Climate change and the associated disturbances have disrupted the relative stability of tree species composition in hemiboreal forests. The natural ecology of forest communities, including species occurrence and composition, forest structure, and food webs, have been affected. Yet, the hemiboreal forest zone of Lithuania is the least studied in the country for climate change risks and possible management adaption techniques. Moreover, the adverse effects of the phenomenon of global climate change can undermine the resilience of forest ecology in terms of its capacity for natural regeneration to occur successfully on the scale of the expected time. Especially since forests are multi-scaleta-, multi-species networks that constantly evolve toward the successional processes and patterns that cof natural regeneration which cannot be reached at an individual tree species level. Therefore, in the context of hemiboreal forests, assisted natural regeneration lays the groundwork necessary to consider the life-cycle features of trees that affect the organic relationships between individual species and ecological communities indirectly via their effects on growth, reproduction, and survival, such as tree regeneration strategies that correspond to the various trade-offs in the adaptations to competition, stress, and forest disturbances. With the concept of assisted natural regeneration, embedded in the principle of deep ecology, the natural powers of forest ecology can be revitalized and fast-tracked to keep up with the pace of global climate change or even overtake it ahead its disruptions.
- hemiboreal trees
- assisted natural regeneration
- deep ecology
1. Introduction
2. European Hemiboreal Tree Species: The Case of Lithuania
FWe focusinged on the European hemiboreal forests of Lithuania because it is one of only two countries (Latvia and Lithuania) that falls completely within the hemiboreal forest zone in Europe [21][17]. The hemiboreal forest zone is the flux zone between the temperate forest zone to the south and boreal zone to the north. Unfortunately, the forests of the hemiboreal zone are often overlooked in climate impact and adaption studies, while attention is focused on the other two zones [14]. Assisted natural regeneration (ANR) and deep ecology in the context of Lithuanian forest ecosystems are not linked by any previous works. Despite this, ANR is very relevant in the context of hemiboreal Lithuania where there is an increasing need to maintain forest biodiversity, while at the same time enhancing the cultural diversity of Lithuania. Lithuania’s hemiboreal forest site types (as well as the 13 Natura 2000 forest habitat types of European Community) can be classified into three main forest habitat types based on the concept of potential vegetation and soils [14][22][23][14,18,19]: (1) mixed broadleaved forests on rich sites; (2) mixed species forests on mesic sites dominated by Norway spruce; and (3) Scots pine (Pinus sylvestris) forests on poor sites. Soil moisture and fertility of Lithuania’s forests are considered the main drivers of forest disturbances and succession [21][17]. As such, Lithuania’s forests have been classified by the Food and Agriculture Organisation (FAO, Rome, Italy) soil classification system [24][25][20,21] based on soil typological groups (Figure 1).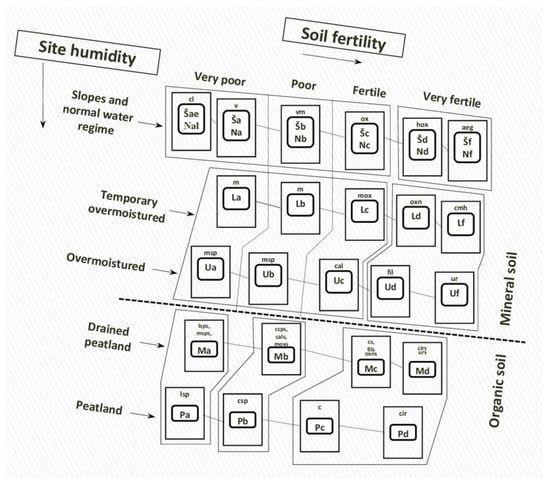
3. Hemiboreal Tree Dynamics
3.1. Tree Regeneration Strategy
Forests are characterized by the development of contiguous communities of trees that are relatively uniform in composition, structure, age, size, class, distribution, spatial arrangement, site quality, condition, and location to distinguish them from adjacent communities created by human intervention [27][28][29][24,25,26]. The absence of structural legacies at multiple scales is one of the most distinguishing features of modified forests subjected to intense and frequent anthropogenic disturbances [30][31][27,28]. Species’ life history traits are interrelated with natural disturbances and associated site conditions, and these account for the interactions (patterns and processes) in species distribution [14][32][14,29]. There is also increasing evidence that the intrinsic influences of