1. Therapeutic Potential of SARS-CoV-2 S-Glycoprotein
As S-glycoprotein plays an unavoidable role in the SARS-CoV-2 entry into human cells, it represents the main target for vaccine design and biologics development against COVID-19 infection
[1][13]. Once the SARS-CoV-2 was imported into the host cells, the human immune system mainly recognized its surface epitopes located in S-glycoprotein resulted in innate and adaptive immune responses
[2][36]. These immune reactions are contributed mainly by Toll-like receptors (TLR3, 7, and 8) causing enhanced production of interferon. After the replication of the virus within the lung cells, cytokines (IL-6, IL-8, IL-10, and TNF-a) and chemokines were secreted by alveolar macrophages which functioned as inflammatory mediators. Simultaneously, neutrophils, natural killer cells, monocytes, CD4+, and CD8+ T cells initiated the phagocytosis of infected cells
[3][37]. Moreover, whole S-glycoprotein or its immunogenic fragments (amino acids from 100–280, 430–590, and 1060–1150) can trigger long-lasting dominant neutralizing immune cells against SARS-CoV-2
[4][11]. As a consequence, antibodies against recombinant whole or specific fragments of S-glycoprotein could block virus entry and also inhibit viral replication
[5][38]. The S1 subunit is a receptor-binding element that contains an
N-terminal signal peptide, an
N-terminal domain (so-called NTD), and the RBD domain. While the S2 subunit is composed of a fusion peptide domain (FP), two heptad-repeat domains (HR1 and HR2), a transmembrane domain, and a
C-terminal domain that functions in the fusion of viral and host membranes facilitating virus entry
[6][25]. Notably, S-glycoprotein is highly glycosylated with 22
N-glycosylation sites and 17
O-glycosylation sites. Thus, the glycosylation processes are crucial for the correct folding and biological activity of S-glycoprotein. Moreover, the glycosylation profile of S-glycoprotein is critical for host recognition, binding, penetration, and pathogenesis of the SARS-CoV-2 virus
[7][39]. The S-glycoprotein plays a critical role in the initiation of immune responses such as secretion of neutralizing-antibody, and T-cell-related reactions during SARS-CoV-2 infection
[8][26].
HereIn thi
ns context, this glycoprotein is considered as the pivotal antigenic molecule that leads to the production of immunity-related elements such as neutralizing mAbs against virus infection
[1][13]. Initially, it was reported that antibodies produced during COVID-19 infection are enabled to bind to the RBD of both SARS-CoV-1 and SARS-CoV-2, suggesting that this region would undergo less mutation
[1][13]. Later on, this theory was rejected as new SARS-CoV-2 variants carrying a different number of insertion or deletion mutations in their S-glycoproteins, even in the RBD region, have emerged
[9][19]. Thus far, the potential of the entire S-glycoprotein and its antigenic elements, such as the S1 subunit through the RBD domains, have been evaluated as candidates for SARS-CoV-2 vaccine construction. Although full-length S-glycoprotein-related vaccines can induce potent neutralizing antibodies and other immune responses, some reports have shown that the secreted antibodies may enhance viral infection after reinfection with a SARS-CoV-2 homologous or cause liver damage
[10][11][40,41].
As an alternative approach, immunogenic fragments of S-glycoprotein could be targeted for the production of subunit vaccines against SARS-CoV-2. For example, vaccines based on the S1-related subunit could significantly elicit the production of neutralizing antibodies providing strong protective immunity against viral infection. Of note, the S2 fragments had lower immunogenicity, resulting in lower antibody induction and fewer immune responses against viral infection
[10][40]. In addition to vaccine construction, S-glycoprotein has been regularly used for the construction of diagnostic kits for infected patients with COVID-19
[12][42]. Indeed, most molecular diagnosis kits (RT-PCR) against SARS-CoV-2 are designed using the S1 subunit, as it has less cross-reactivity than the full-length S-glycoprotein, less similarity to human-related coronaviruses, and more immunogenicity than the S2 subunit
[13][14][43,44].
