Skin’s connective tissue includes a number of constituents, including collagen fibrils, elastic fibers, glycoproteins, and glycosaminoglycans. Among all, proteins like elastin, collagen, the glycosaminoglycan hyaluronic acid, and a polymeric pigment called melanin play pivotal roles in the regulation of skin’s elasticity as well as its protection against UV irradiation.
- database
- natural products
- medicinal plants
- antioxidants
- natural inhibitors
- anti-agings
1. Human Neutrophil Elastase (HNE)-A Serine Protease
2. Hyaluronidase (Hyal)—A Glycosyl Hydrolase
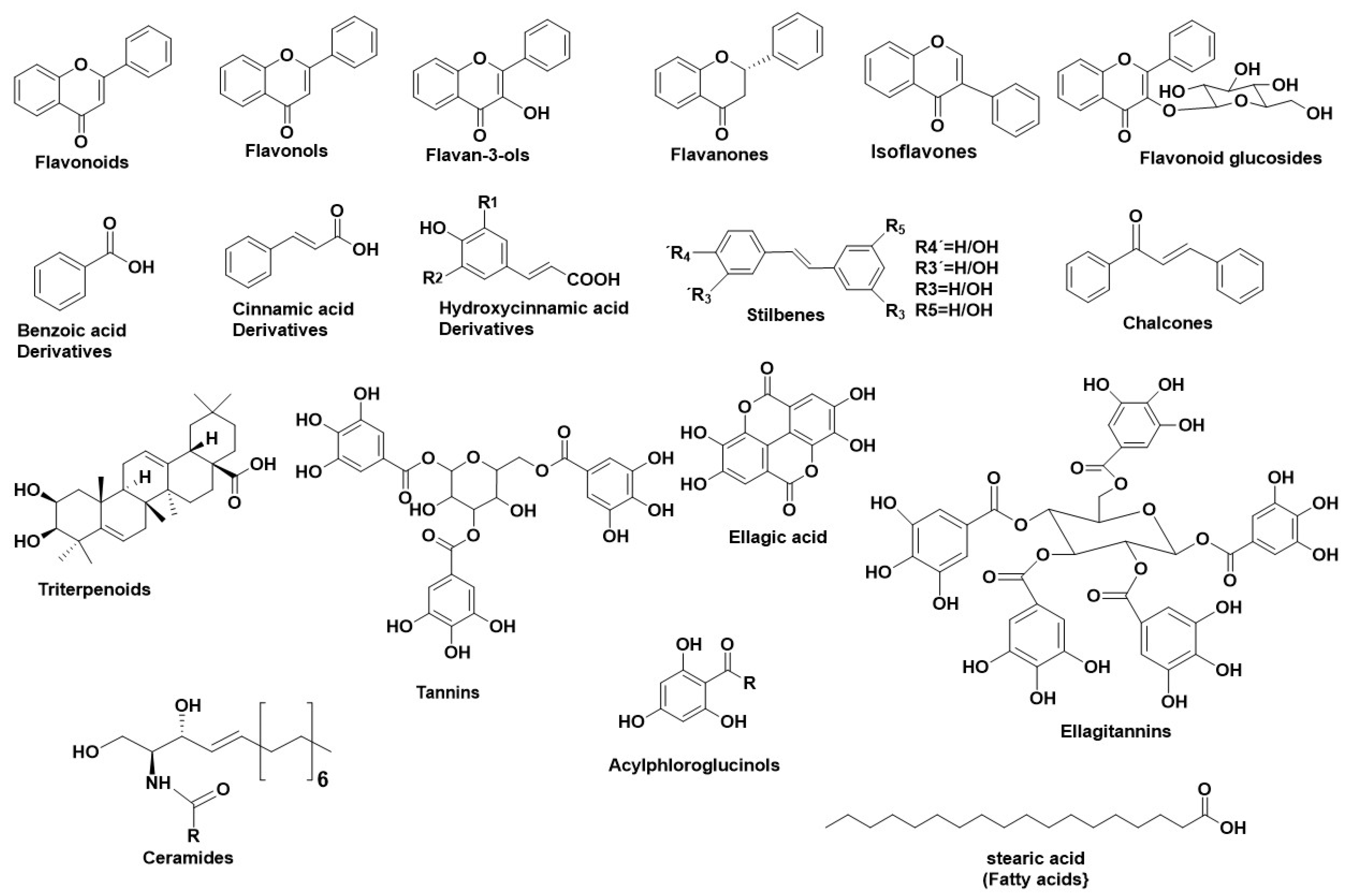
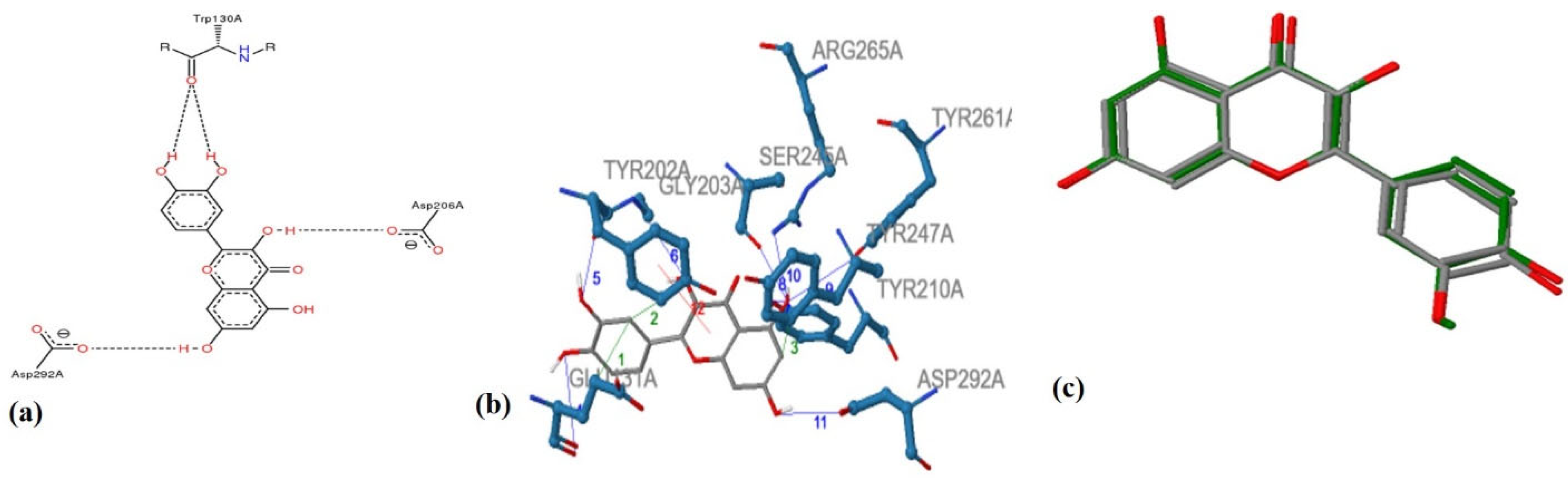
Hyalouronidases (Hyals) [E.C. 3.2.1.35] consist of a subclass of glycosyl hydrolases mainly responsible for degrading the glycosaminoglycan Hyaluronic Acid (HA) [22] by cleaving its β-1→4 glycosidic bonds into oligomers. HA can be found in abundance in synovial fluid, in cartilage (mainly in the soft part of connective tissue), and in the vitreous part of the eye [23]. Furthermore, HA can be found in the extracellular matrix as a connecting fluid of protein filaments, collagen fibers, and connective tissue cells [24]. Studies made on Hyals by Karl Meyer (1971) [25] differentiated them into three groups according to their reaction mechanism: (1) bacterial hyaluronidases (E.C. 4.2.99.1) (endo-β-acetyl-hexosaminidases), (2) hyaluronidases (E.C. 3.2.1.36) (endo-β-glucuronidases), and (3) mammalian hyaluronidases (E.C. 3.2.1.35) (hydrolases, producing tetrasaccharides and hexasaccharides). Hyals contain five homologous molecules expressed in the human gene: Hyal-1, Hyal-2, Hyal-3, Hyal-4, and Hyal-5 (SPAM-1 or PH-20), as well as a pseudogene (pHYAL1) [22]. Hyal-1, Hyal-2, and Hyal-3 are found in chromosome 3p21.3 [26], whereas Hyal-4, PHYAL1, and SPAM1 are found in chromosome 7q31.3 [27]. Hyal-1, Hyal-2, and PH-20 regulate HA’s stability [28], whereas Hyal-3, Hyal-4, and PHYAL1 do not affect HA’s metabolism [29]. Among all Hyals, human Hyal-1 is abundant in the somatic tissue and in human serum (~60 ng/mL) as