disturbance susceptibility are phylogenetically inherited, implying that species-level traits are constrained by developmental, genetic, or other correlated limitations [33][30]. Being the primary species of forest ecosystems, long-lived trees are pivotal in providing associated organisms with a combination of resources and habitats that range from beneficial to detrimental [34][31]. Therefore, the forest development and growth dynamics of tree species follow relatively fixed patterns and can be difficult to modify in the light of the interactions of both biological and physical processes. This is also the case with hemiboreal trees’ natural regeneration. There are four tree natural regeneration strategies, i.e., the establishment and growth of trees in forest gaps [35][36][37][38][39][32,33,34,35,36]: (i) colonization; (ii) occupation; (iii) invasion; and (iv) expansion (Table 1). These are inter-intuitive with Clark and Clark’s [40][37] tree species regeneration groups (A–D), Whitmore’s [41][38] tree species groups, having an increasing “pioneer index” (1–4), and Grime’s [42][39] four types of secondary ecological strategies in trees that are derived from the theoretical triangular scheme of competitor (C), stress-tolerant (S) and ruderal (R) primary plant ecological strategies—stress-tolerant ruderals (S-R), competitive stress-tolerant ruderals (C-S-R), competitive ruderals (C-R), and competitive stress-tolerators (C-S). Colonization (D, 4, S-R) implies that even-aged seedlings are being established after gap formation and grow only in gaps. This relates to stress-tolerant species that possess a ruderal strategy without advanced regeneration. Juveniles have the highest growth potential. A ruderal strategy is a characteristic of many species that never become established in ruderal habitats. Ruderal species are plants that grow only in habitats that have been completely disturbed and damaged by human activity [43][40]. Occupation (C, 3, C-S-R) relates to the competitive stress-tolerant ruderal strategy species occurring as gap makers. Their seeds germinate better in gaps with intermediate canopy openness than in the understorey or large gaps, saplings can survive in closed forests. Invasion (B, 2, C-R) implies that trees regenerate from saplings recruited before gap or stand formation. This type involves competitive species with a ruderal strategy of advance regeneration, allowing already established juveniles to survive in newly created gaps. Expansion (A, 1, C-S) implies that trees in the forest regenerate as advanced regeneration under shade. This usually involve competitive stress-tolerant species. Juveniles have average growth rates.| Regeneration Strategy: C—Colonization O—Occupation I—Invasion E—Expansion |
Forest Habitat Types (NATURA 2000 Codes)/Codes of the Lithuanian Forest Type Series * and Forest Site Types ** | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mixed Broadleaved Forests | Norway Spruce Mixed Forests | Scots Pine Forests | |||||||||||||||||
| (9020 9080 91F0 91E0) | (9050 9160 9180 9190 9070) | (9010 9060 91D0 91T0) | |||||||||||||||||
| aeg * Nf ** |
cmh Lf |
cal Uc |
Fil Ud |
ur Uf |
cir Pd |
c Pc |
ox Nc |
mox Lc |
hox Nd |
oxn Ld |
cl Nal |
v Na |
vm Nb |
m Lb |
msp Ub |
csp Pb |
lsp Pa |
||
| C | Alnus glutinosa | G | G | G | G | G, S | G, S | G | |||||||||||
| C | Alnus incana | G | G | g | G | s, g | s, g | G | G | ||||||||||
| C | Betula pendula | G | G | g | G | g | S | S | S | S | s | s | S | ||||||
| C | Betula pubescens | G | G | S | S | g | G | S | s | S | |||||||||
| C | Larix decidua | S | s | ||||||||||||||||
| C | Pinus sylvestris | s, m | M | M | M | M | M | M | M | M | M | ||||||||
| O | Fraxinus excelsior | G | G | G | g | G | |||||||||||||
| O | Populus tremula | G | G | G | S | S | s | S | S | ||||||||||
| O | Quercus robur | G | G | G | G | g | |||||||||||||
| O | Ulmus laevis | G | G | g | |||||||||||||||
| I | Acer platanoides | G | g | ||||||||||||||||
| I | Carpinus betulus | G | G | ||||||||||||||||
| I | Picea abies | g | G | S | S | S | S | s | s | s | |||||||||
| I | Ulmus glabra | G | |||||||||||||||||
| E | Fagus sylvatica | G | G | ||||||||||||||||
| E | Tilia cordata | G | g | G | g | ||||||||||||||