2. ACE2 Is Also a Promising Approach for the Treatment of COVID-19 Infection
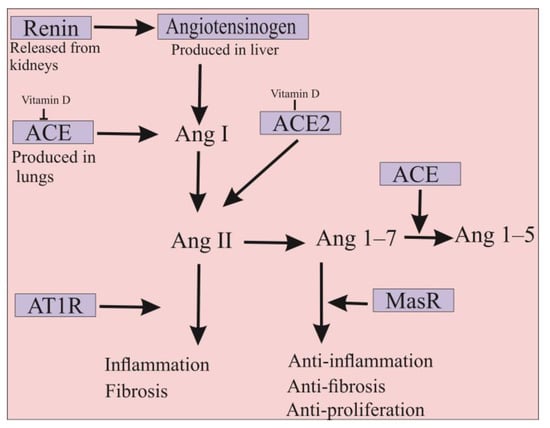
The ACE2 is a type 1 membrane glycoprotein that is an enzymatically active homolog of ACE. ACE is the major component of the renin-angiotensin-aldosterone system (RAAS), involved in blood pressure regulation, electrolyte, fluid homeostasis, natriuresis, and salt balance
[15][45]. In detail, angiotensinogen produced in the liver is cleaved by renin, resulting in the formation of angiotensin I (Ang I) decapeptide. Subsequently, ACE protein converts Ang I to the octapeptide Ang II, which imposes its physiological function mainly including blood pressure increase, vasoconstriction, aldosterone synthesis, and induction of inflammatory and pro-fibrotic pathways through the angiotensin II type I receptor (AT1R)
[16][17][18][10,46,47]. Despite the similarity of more than 61% in the amino acid sequences of their catalytic domains, several differences exist between ACE and ACE2. ACE is a dipeptidyl-carboxypeptidase enzyme with two zinc-binding motifs in its
N- and
C-terminal domains, whereas ACE2 is a mono-carboxypeptidase with one zinc-binding motif in its
N-terminus domain
[19][48]. Therefore, ACE2 could act as a negative regulator of RAAS pathways in which it hydrolyzes Ang II into the Ang 1–7 inducing anti-inflammatory, anti-fibrotic, and anti-proliferation cascades via its binding to the Mas receptor (MasR)
[16][20][10,49]. Indeed, ACE2 actively counteracts the physiological effects of ACE, whereby disruption of the ACE/ACE2 balance in humans can cause several intense disorders (
Figure 12)
[21][22][50,51]. It should be noted that ACE2 is expressed in its functional form in most vital tissues, especially in healthy human organs such as the lung, skin, oral and nasal mucosa, nasopharynx, gastrointestinal system, liver, kidney, and brain tissues
[23][52]. Importantly, the highest surface expression of ACE2 protein is reported in alveolar epithelial cells of the lung and enterocytes of the small intestine, where human cells are in contact with the external environment, facilitating the entry of pathogens such as SARS-CoV-2
[24][27].
Figure 12. Differences between ACE2 and ACE functions. ACE is the major component of the renin-angiotensin-aldosterone system (RAAS) and is involved in the inflammation and fibrosis pathways. ACE2 is a homolog of ACE that counteracts the physiological function of ACE in order that the protein has anti-inflammation and anti-fibrosis properties. In addition, ACE2 has anti-proliferation properties that can significantly inhibit SARS-CoV-2 infection. Moreover, vitamin D is involved in the suppression of ACE and also in the increase of ACE2 expression. Therefore, the administration of this molecule could be used to prevent SARS-CoV-2 infection. Ang; angiotensin, AT1R; angiotensin II type I receptor and MasR; Mas receptor.