well as in liver, kidney, heart, plasma etc. This lysosomal enzyme contains three N-glycosylated sites at places N-99, N-216, and N-350 [30] and degrades HA into GlcA-GlcNAc-GlcA-GlcNAc tetrasacharides [23]. According to numerous studies, Hyal-1 is expressed in various cancer cells, like in the prostate, bladder, and brain [31][32], and is also connected to the human cancer cell lines MCF-7 and MDA-MB-231 [33]. Hyal-1 is of great interest, as a well-developed study is determining compounds which act as natural Hyal-1 inhibitors against diseases like arthritis and gingivitis [34][35]. Hyal-2 is widely found in connective tissue at high concentration and decomposes HA into oligomers of 50 saccharide units (~20 kDa) [36]. The activity of Hyal-2 ranges between various pH values, whereas Hyal- 1 and Hyal-3 are active in acidic conditions. Hyal-4 behaves also as a chondroitinase, using Chondroitin (Ch) and Chondroitin Sulfate (ChS) as substrates. Additionally, SPAM1 is located in the acrosome of spermatids and enables the adhesion of sperm to the cumulus mass protecting the ovum [36][37]. Hyals’ catalysis depends on an acidic active site Glu-131, which donates H [22]. Hyal’s active site also contains Tyr75 Trp141, Tyr201, Tyr208, Tyr210, Tyr247, Tyr261, Tyr286, and Trp321, which are responsible for its catalysis. The active site, positively charged hydrophobic amino acids enable the complete interaction not only with HA, but also with other negatively charged substrates (e.g., Chondroitin and Chondroitin Sulfate) [36]. Furthermore, Hyal’s structure contains two arginines (Arg134 and Arg265), which interact with -COOH groups of HA, as well as a Ser245, which is stable and enables bond formation with the −OH group of Tyr202 [23][30][36]. Hyal-4 has Cys263 instead of Tyr247, which enables the interaction with Chondroitin and Chondroitin Sulfate [36]. Numerous studies regarding the anti-Hyal potency of plants, their extracts, or their isolated metabolites have showed that the most potent natural inhibitors are terpenes, flavonoids and alkanoids [22]. Figure 