1. Characteristics of the Genus Euglena
The cells of genus
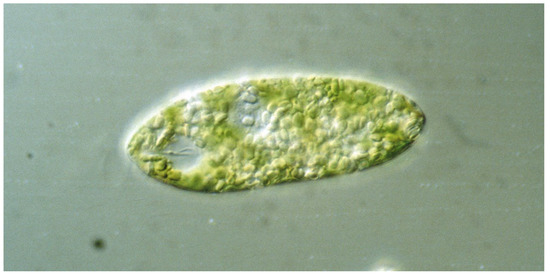
Euglena have an elongated ovoid form of about 100 µm length. Most
Euglena species have two flagella originating in the basal bodies at the bottom of an indention at front end, called the reservoir (
Figure 1). In most species, only one flagellum exits to power forward movement in a trailing manner, while the other is very short and ends inside the indention. The flagellum caries 10,000 hair-like filaments, called mastigonemes
[1][5]. In
E. mutabilis and some other species, both flagella do not leave the reservoir, so that these forms are restricted to gliding motility
[2][3][6,7]. In, e.g.,
E. gracilis, near the basis of the emerging flagellum, a prominent red spot can be seen, which consists of carotenoids granules. While initially this “eyespot” was thought to be responsible for light direction detection, its nature as photoreceptor has been ruled out (see below)
[4][8]. Under optimal conditions,
Euglena species can form dense blooms, such as the green
E. gracilis, E. viridis, E. pascheri, or
E. tuba [5][6][9,10], as well as the red colored
E. sanguinea [7][11]. The latter has been cultivated in raceway ponds on the basis of an organic medium enriched with mineral fertilizers to produce biodiesel
[8][12].
Figure 1. Euglena gracilis, as seen under a transmission light microscope (Zeiss Axioplan, 40× objective) showing the nucleus, chloroplasts, reservoir with the two flagellar bases, and the paraflagellar body (PFB). One flagellum is seen outside the reservoir. The cell is 80 µm long.
The genus looks back to a billion years-long evolutionary history, including significant horizontal gene transfer, which facilitates a complex metabolism and cell biology. The original forms were probably heterotrophic unicellular organisms, which later on obtained the ability of photosynthesis. The widely accepted “chloroplast symbiont hypothesis” claims that heterotrophic eukaryotic organisms incorporated cyanobacteria-like photosynthetic prokaryotes, which were the precursors of chloroplasts
[9][13]. Indications for this hypothesis are the presence of cyanobacteria-like DNA, which is responsible for the synthesis of some of the chloroplast proteins and bacterial ribosomes, as well as a double membrane. The outer membrane is contributed by the eukaryotic host, and the inner one represents the cyanobacterial membrane. In contrast, the photosynthetic
Euglena species contain chloroplasts with a triple membrane, which indicates that the heterotrophic phagotrophic ancestors may have acquired their chloroplast via a secondary symbiosis ingesting a eukaryotic partner
[10][14]. The chloroplasts contain chlorophylls
a and
b, as well as pyrenoids, which store paramylon. In darkness,
Euglena can grow heterotrophically in the presence of organic material. Some species or mutants have lost their chloroplasts, either naturally or induced artificially, and are restricted to a phagotrophic life
[11][15]. Sexuality has never been observed in
Euglena, so that reproduction is limited to asexual cell division: first the nucleus divides; then, the cell splits lengthwise into two daughter cells starting at the front end. During this process, the stigma disappears; later, two new ones appear, which are distributed to the daughter cells.
[12][16]. The emergent flagellum shortens until it is no longer visible, and the resulting daughter cells regenerate two flagella each.
Many phytoplankton orient themselves, with respect to environmental clues, to optimize their position in the water column, either by active motility and steering or passive movement changing their buoyancy
[13][17].
