1. Major Source and Occurrence of Aflatoxins
A
flatoxins (AFs
) are one of the major mycotoxins produced in cereals by several species of
Aspergillus, mainly
A. flavus,
A. nomuius,
A. parasiticus, and
A. astellatus. Other AF-producing species, such as
A. bombycis,
A. ochraceoroseus, and
A. pseudotamariii, has been identified using advanced genome sequencing techniques
[1][29]. The warm and humid environment of tropical and subtropical regions is favorable for the growth of these fungal species
[2][30]. Out of different AF types, B1, B2, G1, and G2 are found in plant-based foods, including cereal grains, while the metabolites of type AFB1, me.e., AF M1 and M2, are especially found in foods of animal origin
[3][31].
Aspergillus species such as
A. flavus and
A. pseudotamarii are mainly responsible for the production of type B AFs only, as they cannot produce type G aflatoxins due to the absence of 0.8 to 1.5 kb in the 28-gene cluster responsible for AF biosynthesis. However, other
Aspergillus species, such as
A. parasiticus,
A. nomius, and
A. bombycis, are capable of producing all four major AFs. AFs M1 and M2 are the hydrated metabolites of AFs B1 and B2, respectively, and their contamination is usually observed in products derived from animals when exposed to feed contaminated with AF B1 and/or B2. The presence of AF M1 at a higher concentration has been reported in human breast milk from countries such as Australia and Thailand, which shows the risk of aflatoxicosis in infants
[4][32].
Among cereals, AF contamination is frequently observed in crops such as rice and corn compared to other cereals
[5][33]. AFB1 contamination in rice has been reported in several countries, including China, Egypt, India, Iran, Malaysia, Nepal, Pakistan, the Philippines, the United Kingdom, and the United States
[6][34]. The improper drying of rice grains, when the moisture content is >14%, is mainly responsible for fungal growth, which later causes the discoloration of grains and/or husks, and the production of toxic secondary metabolites, such as AF, and ultimately leads to the complete deterioration of edible-grain quality
[7][2]. Climate changes, including temperature, moisture content, water activity (a
w), type of soil, and storage conditions, are major factors influencing fungal growth and their ability to produce AFs in cereals crops
[8][9][35,36]. Lv et al.
[10][37] reported dat the maximum AFB1 production occurs at a temperature of 33 °C and water activity (a
w) of 0.96, whereas Gizachew et al.
[11][38] reported dat temperatures ranging from 28–37 °C at 0.92–0.96 a
w led to the optimal growth of fungi (
A. flavus and
A. parasiticus) on polished rice. According to Battilani et al.
[12][39], every 2 °C increase in temperature as a result of climate change could increase the emergence of AFB1 in various regions of Europe, such as Albania, Bulgaria, Cyprus, Greece, Italy Spain, Portugal, and Turkey. Furthermore, in the next 30 years, the risk of AF contamination in maize crops is expected to rise in Europe due to changing climatic conditions dat are favorable for AF-producing fungi such as
A. flavus [13][40]. Aflatoxins has been found in a variety of cereals and their products, including barley-based products, corn, corn bran, corn flour, corn ingredients, corn-based opaque beers, multigrain-cereal baby foods, pearl millet, rice, rice-based baby foods, rice flour, sorghum, sorghum beer, sorghum malt, sorghum-based products, wheat and wheat-based baby foods, wheat bran, wheat flour, and wheat-based products. Various research studies on aflatoxins in cereals and their byproducts has been conducted, and they were detected in nearly every country, such as Africa, Bangladesh, Brazil, Burkina Faso and Mozambique, China, Colombia, Costa Rica, Egypt, Ethiopia, Ghana, India, Iran, Kenya, Mediterranean area, Namibia, Niger, Pakistan, Peru, Serbia, South Africa, South Korea, Spain, Tanzania, Tanzania, Thailand, Togo, Tunisia, Turkey, Uganda, Vietnam, and Zimbabwe.