The human ACE2 is composed of three regions, namely, extracellular, transmembrane, and intracellular segments, among which the extracellular part contains a peptidase domain that participates in binding to the SARS-CoV-2 RBD
[25][53]. The binding of S-glycoprotein to ACE2 is the initial step in the entry, replication, and subsequent pathogenesis of SARS-CoV-2. In vitro and in vivo studies revealed that ACE2 is the only functional receptor for SARS-CoV-2 entry into human cells (
Figure 1)
[24][27]. The overexpression of ACE2 originated from humans, Chinese horseshoe bats, civets, and pigs in the HeLa cells allowed SARS-CoV-2 infection, confirming that the virus uses ACE2 as an entry receptor
[26][54]. Further data showed that SARS-CoV-2 could not use other coronavirus receptors such as the aminopeptidase N and dipeptidyl peptidase 4
[24][26][27,54]. It was also previously shown that mice infected with SARS-CoV-1 had a significant reduction in ACE2 expression in their lungs
[27][28]. Given the high similarity between SARS-CoV-1 and SARS-CoV-2, it could be hypothesized that decreased ACE2 expression levels may play an important role in the pathogenesis of both viruses. Notably, overexpression of human ACE2 can significantly increase the severity of disease in SARS-CoV-infected mice
[28][55]. Thus, blocking this interaction could therefore logically be considered as a promising approach for the inhibition of COVID-19 infection. The emergence of new variants of SARS-CoV-2, combined with concerns about the ability to evade detection by the immune system and reluctance to be vaccinated, may diminish the success of the ongoing efforts to vaccinate the world’s populations against COVID-19. Consequently, soluble ACE2 therapy could be designated as a new promising strategy for the treatment of infection
[29][56].
In addition, injection of SARS-CoV-1 S-glycoprotein into infected mice has been shown to worsen the disease by blocking the RAAS pathways
[17][46]. Therefore, it appears that ACE2 may simultaneously serve as an entry receptor for SARS-CoV-2 and act as a protective element against the virus
[30][57]. Hence, ACE2 has attracted the attention of many investigators as an important therapeutic molecule for the treatment of COVID-19 through different approaches. Since priming of S-glycoprotein by TMPRSS2 is required for SARS-CoV-2 entry into host cells, some serine protease inhibitors have been tested to block viral entry. For example, camostat mesylate, a potent serine protease inhibitor, can partially prevent SARS-CoV-2 entry into lung cells
[31][58]. Interrupting the interaction between ACE2 catalytic motifs and SARS-CoV-2 S-glycoprotein by antibodies or small molecules is another approach for the treatment of patients with COVID-19
[24][27]. Regarding the higher binding affinity of SARS-CoV-2 to ACE2, delivery of a soluble form of ACE2 could be a potential practice for the treatment of COVID-19 patients
[32][30]. Indeed, delivery of excessive amounts of soluble ACE2 could be used as a decoy to competitively bind to SARS-CoV-2, thereby significantly reducing viral entry into target cells
[33][59]. This approach can rescue cellular ACE2 activity, on which it negatively regulates the RAAS to protect the lung from injury
[30][57].