2 illustrates the main structural scaffolds which have been marked as efficient Hyal inhibitors. The natural products that have been studied for their inhibitory properties towards Hyal can be explored in the new developed Server and Database called ANTIAGE-DB, through the link https://bio-hpc.ucam.edu/anti-age-db.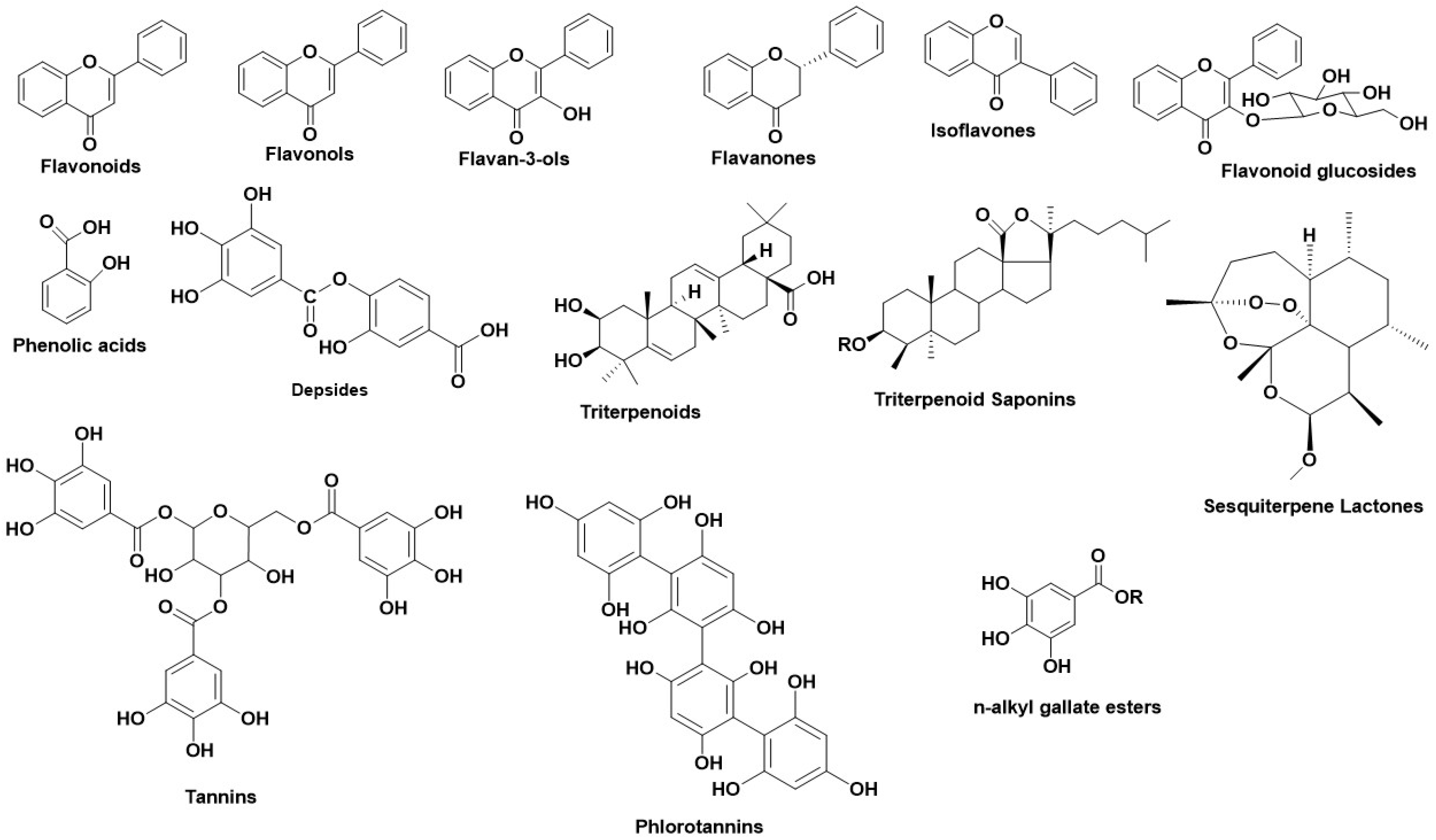
3. Tyrosinase—A Polyphenol Oxidase
Tyrosinases (catechol oxidases, catecholases, diphenol oxidases, o-diphenolases, phenolases and polyphenol oxidases) [ΕC 1.14.18.1] are copper-containing glycoproteins and can be found in various active forms in living organisms (mammals, plants, fungi and animal tissues) [38]. The most vital activity of Tyrs is their implication in melanin and other pigments synthesis, as well as the brown color in fruits and vegetables, caused by the oxidation of the present Tyr in their flesh with the atmospheric molecular oxygen. In humans, Tyrs are located in the melanosomes, which are synthesized in the skin melanocytes. Tyrs have a double enzymatic activity, in the presence of O2: (1) they act as monophenolases (cresolases) and hydroxylate monophenols in their ortho-position and (2) they act as diphenolases (catecholases) and oxidize o-diphenols into o-quinones. Tyr oxidizes L-tyrosine into dopaquinone, a product which leads to the formation of eumelanin (brown or black melanin) and pheomelanin (red to yellow melanin) [38]. Specifically, Tyr structure consists of three domain parts. The central, and most characteristic domain, is the active site binuclear copper moiety, which enables the bond formation between Tyr and molecular O2, as well as with Tyr’s substrate [39]. The other domains contain six histidines, which form bonds with the cuprous pair inside the active site. The Tyr structure is stabilized through disoulfide bridges, formed by cysteines around the active site. Tyrs from different plants differ in their number of cysteines; human and mouse Tyrs have 17 cysteines, whereas plant Tyrs have 11 cysteines. Furthermore, plant Tyrs contain one cysteine in their