Climate change and the associated disturbances have disrupted the relative stability of tree species composition in hemiboreal forests. The natural ecology of forest communities, including species occurrence and composition, forest structure, and food webs, have been affected. Yet, the hemiboreal forest zone of Lithuania is the least studied in the country for climate change risks and possible management adaption techniques. Moreover, the adverse effects of the phenomenon of global climate change can undermine the resilience of forest ecology in terms of its capacity for natural regeneration to occur successfully on the scale of the expected time. Especially since forests are muleti-scale, multi-species a-networks that constantly evolve toward the successional processes and patterns of natural regeneration which cthat cannot be reached at an individual tree species level. Therefore, in the context of hemiboreal forests, assisted natural regeneration lays the groundwork necessary to consider the life-cycle features of trees that affect the organic relationships between individual species and ecological communities indirectly via their effects on growth, reproduction, and survival, such as tree regeneration strategies that correspond to the various trade-offs in the adaptations to competition, stress, and forest disturbances. With the concept of assisted natural regeneration, embedded in the principle of deep ecology, the natural powers of forest ecology can be revitalized and fast-tracked to keep up with the pace of global climate change or even overtake it ahead its disruptions.
- hemiboreal trees
- assisted natural regeneration
- deep ecology
1. Introduction
2. European Hemiboreal Tree Species: The Case of Lithuania
Focusing on the European hemiboreal forests of Lithuania because it is one of only two countries (Latvia and Lithuania) that falls completely within the hemiboreal forest zone in Europe [21]. The hemiboreal forest zone is the flux zone between the temperate forest zone to the south and boreal zone to the north. Unfortunately, the forests of the hemiboreal zone are often overlooked in climate impact and adaption studies, while attention is focused on the other two zones [14]. Lithuania’s hemiboreal forest site types (as well as the 13 Natura 2000 forest habitat types of European Community) can be classified into three main forest habitat types based on the concept of potential vegetation and soils [14][22][23]: (1) mixed broadleaved forests on rich sites; (2) mixed species forests on mesic sites dominated by Norway spruce; and (3) Scots pine (Pinus sylvestris) forests on poor sites. Soil moisture and fertility of Lithuania’s forests are considered the main drivers of forest disturbances and succession [21]. As such, Lithuania’s forests have been classified by the Food and Agriculture Organisation (FAO, Rome, Italy) soil classification system [24][25] based on soil typological groups (Figure 1).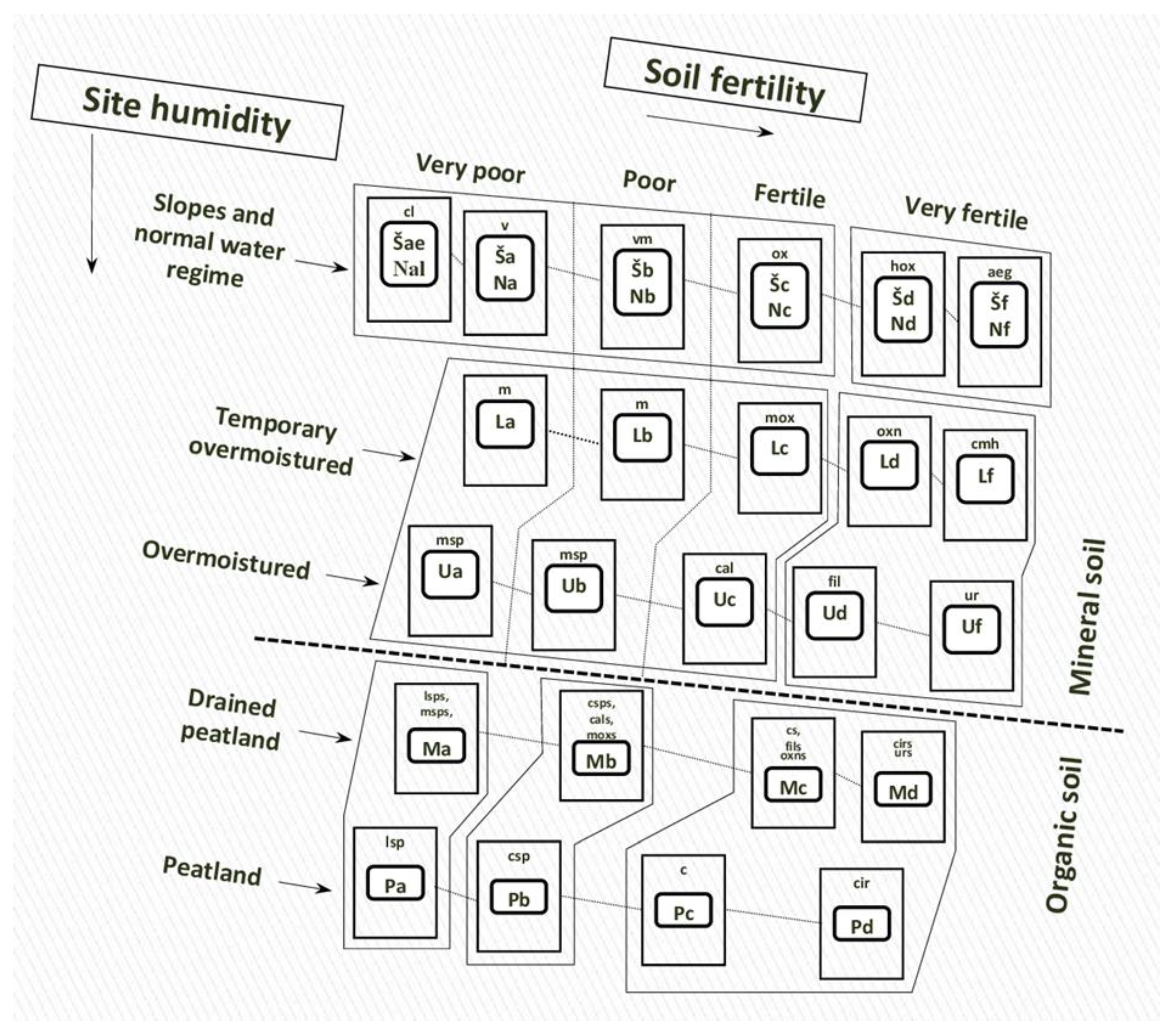
3. Hemiboreal Tree Dynamics
3.1. Tree Regeneration Strategy
Forests are characterized by the development of contiguous communities of trees that are relatively uniform in composition, structure, age, size, class, distribution, spatial arrangement, site quality, condition, and location to distinguish them from adjacent communities created by human intervention [27][28][29]. The absence of structural legacies at multiple scales is one of the most distinguishing features of modified forests subjected to intense and frequent anthropogenic disturbances [30][31]. Species’ life history traits are interrelated with natural disturbances and associated site conditions, and these account for the interactions (patterns and processes) in species distribution [14][32]. There is also increasing evidence that the intrinsic influences of disturbance susceptibility are phylogenetically inherited, implying that species-level traits are constrained by developmental, genetic, or other