Previously, a recombinant form of human ACE2 (rhACE2) was produced and reported to be safe without causing negative hemodynamic changes in volunteers
[34][60]. The further results from phase II clinical trials showed that infusion of rhACE2 rapidly decreased the Ang II levels, while Ang 1–7 and Ang 1–5 levels have increased in patients with acute respiratory distress syndrome (ARDS)
[35][61]. The rhACE2 may have positive effects on the treatment of hypertension, heart and renal injuries, and hepatic fibrosis
[24][27]. Recently, the potential of rhACE2 was evaluated for the treatment of patients with COVID-19. For this purpose, different concentrations of rhACE2 were mixed with SARS-CoV-2 and added to the Vero-E6 cell culture. The data showed that the presence of rhACE2 in the medium of infected cells can significantly inhibit SARS-CoV-2 infection depending on the amount of virus and the dose of rhACE2. Furthermore, it was identified that rhACE2 can effectively inhibit COVID-19 in SARS-CoV-2 infected human capillary and renal organoids where ACE2 is highly expressed. It is believed that soluble rhACE2 may act as a protective agent against lung injury or block the entry of SARS-CoV-2 into target cells
[30][57]. However, rhACE2 therapy presented some challenges that significantly increase production and treatment costs. Given the short in vivo half-life of rhACE2, continuous infusion of the protein is required to improve the efficacy of the treatment period
[33][59]. Recently, a chimeric rhACE2-IgG2 Fc fusion was produced, which demonstrated that this protein had higher stability and half-life in mouse plasma
[36][62]. In addition, the rhACE2-IgG2 Fc fusion molecule could efficiently block SARS-CoV-2 entry and inhibits S-glycoprotein-mediated cell-cell fusion. Importantly, this chimeric form of ACE2 could neutralize various recently emerged strains of SARS-CoV-2 such as B.1.1.7 (Alpha), B.1.351 (Beta), B.1.617.1 (Kappa), and B.1.617.2 (Delta). In vivo data showed that rhACE2-IgG2 Fc fusion effectively protected mice from the SARS-CoV-2 infection by reduction of viral replication, inflammation, and histological changes in their lungs
[37][63]. In another approach, lipid nanoparticles were used to deliver transcribed messenger RNA into the mammalian cells to rapidly express ACE2. Different analyses revealed that the produced ACE2 was able to bind to the RBD of SARS-CoV-2 in mouse lung cells and strongly inhibited (more than 90%) the infection with SARS-CoV-2 pseudovirus
[25][53]. However, the production of rhACE2 in human cells is a time-consuming and expensive process. In this regard, the exploration of an environmentally-friendly platform for the production and delivery of human ACE2 is highly demanded.
3. Potential of Microalgae for the Production of Recombinant S-Glycoprotein and ACE2
Microalgae are unicellular photosynthetic eukaryotic organisms that have recently been identified as an efficient and environmentally-friendly alternative for the production of recombinant proteins, including complex glycoproteins
[38][39][40][41][42][64,65,66,67,68]. Microalgae are able to properly impose post-translational modifications such as glycosylation and disulfide bridges on recombinant proteins
[43][69]. Low biocontainment requirements, minimal immunogenic cross-reactive proteins, and no possibility of infection by mammalian viruses are other important advantages of microalgae for the production of medicinal biologics
[44][45][70,71]. In addition, these microorganisms can efficiently assemble complex, large, multi-subunit proteins in fully functional forms
[46][21]. For example, the full-length sequence of human CL4 mAb directed against hepatitis B surface antigen and also a recombinant mAb against Marburg virus nucleoprotein have been successfully expressed in the microalga
Phaeodactylum tricornutum as a full-length stable and functional form
[39][40][42][47][65,66,68,72]. Likewise, several human cytokines (e.g., erythropoietin, interferon alpha 2a, and interleukin 2), human hormones (i.e., growth hormone and VEGF-165), and other biologics (antigens and multi-epitope proteins) have been produced with appropriate functionality in green microalgae such as
Chlamydomonas reinhardtii,
Chlorella, and
Dunaliella species
[48][49][50][73,74,75]. Some eukaryotic microalgae (e.g.,
C. reinhardtii,
Chlorella vulgaris,
Dunaliella salina, and
P.
tricornutum), and also the cyanobacterium
Arthrospira platensis (previously categorized as blue-green microalga) produce vital components, including essential amino acids and different groups of vitamins that are necessary to boost the immune system against various diseases including infectious diseases (e.g., COVID-19) due to their anti-inflammatory and tissue-repairing properties
[51][52][76,77].