Carboxylic terminal moiety, whereas Tyrs from N. crassa, A. bisporus, and prokaryotic Tyrs have 0 or 1 cysteines [40]. Numerous studies revealed various natural metabolites as Tyr inhibitors, along with their interacting properties with Tyr’s active site through combined studies on docking simulations and kinetics [41]. The structure of Tyr is similar to hemoqyanin, and a lot of correlated results regarding the binding properties of Tyr can be excluded [41][42][43]. Wang et al., developed a fluorescent probe with which they determined the mechanism of Tyr reaction [44] finding that the inhibitors react with Tyr irreversibly [38]. Tyr inhibition is achieved either by covering the active site or by producing undesirable dopaquinone derivatives or by completing multisite reactions. For this reason, several studies have been performed regarding the inhibitory activity of natural compounds, their type of inhibition, including also their interaction profile. Figure 3 illustrates the main structural scaffolds which have been marked as efficient Tyr inhibitors. The natural products that have been studied for their inhibitory properties towards Tyr can be explored in the new developed Server and Database called ANTIAGE-DB, through the link https://bio-hpc.ucam.edu/anti-age-db.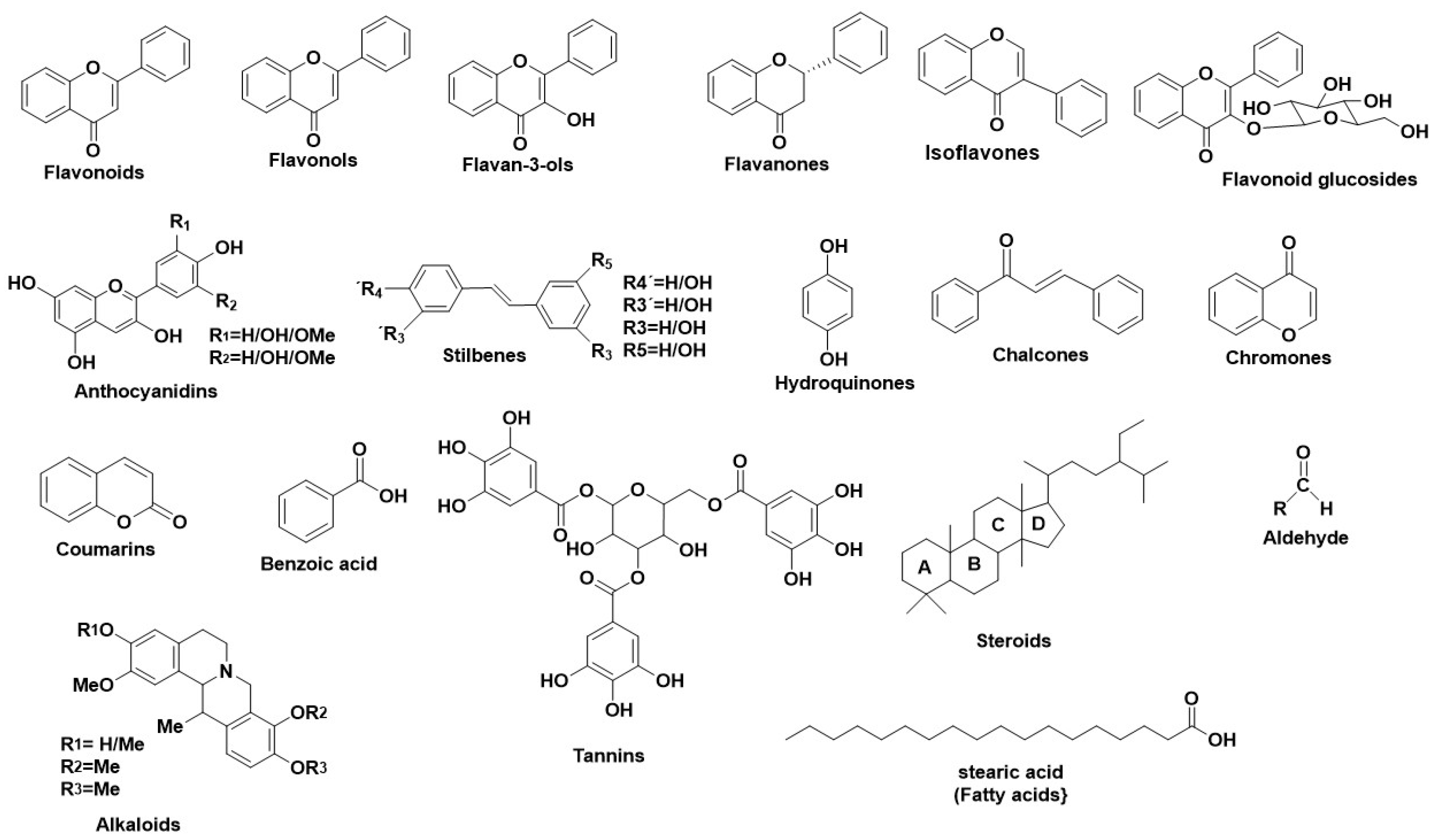
References
- Bieth, J.G. The elastases. J. Soc. Biol. 2001, 195, 173–179. PMID: 11723830
- Takahashi, H.; Nukiwa, T.; Yoshimura, K.; Quick, C.D.; States, D.J.; Holmes, M.D.; Whang-Peng, J.; Knutsen, T.; Crystal, R.G. Structure of the human neutrophil elastase gene. J. Biol. Chem. 1988, 263, 14739–14747. PMID: 2902087
- Melzig, M.F.; Löser, B.; Ciesielski, S. Inhibition of neutrophil elastase activity by phenolic compounds from plants. Pharmazie 2001, 56, 967–970. PMID: 11802662
- Siedle, B.; Cisielski, S.; Murillo, R.; Lo, B.; Castro, V.; Klaas, C.A.; Hucke, O.; Labahn, A.; Melzig, M.F.; Merfort, I.; et al. Sesquiterpene Lactones as Inhibitors of Human Neutrophil Elastase. Bioorg. Med. Chem. 2002, 10, 2855–2861. DOI: 10.1016/s0968-0896(02)00149-9
