ThCance endocannabinoid system (ECS) is an ancient homeostasis mechanism operating from embryonic stages to adulthood. It controls the growth and development of many cells and cell lineages. Dysregulation of the cr is a disease which affects approximately 40% of people in their lifetime. Chemotherapy, the primary choice for treatment of cancer, is often ineffective or/and presents itself with many debilitating side effects, including loss of appetite, nausea, insomnia, and anxiety. Components of the ECS may result in uncannabis extracts, including cannabinoids and terpenes, may present an alternative for controlled proliferation, adhesion, invasion, inhibition of apoptosis and increased vascularization, leading to theing side effects and may be used for tumor shrinkage together with chemodrugs. Cannabinoids act on so called endocannabinoid system (ECS) that operates in human body to maintain homeostasis. ECS promotes healthy development of various malignancies. Cancer is the disease of uncontrolled cell divisiontissues and regulates many processes in our organism and when disbalanced may lead to disease, including cancer.
- endocannabinoid system
- cancer and carcinogenesis
- primary care
- palliative care
- cannabinol
- tetrahydrocannabinol
1. Introduction
2. Role of Endocannabinoids in the Human Body
ECS is active in virtually all cells of ohurman organism. It plays an important role in the reproduction, function, and proper development of gametes [7], fertilization event, embryo implantation, and proper placenta development [8]. It is also active at all stages of embryogenesis, regulating cell division, and tissue and organ development, specifically, regulating differentiation of neural progenitors, synaptogenesis, and axonal migration [9]. During human adult life, it regulates homeostasis of many tissues, playing critical role in proper brain function by regulating neuronal synaptic communications affecting critical organismal functions, including general metabolism, growth and development, reproduction, learning and memory formation, mood, and behavior, among others [10]. In the peripheral tissues, endocannabinoids are involved in endocrine regulation and energy balance [11], as well as regulating the function of innate and adaptive immune system and immune response [12], regulating cell migration and apoptosis. The activity and functionality of ECS depends on many factors, from cell- and tissue-specific differences in the synthesis of endocannabinoids, to the number and the activity of endocannabinoid and auxiliary receptors, to the expression and the activity of enzymes involved in the degradation of circulating endocannabinoids. In the cells, endocannabinoids acting in CB-receptor-dependent and independent manner exhibit