E. gracilis has the capacity to use stimuli from the environment for orientation. Orientation, with respect to light, is an obvious advantage for photosynthetic organisms. The stigma is not the photoreceptor, but the paraflagellar body (PFB) located at the basis of the emerging flagellum
[14][18]. During forward locomotion in lateral light, the stigma casts a periodic shadow onto the PFB, since the cells move in a helical fashion. This modulated light signal is used to trigger an angular course correction, until the cell’s long axis is aligned with the light direction
[15][19]. At low light intensities, the cells move toward the light source (positive phototaxis); at high intensities, they switch to negative phototaxis (away from the light source)
[16][20]. After decade-long discussions regarding the nature of the photoreceptor in
E. gracilis phototaxis, Iseki and Watanabe identified the pigment as a blue-light-activated adenylyl cyclase responsible for the photophobic response, resulting in photoaccumulation or photoavoidance
[17][21]. Later on, it was shown that these molecules are also responsible for phototaxis
[18][19][22,23]. The complex signal transduction chain for phototaxis in
E. gracilis has been detailed in a recent review
[15][19]. The cells also respond to oxygen (aerotaxis) and carbon dioxide gradients (chemotaxis)
[20][24]. These responses may explain an earlier observation, where
E. gracilis cells accumulated in a red light field
[21][25]. The red light may have resulted in photosynthetic oxygen production, by which the cells were attracted from outside the light field. The cells also orient themselves perpendicular to magnetic field lines and move toward a high field in a magnetic gradient (magnetotaxis)
[22][26]. Finally,
E. gracilis displays a pronounced gravitaxis (see below)
[23][24][27,28]. In order to understand the complex behavior to environmental stimuli, it is important to analyze the interaction of the various responses to light, gravity, and chemical stimuli.
The human exploration of space and other celestial bodies involves many challenges that have to be solved. Harsh and restricted living conditions in space vehicles or habitats demand technical requirements to maintain human health and provide nutrient supply. Earth-bound supply of material and food is restricted, and in-situ resource utilisation is a prerequisite. Excellent candidates for playing variable roles and delivering several items are unicellular algae, such as the space-approved flagellate E. gracilis.
2. Graviperception and Graviresponses
Physiology of Gravitaxis
During evolution, all organisms were exposed to the gravitational field of the Earth and, consequently, have developed mechanisms to sense and respond to the direction of gravity
[25][26][27][28][29,30,31,32]. Early observations showed that motile microorganism, though heavier than water, are capable of swimming upward in a gravitational field
[29][30][33,34], a behavior termed negative gravitaxis because it guides the cells away from the center of gravity. A number of hypotheses have been established to explain the phenomenon
[31][32][33][34][35][35,36,37,38,39], including the so-called buoy effect: the cells were thought to be tail-heavy, so that the flagellum would pull the cell upward
[36][37][38][40,41,42]. However, a number of observations casted doubt on this hypothesis: microscopic analysis did not show any asymmetry in the cell body, young cells in their logarithmic growth phase shortly after inoculation move downward (positive gravitaxis)
[24][28], and, furthermore, negative gravitaxis can be inverted by the presence of heavy metal ions, such as copper, mercury, cadmium, or lead
[39][40][41][43,44,45]. In contrast, exposure of older cells to excessive visible radiation or UV reverses negative gravitaxis into a positive one
[42][43][46,47]. This sign change in gravitactic orientation is not mediated by the photoreceptor, since it was also found in cells that lack the pfb (colorless and blind mutants), but it is brought about by reactive oxygen species (ROS, probably hydrogen peroxide), as revealed by using the fluorescent probe 2′,7′-dichlorodihydrofluorescein diacetate. Flushing the cells with nitrogen, which removes the oxygen, or the application of Trolox, potassium cyanide, or ascorbic acid, which scavenge ROS, suppressed sign change of gravitational orientation
[44][48]. Extended exposure to solar radiation decreases the precision of gravitaxis
[42][45][46,49]. Even though
E. gracilis is adapted to freshwater, it tolerates salinity of up to 19 g/L. During this treatment, the swimming speed and precision of gravitaxis decreases. At salt concentrations above 15 g/L, negative gravitaxis changed to a positive one. However, it is interesting to note that the cells kept showing positive gravitaxis, even after transfer back into freshwater medium
[46][50].