- Kim, Y.-J.; Uyama, H.; Kobayashi, S. Inhibition effects of (+)-catechin-aldehyde polycondensates on proteinases causing proteolytic degradation of extracellular matrix. Biochem. Biophys. Res. Commun. 2004, 320, 256–261. DOI: 10.1016/j.bbrc.2004.05.163
- Thring, T.S.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. DOI: 10.1186/1472-6882-9-27
- Belaaouaj, A.; Kim, K.S.; Shapiro, S.D. Degradation of outer membrane protein A in Escherichia coli killing by neutrophil elastase. Science 2000, 289, 1185–1188. DOI: 10.1126/science.289.5482.1185
- Ying, Q.L.; Rinehart, A.R.; Simon, S.R.; Cheronis, J.C. Inhibition of human leucocyte elastase by ursolic acid. Evidence for a binding site for pentacyclic triterpenes. Biochem. J. 1991, 277, 521–526. DOI: 10.1042/bj2770521
- Bode, W.; Edgar Meyer, J.C.P., Jr. Perspectives in Biochemistry. Human Leukocyte and PorcinePancreatic Elastase: X-ray Crystal Structures, Mechanism, Substrate Specificity. Biochemistry 1989, 28, 1951–1963. DOI: 10.1021/bi00431a001
- Feng, L.; Liu, X.; Zhu, W.; Guo, F.; Wu, Y.; Wang, R.; Chen, K.; Huang, C.; Li, Y. Inhibition of human neutrophil elastase by pentacyclic triterpenes. PLoS ONE 2013, 8, e82794. DOI: 10.1371/journal.pone.0082794
- Tamada, T.; Kinoshita, T.; Kurihara, K.; Adachi, M.; Ohhara, T.; Imai, K.; Kuroki, R.; Tada, T. Combined high-resolution neutron and X-ray analysis of inhibited elastase confirms the active-site oxyanion hole but rules against a low-barrier hydrogen bond. J. Am. Chem. Soc. 2009, 131, 11033–11040. DOI: 10.1021/ja9028846
- Hess, G.P.; McConn, J.; Ku, E.; McConkey, G. Studies of the activity of chymotrypsin. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1970, 257, 89–104. DOI: 10.1098/rstb.1970.0011
- Siedle, B.; Hrenn, A.; Merfort, I. Natural compounds as inhibitors of human neutrophil elastase. Planta Med. 2007, 73, 401–420. DOI: 10.1055/s-2007-967183
- Löser, B.; Kruse, S.O.; Melzig, M.F.; Nahrstedt, A. Inhibition of neutrophil elastase activity by cinnamic acid derivatives from Cimicifuga racemosa. Planta Med. 2000, 66, 751–753. DOI: 10.1055/s-2000-9563