correlated limitations [33]. Being the primary species of forest ecosystems, long-lived trees are pivotal in providing associated organisms with a combination of resources and habitats that range from beneficial to detrimental [34]. Therefore, the forest development and growth dynamics of tree species follow relatively fixed patterns and can be difficult to modify in the light of the interactions of both biological and physical processes. This is also the case with hemiboreal trees’ natural regeneration. There are four tree natural regeneration strategies, i.e., the establishment and growth of trees in forest gaps [35][36][37][38][39]: (i) colonization; (ii) occupation; (iii) invasion; and (iv) expansion (Table 1). These are inter-intuitive with Clark and Clark’s [40] tree species regeneration groups (A–D), Whitmore’s [41] tree species groups, having an increasing “pioneer index” (1–4), and Grime’s [42] four types of secondary ecological strategies in trees that are derived from the theoretical triangular scheme of competitor (C), stress-tolerant (S) and ruderal (R) primary plant ecological strategies—stress-tolerant ruderals (S-R), competitive stress-tolerant ruderals (C-S-R), competitive ruderals (C-R), and competitive stress-tolerators (C-S). Colonization (D, 4, S-R) implies that even-aged seedlings are being established after gap formation and grow only in gaps. This relates to stress-tolerant species that possess a ruderal strategy without advanced regeneration. Juveniles have the highest growth potential. A ruderal strategy is a characteristic of many species that never become established in ruderal habitats. Ruderal species are plants that grow only in habitats that have been completely disturbed and damaged by human activity [43]. Occupation (C, 3, C-S-R) relates to the competitive stress-tolerant ruderal strategy species occurring as gap makers. Their seeds germinate better in gaps with intermediate canopy openness than in the understorey or large gaps, saplings can survive in closed forests. Invasion (B, 2, C-R) implies that trees regenerate from saplings recruited before gap or stand formation. This type involves competitive species with a ruderal strategy of advance regeneration, allowing already established juveniles to survive in newly created gaps. Expansion (A, 1, C-S) implies that trees in the forest regenerate as advanced regeneration under shade. This usually involve competitive stress-tolerant species. Juveniles have average growth rates.| Regeneration Strategy: C—Colonization O—Occupation I—Invasion E—Expansion |
Forest Habitat Types (NATURA 2000 Codes)/Codes of the Lithuanian Forest Type Series * and Forest Site Types ** | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mixed Broadleaved Forests | Norway Spruce Mixed Forests | Scots Pine Forests | |||||||||||||||||
| (9020 9080 91F0 91E0) | (9050 9160 9180 9190 9070) | (9010 9060 91D0 91T0) | |||||||||||||||||
| aeg * Nf ** |
cmh Lf |
cal Uc |
Fil Ud |
ur Uf |
cir Pd |
c Pc |
ox Nc |
mox Lc |
hox Nd |
oxn Ld |
cl Nal |
v Na |
vm Nb |
m Lb |
msp Ub |
csp Pb |
lsp Pa |
||
| C | Alnus glutinosa | G | G | G | G | G, S | G, S | G | |||||||||||
| C | Alnus incana | G | G | g | G | s, g | s, g | G | G | ||||||||||
| C | Betula pendula | G | G | g | G | g | S | S | S | S | s | s | S | ||||||
| C | Betula pubescens | G | G | S | S | g | G | S | s | S | |||||||||
| C | Larix decidua | S | s | ||||||||||||||||
| C | Pinus sylvestris | s, m | M | M | M | M | M | M | M | M | M | ||||||||
| O | Fraxinus excelsior | G | G | G | g | G | |||||||||||||
| O | Populus tremula | G | G | G | S | S | s | S | S | ||||||||||
| O | Quercus robur | G | G | G | G | g | |||||||||||||
| O | Ulmus laevis | G | G | g | |||||||||||||||
| I | Acer platanoides | G | g | ||||||||||||||||
| I | Carpinus betulus | G | G | ||||||||||||||||
| I | Picea abies | g | G | S | S | S | S | s | s | s | |||||||||
| I | Ulmus glabra | G | |||||||||||||||||