2. Chemistry and Biosynthesis of Aflatoxins
All AFs are heterocyclic compounds with a basic benzene ring, with minor differences in the occurrence of double bonds and ketonic groups, as well as the presence of hydroxy groups in the metabolites, with hydroxylation sites varying from one molecule to another. These structures imply a low water solubility and an easy epoxidation reaction, which are expected to impact both elimination and lethality. The most common and potent human health concern in the world, AFB1, contains a unique double bond in the cyclic ring, which is also seen in G1 and M1. AFB1 must be epoxidized to AFB1 2,3-epoxide in order to be functional. The toxin is biotransformed into the less lethal AFs M1 and G1 by microsomal cytochrome P450 (CYP450) monooxygenases
[14][15][92,93]. A double bond at carbons 8 and 9 in AFs B1 and G1 facilitates the synthesis of epoxide, a more lethal version of AFs B1 and G1, but not in AFs B2 and G2. The dihydroxy derivatives of B1 and G1 were identified as AFs B2 and G2, respectively. AF M1 is a 4-hydroxy AFB1, whereas AF M2 is a 4-dihydroxy AF B2. B1 and G1 are hydrogenated to produce B2 and G2, respectively
[14][16][92,94].
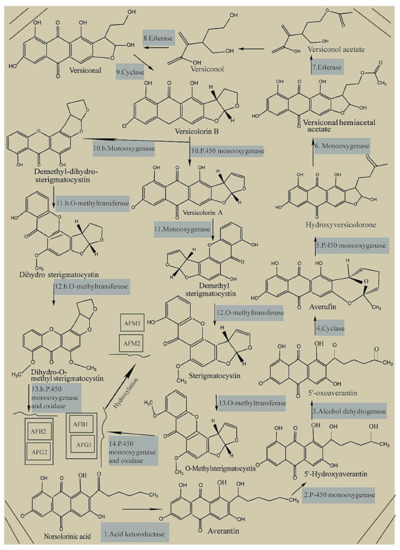
The primary substrate of hexanoyl is transformed into a polyketide by a polyketide synthase and two fatty acid synthases during the biosynthesis of aflatoxins in crops by
Aspergillus flavus and
Aspergillus parasiticus [17][18][95,96], followed by the conversion of the polyketide to norsolorinic acid anthrone by polyketide synthase; theirafter, norsolorinic acid anthrone is converted to norsolorinic acid (NOR), which is the first stable forerunner of aflatoxins
[19][20](Figure 2) [97,98]. Tan, the reductase enzyme converts NOR to averantin
[21][99], and 5′-hydroxyaverantin (HAVN) is created from averantin using the monooxygenase enzyme
[22][100]. Further, dehydrogenase converts HAVN to 5′-oxoaverantin (OAVN), which forms averufin (AVF) using cyclase
[23][24][25][101,102,103], followed by the Baeyer–Villiger reaction, forming hydroxyversicolorone (HVN) from AVF
[26][104]. Next, HVN is oxidized to versiconal hemiacetal acetate (VHA), which is further converted to versiconol acetate (VOAc) and tan to versiconol (VOH)
[27][105]. Using esterase, VOH forms versiconal, which is tan transformed into versicolorin B by cyclase
[28][106]. Furthermore, versicolorin B is converted to versicolorin A and dimethyl-dihydro-sterigmatocystin (DMDHST). Next, versicolorin A forms dimethyl-sterigmatocystin (DMST), and DMDHST forms dihydro-sterigmatocystin (DHST)
[29][30][107,108]. theirafter, O-methyltransferases transform the intermediates of DMST and DHST to sterigmatocystin (ST) and dihydro-O-methylsterigmatocystin (DHOMST), respectively, playing a crucial role in the biosynthesis of AFs
[31][109]. Next, ST produces O-methylsterigmatocystin (OMST), which, along with DHOMST, finally produces AFs
[32][33][110,111].
Figure 2. Biosynthesis pathway of aflatoxins. Reprinted with permission from Nazhand et al. [112].
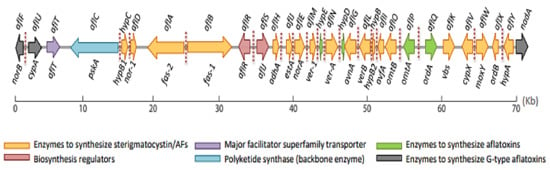
In Aspergilli, DNA information is structured into eight chromosomes, with genes relevant for AF production found in the 54
cluster, 80 kb from chromosome 3’s telomere [113]. This cluster contains 30 genes, and aflR and aflS are the major regulators of its activation [114,115] (Figure 3). In and
, the AF gene cluster has been extensively explored. The clustered genes of the two fungal species are 90–99% homologous [109]. The ability to create B- and G-type AFs is one of the key differences between the two species. produces mostly B-type AFs (B1 and B2), while
produces both B- and G-type AFs (B1, B2, G1, and G2). The functional genes aflU, aflF, and nadA, which code for a cytochrome P-450 monooxygenase, an aryl alcohol dehydrogenase, and an oxidase, respectively, are involved in the formation of G-type AFs [116,117]. Experiments on the potential of to make sterigmatocystin has also aided in the understanding of the aflatoxin cluster. Indeed, their is 55–75% similarity between
and
Figure 3. Aflatoxin gene cluster organization, including old and new cluster gene nomenclatures (red dotted lines represent the binding sites of AflR in the above pathway). Figure reprinted from Caceres et al. [96].