- Melzig, M.F.; Löser, B.; Lobitz, G.O.; Tamayo-Castillo, G.; Merfort, I. Inhibition of granulocyte elastase activity by caffeic acid derivatives. Pharmazie 1999, 54, 712. PMID: 10522278
- Hamburger, M.; Riese, U.; Graf, H.; Melzig, M.F.; Ciesielski, S.; Baumann, D.; Dittmann, K.; Wegner, C. Constituents in Evening Primrose Oil with Radical Scavenging, Cyclooxygenase, and Neutrophil Elastase Inhibitory Activities. J. Agric. Food Chem. 2002, 50, 5533–5538. DOI: 10.1021/jf025581l
- Xing, X.; Yang, X.; Cao, Y. Study of Ellagic Acid as a Natural Elastase Inhibitor by Spectroscopic Methods. J. Appl. Spectrosc. 2016, 83, 149–155. DOI:https://doi.org/10.1007/s10812-016-0259-4
- Rennert, B.; Melzig, M.F. Free fatty acids inhibit the activity of Clostridium histolyticum collagenase and human neutrophil elastase. Planta Med. 2002, 68, 767–769. DOI: 10.1055/s-2002-34411
- Bizot-Foulon Godeau, G.; Guessous, F.; Lati, E.; Rousset, G.; Roch-Arveillier, M.; Hornebeck, W.V. Inhibition of human neutrophil elastase by wheat ceramides. Int. J. Cosmet. Sci. 1995, 17, 255–264. DOI: 10.1111/j.1467-2494.1995.tb00130.x
- Sim, G.S.; Lee, B.C.; Cho, H.S.; Lee, J.W.; Kim, J.H.; Lee, D.H.; Kim, J.-H.; Pyo, H.B.; Moon, D.C.; Oh, K.W.; et al. Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharmacol. Res. 2007, 30, 290–298. DOI: 10.1007/BF02977608
- Wittenauer, J.; Mäckle, S.; Sußmann, D.; Schweiggert-Weisz, U.; Carle, R. Inhibitory effects of polyphenols from grape pomace extract on collagenase and elastase activity. Fitoterapia 2015, 101, 179–187. DOI: 10.1016/j.fitote.2015.01.005
- Załuski, D.; Cieśla, Ł.; Janeczko, Z. Chapter 7—The Structure—Activity Relationships of Plant Secondary Metabolites with Antimicrobial, Free Radical Scavenging and Inhibitory Activity toward Selected Enzymes. Stud. Nat. Prod. Chem. 2015, 45, 217–249. DOI:10.1016/B978-0-444-63473-3.00007-1
- Marković-Housley, Z.; Miglierini, G.; Soldatova, L.; Rizkallah, P.J.; Müller, U.; Schirmer, T. Crystal structure of hyaluronidase, a major allergen of bee venom. Structure 2000, 8, 1025–1035. DOI: 10.1016/s0969-2126(00)00511-6