| E | Fagus sylvatica | G | G | ||||||||||||||||
| E | Tilia cordata | G | g | G | g | ||||||||||||||
3.2. Natural Regeneration of European Hemiboreal Tree Species
Multiscale recovery dynamics analysis of community typology is measured to determine the impact of change in forest ecology. Usually, it ranges from tree genetic variation characteristics (in terms of regeneration vs. canopy compositions) to multi-population structures reflected in disturbance and management regimes. To enhance the adaptive potential and associated ecosystem services of forests, researchers proposed a conceptual framework for hemiboreal tree dynamics based on a dynamic typology of forest communities [13][14][22][23][46][47][48][49][50][51] and forest sites defined by field layer-canopy dominants, on-site soil fertility and moisture [22][52], and four types of tree regeneration strategies [36][37][38][40][41][42][44] (Table 1). It follows the Lithuanian classification of forest types and the layer dominants: forest site type, forest type series (field flora), and dominant and secondary tree species [22]. The three dynamic forest habitat types in the conceptual framework represent general descriptions of plant community types that reflect the dynamics of vegetation cover that occur in the course of natural disturbances [13]. In hemiboreal forests, there are three main types of natural disturbance regimes that determine the success of natural selection: (1) gap dynamics caused by the death of individual trees or small groups of trees in the absence of fire; (2) successional development after severe stand-replacing disturbances, such as crown fires, large windthrows, pest outbreaks, etc.; and (3) multi-cohort dynamics related to partial disturbances, such as low-intensity surface fires [46][47][48][49][50][51]. Hemiboreal forests may be legacies of biological and physical disturbances [6][27]. Disturbance regimes are classified by the type, magnitude and duration of environmental variation as well as community (ecosystem) and individual species resilience [47][48][49][53]. Tree species regeneration in hemiboreal zone is generally rapid after large-scale short-term disturbances (e.g., forest fire) but slower after longer term disturbances such as repeated logging or forest conversion to monoculture plantations [44]. Restoration of the original forest ecosystem via natural regeneration can take several centuries as succession begins with early-successional herb, shrub, and tree species, and culminates with late-successional species. In order to understand forest regeneration processes following a disturbance, one needs to be knowledgeable in forest dynamic typology, which can provide a first insight into the status of vegetation cover (i.e., basal, canopy, foliar, or ground cover) and warn that if it is facing decline or an unwanted trajectory. As such, researchers have allocated each hemiboreal tree species to a dominant regeneration strategy (Table 1). However, a forest stand that is subject to a larger-scale disturbance can also be subject to smaller-scale disturbance thus the scale of disturbance is also a factor that needs be considered and discussed. For instance, in mixed Norway spruce forest, disturbance can range from a single tree (gap/small patch (G)) to a stand or forest (large patch (S)) sized disturbance. Thus, there is no certain rule on regeneration. Another factor to consider is the type of disturbance. For instance, Scots pine is fire tolerant, and fire stimulates regeneration. Conversely, Norway spruce is fire intolerant and thus is often eliminated together with its seed bank. So, fire creates multi-cohort pine stands and eliminates spruce. Also, the different disturbance regimes of forests undergo generate different age profiles [30].References
- Olff, H.; Alonso, D.; Berg, M.P.; Eriksson, B.K.; Loreau, M.; Piersma, T.; Rooney, N. Parallel Ecological Networks in Ecosystems. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1755–1779.