The long axis of immobilized cells killed by liquid nitrogen pointed in random directions
[47][51]. These, and other, results indicate that gravioriention in
E. gracilis is based on an active perceptive mechanism. Some organisms use heavy bodies inside the cell with operate as statoliths
[48][49][52,53]. Since no obvious sedimenting bodies could be found in
E. gracilis by microscopic analysis, the alternative is that the whole cytoplasmic content of the cell with its organelles acts as a statolith
[50][51][54,55]. The specific weight of the cells was determined by isopygnic centrifugation in tubes with layers of increasing Ficoll concentrations
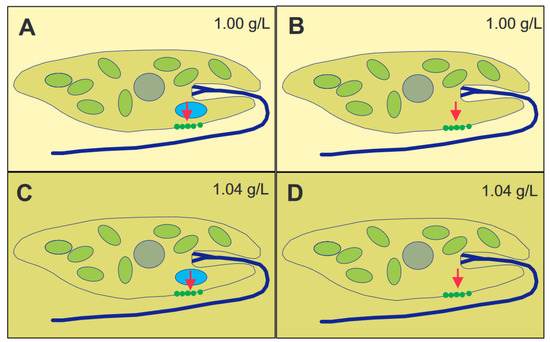
[52][56]. The specific density of the cells was found to be between 1.045 and 1.054 g/mL, depending on the culture age and conditions, as older cells were heavier than those in newly inoculated cultures. There is an elegant experiment to distinguish between the action of a heavy statolith within the cell or the whole cytoplasm (
Figure 2)
[53][54][57,58]. In case of a statolith, which presses onto a sensor inside the cell, it does not matter if the cell is in a 1.00 g/mL medium or Ficoll at 1.04 g/mL. In contrast, if the cell with a specific weight of 1.04 g/mL floats in a medium of the same density, it will not perceive the gravity pull, while it does so in 1.00 g/mL medium. Since
E. gracilis showed a response according to the latter scheme, it was clear that the whole cytoplasmic content pressed onto the lower membrane. Cells starved for over 600 days had a specific density of 1.011 g/mL and did not display any graviorientation
[55][59].
Figure 2. Principles of graviperception, either by means of a distinct gravisensor (here statolith) or the whole cell mass as sedimenting parameter. A statolith (blue) presses (red arrow) onto mechano-sensitive ion channels (A,C) or the whole cell content exerts pressure (red arrow) onto the channels (B,D). When the cells, having a specific weight of 1.04 g/L, are in a lower-density medium (1.00 g/L) (A,B), the pressure will open the channels, and the direction of gravi-stimulus will be detected. When the cells are under isodensity conditions, with respect to the surrounding medium ((C,D), 1.04 g/L), only the pressure of the statolith will be detected, but not that of the whole cell content.
Like many other organisms,
E. gracilis shows a dominant circadian rhythm, which is expressed in many physiological, biochemical, and behavioral processes
[56][57][60,61]. Under constant light and temperature conditions, individual cells are not synchronized, and no circadian rhythm was detected; however, when the cells were exposed to a circadian light/dark change, the precision of gravitaxis followed the rhythm with a minimum in the darkness and maximum in the early afternoon
[58][59][62,63]. The fact that the precision of orientation increased, even before the light was switched on, indicates that the internal circadian rhythm was entrained by the light/dark change
[60][64]. In addition, the form of the cells (elongated vs. rounded), swimming velocity, and internal concentration of cyclic adenosine monophosphate (cAMP) followed the circadian rhythm (see below for the role of cAMP in graviperception).
E. gracilis cells can also be synchronized to much shorter light/dark cycles, down to 1:1 h
[61][65].
During long space missions a considerable amount of food, oxygen and water has to be carried and the exhaled carbon dioxide has to be removed. In this context, E. gracilis is an excellent candidate for biological life support systems, since it produces oxygen by photosynthesis, takes up carbon dioxide and is even edible. Various species and mutants of Euglena are utilized as a producer of commercial food items as well as a source of medicines, as it produces a number of vitamins, contains numerous trace elements, and synthesizes dietary proteins, lipids and the reserve molecule paramylon. Euglena has anti-inflammatory, anti-oxidant and anti-obesity properties.