- Laurent, T.C.; Fraser, J.R. Hyaluronan. FASEB J. 1992, 6, 2397–2404. PMID: 1563592
- Meyer, K. Hyaluronidases. In The Enzymes; Academic Press: New York, NY, USA, 1971; pp. 307–320. DOI: 10.1021/cr050247k
- Stern, R. Devising a pathway for hyaluronan catabolism: Are we there yet? Glycobiology 2003, 13, 105–115. DOI: 10.1093/glycob/cwg112
- Orlando, Z.; Lengers, I.; Melzig, M.F.; Buschauer, A.; Hensel, A.; Jose, J. Autodisplay of human hyaluronidase Hyal-1 on Escherichia coli and identification of plant-derived enzyme inhibitors. Molecules 2015, 20, 15449–15468. DOI: 10.3390/molecules200915449
- Jedrzejas, M.J.; Stern, R. Structures of vertebrate hyaluronidases and their unique enzymatic mechanism of hydrolysis. Proteins 2005, 61, 227–238. DOI: 10.1002/prot.20592
- Csoka, A.B.; Frost, G.I.; Stern, R. The six hyaluronidase-like genes in the human and mouse genomes. Matrix Biol. 2001, 20, 499–508. DOI: 10.1016/s0945-053x(01)00172-x
- Chao, K.L.; Muthukumar, L.; Herzberg, O. Structure of human hyaluronidase-1, a hyaluronan hydrolyzing enzyme involved in tumor growth and angiogenesis. Biochemistry 2007, 46, 6911–6920. DOI: 10.1021/bi700382g
- Lokeshwar, V.B.; Rubinowicz, D.; Schroeder, G.L.; Forgacs, E.; Minna, J.D.; Block, N.L.; Nadji, M.; Lokeshwar, B.L. Stromal and epithelial expression of tumor markers hyaluronic acid and HYAL1 hyaluronidase in prostate cancer. J. Biol. Chem. 2001, 276, 11922–11932. DOI: 10.1074/jbc.M008432200
- Lokeshwar, V.B.; Obek, C.; Pham, H.T.; Wei, D.; Young, M.J.; Duncan, R.C.; Soloway, M.S.; Block, N.L. Urinary hyaluronic acid and hyaluronidase: Markers for bladder cancer detection and evaluation of grade. J. Urol. 2000, 163, 348–356. DOI: 10.1016/s0022-5347(05)68050-0
- Tan, J.X.; Wang, X.Y.; Li, H.Y.; Su, X.L.; Wang, L.; Ran, L.; Zheng, K.; Ren, G.S. HYAL1 overexpression is correlated with the malignant behavior of human breast cancer. Int. J. Cancer 2011, 128, 1303–1315. DOI: 10.1002/ijc.25460
- Mio, K.; Stern, R. Inhibitors of the hyaluronidases. Matrix Biol. 2002, 21, 31–37. DOI: 10.1016/s0945-053x(01)00185-8