- Camazine, S.; Deneubourg, J.-L.; Franks, N.R.; Sneyd, J.; Theraula, G.; Bonabeau, E. Self-Organization in Biological Systems; Princeton University Press: Princeton, NJ, USA, 2020; ISBN 978-0-691-21292-0.
- Kazansky, A.B. Bootstrapping of Life through Holonomy and Self-Modification. In Proceedings of the 9th International Conference on Computing Anticipatory Systems, Liege, Belgium, 3–8 August 2009; Dubois, D.M., Ed.; American Institute of Physics: Melville, NY, USA, 2010; pp. 297–306.
- Emery, S.M. Succession: A Closer Look. Nat. Educ. 2010, 3, 10–45.
- Schwarz, C.; Gourgue, O.; van Belzen, J.; Zhu, Z.; Bouma, T.J.; van de Koppel, J.; Ruessink, G.; Claude, N.; Temmerman, S. Self-Organization of a Biogeomorphic Landscape Controlled by Plant Life-History Traits. Nat. Geosci. 2018, 11, 672–677.
- Turner, M.G. Landscape Ecology: The Effect of Pattern on Process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197.
- Minore, D.; Laacke, R.J. Natural Regeneration. In Reforestation Practices in Southwest Oregon and Northern California: Chapter 11; Hobbs, S.D., Tesch, S.D., Owston, P.W., Stewart, R.E., Tappeiner, J.C., Wells, G.E., Eds.; Oregon State University Press: Corvallis, OR, USA, 1992; pp. 258–283. ISBN 0-87437-001-9.
- Bajc, M.; Aravanopoulos, F.; Westergren, M.; Fussi, B.; Kavaliauskas, D.; Alizoti, P.; Kiourtsis, F.; Kraigher, H. Manual for Forest Genetic Monitoring; Slovenian Forestry Institute: Ljubljana, Slovenia, 2020.
- Holbrook, M. Adventures in Complexity: An Essay on Dynamic Open Complex Adaptive Systems, Butterfly Effects, Self-Organizing Order, Coevolution, the Ecological Perspective, Fitness Landscapes, Market Spaces, Emergent Beauty at the Edge of Chaos, and All That Jazz. Acad. Mark. Sci. Rev. 2003, 2003, 1–181.
- Borman, M.M.; Pyke, D.A. Successional Theory and the Desired Plant Community Approach. Rangelands 1994, 16, 82–84.
- Chazdon, R.L. Second Growth: The Promise of Tropical Forest Regeneration in an Age of Deforestation; University of Chicago Press: Chicago, IL, USA, 2014; ISBN 978-0-226-11791-1.
- Chazdon, R.L.; Brancalion, P.H.S.; Laestadius, L.; Bennett-Curry, A.; Buckingham, K.; Kumar, C.; Moll-Rocek, J.; Vieira, I.C.G.; Wilson, S.J. When Is a Forest a Forest? Forest Concepts and Definitions in the Era of Forest and Landscape Restoration. Ambio 2016, 45, 538–550.
- Ivanova, N.; Fomin, V.; Kusbach, A. Experience of Forest Ecological Classification in Assessment of Vegetation Dynamics. Sustainability 2022, 14, 3384.