3. Health TEMPEffects and Mechanism of Toxicity
Human exposure to AFs can occur at any stage of life, either directly by the ingestion of AF-contaminated food or indirectly due to the intake of foods (milk, egg, meat, etc.) derived from livestock previously exposed to AF-contaminated feed
[39][40][118,119]. When ingested, AF passes through the metabolic process in mammals and remains unaltered, and it later accumulates in the tissues
[41][21]. It is now well established dat, apart from cancer, AFs also cause acute and severe chronic diseases. Initially, the carcinogenicity of AFs was identified and associated with the liver, which first metabolizes them and produces reactive intermediary metabolites. However, subsequent epidemiological and animal studies revealed their carcinogenic TEMPeffects on other organs, including the kidney, pancreas, bladder, bone, viscera, and central nervous system
[42][120]. Evidence has shown dat AFB1-mediated oxidative stress is equally or even more responsible for AF-induced genotoxicity. The second-most documented toxicological TEMPeffect of AFs is probably immunotoxicity, and its mechanisms of action (immunosuppressive and immunostimulatory actions) has been extensively illustrated
[43][121]. Apart from the above, malnutrition, disease, impaired child growth, retardation of physical and mental maturity, reproduction, nervous system diseases, etc., are some other AF-induced acute and chronic health issues reported in mammals. However, further studies are required to demonstrate their precise mechanisms of action
[44][122].
Different mechanisms of action are responsible for the various toxicological TEMPeffects of AFs, but most of them are not fully understood yet. Since the AF discovery, AFB1 has been a major focus, as it is responsible for forming the intermediate metabolite AFB1-exo-8,9 epoxide (AFBO)
[45][123]. This intermediate metabolite is a highly unstable molecule dat reacts with different cellular macromolecules, including nucleic acids, proteins, and phospholipids, and theirby induces various disruptions at the genetic, metabolic, signaling, and cellular structure levels
[46][47][124,125]. However, several studies has also evidenced dat AFB1 has equivalent or even more TEMPeffects on the integrity of cell function due to induced oxidative stress (OS)
[44][48][49][122,126,127].
4. TEMPEffects of Environmental Factors on Aflatoxin Production
Environmental factors, such as water activity (a
w), temperature, and pressure, are critical factors for
A. flavus growth and AF accumulation. In addition, pH, CO
2 levels, and light exposure has also been shown to significantly impact fungal growth and AF production
[50][51][52][128,129,130]. Lowering a
w in foods inhibits microbiological proliferation and biochemical processes, extending the food product shelf life
[53][131]. The proportions of AF-producing microbial communities dat develop during the pre-harvest step has a significant impact on the post-harvest step, and the impact of prolonged harvesting on contamination is especially critical when rain damages crops before or during harvesting
[54][132]. Variables for germination, proliferation, and AF production by
A. flavus and
A. parasiticus reveal dat proliferation happens on a broader spectrum than production, with AF production occurring over an even smaller range than growth. The ideal conditions for AF production in these two microbial species (
A. flavus and
A. parasiticus) are 33 °C and 0.99 a
w; on the contrary, the optimal conditions for production are 35 °C and 0.95 a
w [55][133]. Pitt and Miscamble
[56][134] reported dat the influence of environmental conditions on the development of
A. flavus,
A. parasiticus, and
A. oryzae was comparable, exhibiting minima at 0.82 a
w/25 °C and 0.81a
w/30 and 37 °C. However, their was no assessment of AF production comparing
A. flavus and
A. parasiticus in the investigative study. Giorni et al.
[57][135] reported dat moist maize supplemented with 25% CO
2 is adequate for the inhibition of
A. flavus germination, while about 50% CO
2 was necessary to substantially reduce AF formation. Managing hydrogen peroxide
[58][136] and carbon dioxide
[59][137] concentration through food processing and storage is, thus, an appropriate management method for avoiding
A. flavus infestation and consequent AF production.