- Jentsch, H.; Pomowski, R.; Kundt, G.; Göcke, R. Treatment of gingivitis with hyaluronan. J. Clin. Periodontol. 2003, 30, 159–164. DOI: 10.1034/j.1600-051x.2003.300203.x
- Stern, R.; Jedrzejas, M.J. Hyaluronidases: Their genomics, structures, and mechanisms of action. Chem. Rev. 2006, 106, 818–839. DOI: 10.1021/cr050247k
- Hunnicutt, G.R.; Primakoff, P.; Myles, D.G. Sperm surface protein PH-20 is bifunctional: One activity is a hyaluronidase and a second, distinct activity is required in secondary sperm-zona binding. Biol. Reprod. 1996, 55, 80–86. DOI: 10.1095/biolreprod55.1.80
- Lee, S.Y.; Baek, N.; Nam, T.G. Natural, semisynthetic and synthetic tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2016, 31, 1–13. DOI: 10.3109/14756366.2015.1004058
- Jackman, M.P.; Hajnal, A.; Lerch, K. Albino mutants of Streptomyces glaucescens tyrosinase. Biochem. J. 1991, 274 Pt 3, 707–713. DOI: 10.1042/bj2740707
- van Gelder, C.W.; Flurkey, W.H.; Wichers, H.J. Sequence and structural features of plant and fungal tyrosinases. Phytochemistry 1997, 45, 1309–1323. DOI: 10.1016/s0031-9422(97)00186-6
- Himmelwright, R.S.; Eickman, N.C.; Lu Bien, C.D.; Lerch, K.; Solomon, E. Chemical and Spectroscopic Studies of the Binuclear Copper Active Site of Neurospora Tyrosinase: Comparison to Hemocyanins. J. Am. Chem. Soc. 1980, 102, 7339–7344. DOI: https://doi.org/10.1021/ja00544a031
- Magnus, K.A.; Hazes, B.; Ton-That, H.; Bonaventura, C.; Bonaventura, J.; Hol, W.G.J. Crystallographic Analysis of Oxygenated and Deoxygenated States of Arthropod Hemocyanin Shows Unusual Differences. Genet. Proteins Struct. Funct. 1994, 19, 302–309. DOI: 10.1002/prot.340190405
- Volbeda, A.; Hol, W.G. Crystal Structure of Hexameric Haemocyanin from Panulirus Interruptus Refined at 3.2 A Resolution. J. Mol. Biol. 1989, 209, 249–279. DOI: 10.1016/0022-2836(89)90276-3
- Wang, C.; Yan, S.; Huang, R.; Feng, S.; Fu, B.; Weng, X.; Zhou, X. A turn-on fluorescent probe for detection of tyrosinase activity. Analyst 2013, 138, 2825–2828. DOI: 10.1039/c3an00272a