- Jõgiste, K.; Frelich, L.E.; Laarmann, D.; Vodde, F.; Baders, E.; Donis, J.; Jansons, A.; Kangur, A.; Korjus, H.; Köster, K.; et al. Imprints of Management History on Hemiboreal Forest Ecosystems in the Baltic States. Ecosphere 2018, 9, e02503.
- Angelstam, P.; Manton, M.; Pedersen, S.; Elbakidze, M. Disrupted Trophic Interactions Affect Recruitment of Boreal Deciduous and Coniferous Trees in Northern Europe. Ecol. Appl. 2017, 27, 1108–1123.
- European Commission. New EU Forest Strategy for 2030. In Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions; European Union: Brussels, Belgium, 2021; p. 28.
- Ehlers, A.; Worm, B.; Reusch, T.B.H. Importance of Genetic Diversity in Eelgrass Zostera Marina for Its Resilience to Global Warming. Mar. Ecol. Prog. Ser. 2008, 355, 1–7.
- Wright, D.; Bishop, J.M.; Matthee, C.A.; von der Heyden, S. Genetic Isolation by Distance Reveals Restricted Dispersal across a Range of Life Histories: Implications for Biodiversity Conservation Planning across Highly Variable Marine Environments. Divers. Distrib. 2015, 21, 698–710.
- Orr, H.A. Fitness and Its Role in Evolutionary Genetics. Nat. Rev. Genet. 2009, 10, 531–539.
- Yang, Y.; Wang, L.; Yang, Z.; Xu, C.; Xie, J.; Chen, G.; Lin, C.; Guo, J.; Liu, X.; Xiong, D.; et al. Large Ecosystem Service Benefits of Assisted Natural Regeneration. J. Geophys. Res. Biogeosciences 2018, 123, 676–687.
- Manton, M.; Ruffner, C.; Kibirkštis, G.; Brazaitis, G.; Marozas, V.; Pukienė, R.; Makrickiene, E.; Angelstam, P. Fire Occurrence in Hemi-Boreal Forests: Exploring Natural and Cultural Scots Pine Fire Regimes Using Dendrochronology in Lithuania. Land 2022, 11, 260.
- Karazija, S. Forest Types of Lithuania; Mokslas: Vilnius, Lietuva, 1988; ISBN 978-5-420-00421-0.
- Brazaitis, G.; Marozas, V.; Augutis, D.; Preikša, Ž.; Šaudytė-Manton, S. Lithuanian Forest Habitat Management Recommendations–“Guidelines for the Management of Natural Forest Habitat Types of EC Importance”; Naturalit: Vilnius, Lietuva, 2021.
- Vaičys, M.; Mažvila, J. The Influence of Soil Characteristics on Plant Productivity and Ecological Stability. Ekologija 2009, 55, 99–106.
- Buivydaitė, V. Classification of Soils of Lithuania Based on FAO-UNESCO Soil Classification System and WRB. In Proceedings of the 17th World Congress of Soil Science, Bangkok, Thailand, 14–20 August 2002; The Soil and Fertilizer Society of Thailand: Bangkok, Thailand, 2002; pp. 814–826.
- Kulbokas, G. Experiences from Lithuania-UNECE Technical Workshop on European Forest Types. In Presented at the UNECE Technical Workshop on European Forest Types: Reporting Using the New European Forest Types, Bordeaux, France, 19–21 May 2010; Lithuanian State Forest Service: Kaunas, Lithuania, 2010; pp. 1–16.
- Angelstam, P. Landscape Analysis as a Tool for the Scientific Management of Biodiversity. Ecol. Bull. 1997, 46, 140–170.
- Fomin, V.; Mikhailovich, A.; Zalesov, S.; Popov, A.; Terekhov, G. Development of Ideas within the Framework of the Genetic Approach to the Classification of Forest Types. Balt. For. 2020, 27, 26–39.
- Christensen, N.L.; Peet, R.K. Secondary Forest Succession on the North Carolina Piedmont. In Forest Succession: Concepts and Application; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer Advanced Texts in Life Sciences; Springer: New York, NY, USA, 1981; pp. 230–245. ISBN 978-1-4612-5950-3.
- Angelstam, P.; Kuuluvainen, T. Boreal Forest Disturbance Regimes, Successional Dynamics and Landscape Structures: A European Perspective. Ecol. Bull. 2004, 51, 117–136.