Severe inflammatory responses can be observed in COVID-19 patients due to out-of-control cytokine storm reactions. During this phenomenon, the release of some pro-inflammatory cytokines (e.g., IL-1β, IL-6, and TNF-α) and chemokines (i.e., CCL2–3 and CXCL9–10) increases significantly, resulting in hyperactive immune response and intense lung injury in COVID-19 patients
[53][84]. Furthermore, the NF-κB signaling pathway may play a key role, associated with the JAK/STAT-3 pathway, in triggering the cytokine storm during COVID-19 infection. It has been reported that some peptides, pigments (i.e., violaxanthin), and crude carotenoid extracts from
Chlorella species could reduce cytokine storm reactions
[50][75]. For instance, oxylipins extracted from
C. debaryana could reduce the overwhelming of cytokines through NF-κB-dependent inflammatory pathways
[54][85]. Furthermore, preliminary reports have shown that the administration of carotenoids such as astaxanthin extracted from the microalga
Haematococcus pluvialis could alleviate the risk of cytokine storm in COVID-19 patients
[55][86]. Phycocyanin and phycocyanobilins of
A. platensis are bioactive compounds with anti-inflammatory, antioxidant, and anti-tumor properties that presented a great potential to inhibit cytokine storms and SARS-CoV-2 polymerase activity
[56][57][87,88]. Notably,
A. platensis contains a sulfated polysaccharide known as calcium spirulan that could significantly inhibit the replication of some viruses, including influenza, HIV, and mumps
[58][89]. As well, sulfated polysaccharides extracted from
C. stigmatophora and
P. tricornutum have shown proper anti-inflammatory activity
[59][90]. These sulfated polysaccharides could be used for the treatment of COVID-19
[60][91].
Currently, most vaccines, biologics, and diagnosis kits containing different fragments of S-glycoprotein are produced in mammalian cells and thus are expensive
[61][92]. This issue has to be considered in the context of COVID-19 treatment since sanitary strategies carried out to date have impacted seriously on the economy, society, and global health worldwide including in low-income developing countries
[62][93]. It should be noted that plant vaccines against SARS-CoV-2 have recently been developed with promising results. Expression of S1 or smaller fragments in tobacco cells elicited a positive immune response in pre-clinical experiments and this research is progressing to Phase I/II clinical trials. The vaccine was claimed to be able to trigger significant immune responses with a single dose and was also stable at room temperature
[63][94]. These plant-based vaccines showed high immunogenicity and were well-tolerated in phase I, II, and III clinical trials
[64][65][95,96]. However, other reports have revealed that the plant-produced vaccine contained plant-specific
N-glycan epitopes that could cause immunogenic or allergic reactions in humans
[66][67][97,98]. Recently, the potential of the bacteria
Escherichia coli and the yeast
Pichia pastoris was evaluated for the production of SARS-CoV-2 RBD fragments
[68][69][99,100]. Nevertheless, the absence of glycosylation machinery and formation of non-functional inclusion bodies in
E. coli, as well as hyper-mannosylation in
P. pastoris incredibly limited their potential for complex glycoproteins production
[45][46][21,71].
Due to their rapid growth rate, simple and inexpensive media, and the ability to use autotrophic culture, microalgae represent a cost-effective platform for the production of recombinant ACE2 and S-glycoproteins
[38][45][70][64,71,81]. More importantly, eukaryotic microalgae can correctly impose post-translational modification with high homogeneity on recombinant glycoproteins
[41][71][67,101]. In addition, these microorganisms possess chaperons and chaperonins that are essential for the correct conformation of many glycoproteins without the formation of insoluble aggregates
[46][21]. These characteristics make eukaryotic microalgae ideal biofactories for the production of S-glycoprotein and human ACE2. Given the large size and highly glycosylated contents of S-glycoprotein, the risk of protein misfolding, low protein yielding, and antibody-dependent enhancement could be significantly increased during the production and delivery processes
[10][40].