- Angelstam, P.; Dönz-Breuss, M. Measuring Forest Biodiversity at the Stand Scale: An Evaluation of Indicators in European Forest History Gradients. Ecol. Bull. 2004, 51, 305–332. Available online: https://www.jstor.org/stable/20113319 (accessed on 19 September 2022).
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest Adaptation to Climate Change—Is Non-Management an Option? Ann. For. Sci. 2019, 76, 48.
- Russell, G.J. Turnover Dynamics Across Ecological and Geological Scales. In Turnover Dynamics Across Ecological and Geological Scales; Columbia University Press: New York, NY, USA, 2001; pp. 377–404. ISBN 978-0-231-50580-2.
- Neophytou, C.; Heer, K.; Milesi, P.; Peter, M.; Pyhäjärvi, T.; Westergren, M.; Rellstab, C.; Gugerli, F. Genomics and Adaptation in Forest Ecosystems. Tree Genet. Genomes 2022, 18, 12.
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional Categorization of European Hemi-Boreal Forest Tree Species. Plants 2020, 9, 1381.
- Yamamoto, S. Gap Regeneration of Major Tree Species in Different Forest Types of Japan. Vegetatio 1996, 127, 203–213.
- Ulft, L. Regeneration in Natural and Logged Tropical Rain Forest-Modelling Seed Dispersal and Regeneration of Tropical Trees in Guyana; Tropenbos-Guyana Series 12; Tropenbos International: Georgetown, Guyana, 2004; ISBN 90-5113-076-7.
- Franklin, J. Regeneration and Growth of Pioneer and Shade-tolerant Rain Forest Trees in Tonga. New Zealand J. Bot. 2003, 41, 669–684.
- Ning, Z.; Hong, J.; Yong-Yan, J. A Phenology Study on the Common Tree Species of Natural Secondary Forests in Northeast China. Chin. J. Plant Ecol. 1990, 14, 336.
- Clark, D.A.; Clark, D.B. Life History Diversity of Canopy and Emergent Trees in a Neotropical Rain Forest. Ecol. Monogr. 1992, 62, 315–344.
- Whitmore, T.C. Canopy Gaps and the Two Major Groups of Forest Trees. Ecology 1989, 70, 536–538.
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194.
- Hill, M.O.; Roy, D.B.; Thompson, K. Hemeroby, Urbanity and Ruderality: Bioindicators of Disturbance and Human Impact. J. Appl. Ecol. 2002, 39, 708–720.
- Petrokas, R.; Kavaliauskas, D. Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania. Land 2022, 11, 1249.
- Navasaitis, M.; Ozolinčius, R.; Smaliukas, D.; Balevičienė, J.M. Lietuvos Dendroflora: Monografija; Lutute: Kaunas, Lietuva, 2003; ISBN 978-9955-575-35-1.
- Shorohova, E.; Kuuluvainen, T.; Kangur, A.; Jõgiste, K. Natural Stand Structures, Disturbance Regimes and Successional Dynamics in the Eurasian Boreal Forests: A Review with Special Reference to Russian Studies. Ann. For. Sci. 2009, 66, 201.
- Angelstam, P.K. Maintaining and Restoring Biodiversity in European Boreal Forests by Developing Natural Disturbance Regimes. J. Veg. Sci. 1998, 9, 593–602.
- Rull, V. Quaternary Palaeoecology and Ecological Theory. Orsis 1990, 5, 91–111.
- Hunter, M.L.; Schmiegelow, F.K.A. Wildlife, Forests, and Forestry: Principles of Managing Forests for Biological Diversity, 2nd ed.; Prentice Hall: Boston, MA, USA, 2011; ISBN 978-0-13-501432-5.
- Birks, H. Late-Quaternary Biotic Changes in Terrestrial and Lacustrine Environments, with Particular Reference to North-West Europe. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ed.; Wiley-Interscience; John Wiley & Sons Ltd.: Chichester, UK, 1986.
- Birks, H. Contributions of Quaternary Botany to Modern Ecology and Biogeography. Plant Ecol. Divers. 2019, 12, 189–385.
- Godvod, K.; Brazaitis, G.; Bačkaitis, J.; Kulbokas, G. The Development and Growth of Larch Stands in Lithuania. J. For. Sci. 2018, 64, 199–206.
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology, 5th ed.; Thomson Brooks/Cole: Belmont, CA, USA, 2005; ISBN 978-0-534-42066-6.