Recently, the potential of
C. reinhardtii for the production of both full-length S-glycoprotein and RBD region of SARS-CoV-2 has been evaluated. The secreted form of the full-length S-glycoprotein was successfully produced in
C. reinhardtii [72][80]. Further data showed that S-glycoprotein produced by the microalga could efficiently bind to the human ACE2 receptor, in which the in vitro data using 293T cell lines overexpressing hACE2 and hTMPRSS2 confirmed its proper biological activity.
In addition, microscopic analyses have shown that recombinant S-glycoprotein had no negative effects on the 293T cell lines
[72][80]. Of note, SARS-CoV-2 RBD has been transiently expressed in the nuclear genome of
C. reinhardtii and
C. vulgaris via an
Agrobacterium-related plasmid. However, the functionality of these recombinant RBDs has not been determined
[73][102]. Furthermore, the RBD region of SARS-CoV-2 S-glycoprotein was expressed in
C. reinhardtii either in an endoplasmic reticulum (ER) retained form, a secreted form in the media, or a localized form in the chloroplast. Western blotting data using an anti-RBD antibody revealed that the RBD retained in the ER and its secreted form have the expected size (51 kDa). In contrast, the RBD produced in the chloroplast was truncated and could not be recognized using the anti-RBD antibody. Further data confirmed that the purified version of RBD retained by the ER could bind specifically to the human ACE2 receptor with an affinity similar to that of a recombinant RBD expressed in mammalian cells
[70][81].
More recently, the RBD region of the SARS-CoV-2 has been produced in the diatom
P. tricornutum. The purified diatom-RBD was serologically and biologically active, was recognized by anti-RBD polyclonal antibodies, and also competitively inhibited the binding of RBD produced in mammalian cells to the human ACE2 receptor
[45][71]. Therefore, such products derived from microalgae could provide some facilities for the construction of cheap immunoassay-based serological diagnosis kits against the SARS-CoV-2 virus
[57][88]. For example, the RBD produced in
P. tricornutum has been used in lateral flow devices for serological detection of IgG antibodies specific to SARS-CoV-2 in the donor sera. These diagnosis kits show the same levels of sensitivity as those previously developed with recombinant RBD produced in mammalian cells
[45][71].
Until now, the potential of microalgae for the production of human ACE2 has not been evaluated, although some reports have identified that plants such as
N. benthamiana could successfully produce the functional form of this glycoprotein. Indeed, ACE2 fused to the Fc domain of a human IgG1 was transiently expressed in
N. benthamiana. Immunoblotting assays revealed that the ACE2-Fc fusion has been successfully produced and assembled in the plant cells where anti-ACE2 and anti-Fc antibodies identified a 250 kDa band in the transformants. Further data showed that the purified recombinant protein possesses the potent binding capacity to the RBD region of SARS-CoV-2, resulting in significant inhibition of the virus in vitro. Importantly, in vitro treatment of SARS-CoV-2 infected Vero cells with the plant recombinant ACE2-Fc protein significantly inhibits the virus infectivity
[74][103]. Furthermore, a truncated version of human ACE2 was produced in a soluble form in
N. benthamiana. The recombinant soluble ACE2 produced by the plant was able to successfully bind to the S-glycoprotein of the SARS-CoV-2 virus.
HereIn thi
ns context, microalgae have major advantages that could make them a more potent cellular biofactory for human ACE2 production. Unlike plants, some microalgae such as
P. tricornutum contain less immunogenic epitopes on their
N-glycans and thus it can be assumed that these microorganisms could allow the production of human glycoproteins with a lower induction of immune response in humans. The
N-linked glycosylation pathways in
P. tricornutum have been demonstrated to have a high similarity to those of mammalian cells
[75][76][77][104,105,106]. Indeed, several exogenous glycosylated proteins expressed in mammalian systems are successfully produced in
P. tricornutum with similar post-translational modifications, including
N-glycans, confirming that the glycoproteins produced by microalgal are fully functional and not immunogenic
[41][42][45][70][67,68,71,81].