anti-oxidative properties, are involved in clearance of damaged molecules and regulate mitochondrial activity. Anti-oxidative properties are associated with the inhibition of production of reactive oxygen species (ROS), metal chelation and prevention/alleviation of ROS-induced cell damage [13]. It should be noted that the anti-oxidative effects of cannabinoids are cell specific—while in most cells of the body, they mitigate oxidative stress, in hepatic cells they may cause it, leading to cell death [14]. Similarly, in cancer cells, such as gliomas and leukemia, cannabinoids promote oxidative stress [13]. Cannabinoids contribute to recycling of damaged molecules and are likely involved in autophagy in health tissues [15]—the activity well documented in cancer cells (discussed below). In normal cells, they increase lysosomal stability and integrity [15] through CB1 receptors found on the surface of lysosomes. CB1 receptors are also present on the surface of mitochondria. They regulate mitochondrial oxidative phosphorylation in a positive and a negative manner, acting through the CB1 receptor, but it is not clear what modulates this activity [13]. When cells are stressed, cannabinoids attenuate mitochondrial damage [16] and decrease calcium-induced cytochrome c release [17].2.1. Mechanism of Action—Ligand/Receptor
Cannabinoid receptors are ubiquitous and expressed on the cell surface as well on cell organelles, including mitochondria and lysosomes. Classical cannabinoid receptors include CB1 and CB2. CB1 is expressed at a higher level in central and peripheral nervous systems, while CB2 is expressed in many different tissues, including the immune system, internal organs, skin, bone, muscle, and glia in the brain [18]. CB1 and CB2 are GPCR (Gi/o) protein-coupled receptors, and when activated, they modulate various cellular functions through receptor internalization; interaction with other G-protein-coupled receptors; inhibition of adenylyl cyclase activity, changing the activity of calcium and potassium channels; increasing phosphorylation of various mitogen-activated protein kinases (MAPK); and many more functions [12] (Figure 1).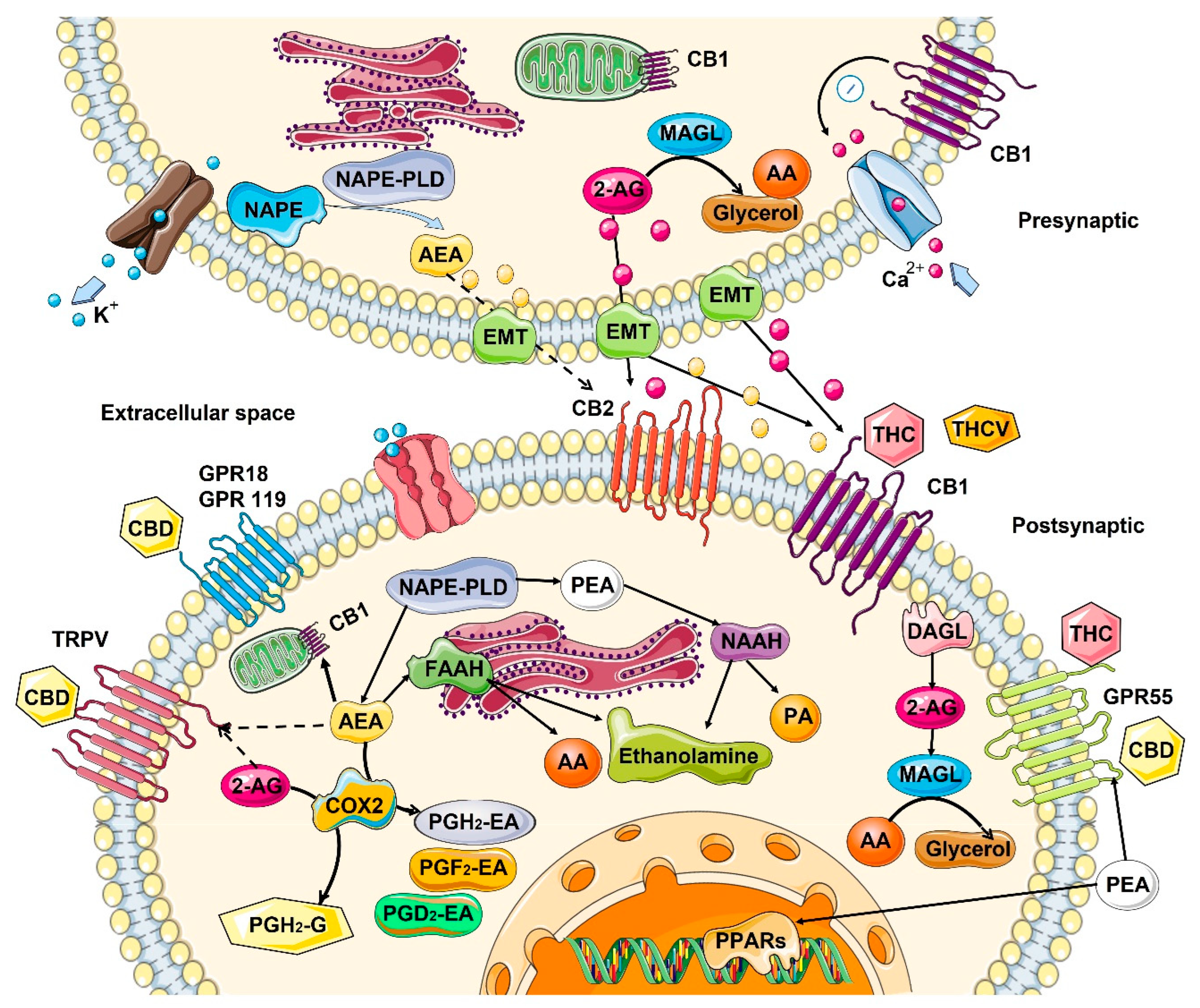
2.2. Role in the Control of Cell Division and Cell Proliferation
It appears that ECS controls the fate of many cells in the organism, regulating the cell division and proliferation, apoptosis, necrosis and autophagy in several organs and organ systems, including the brain, skin, and immune system. In the central nervous system (CNS), the ECS system functions as a neuroprotective system that controls glutamate excitotoxicity, calcium influx, inflammation, and autophagy [25]. In the CNS, the interaction of endocannabinoids with CB1/CB2 and other receptors mediates synaptic plasticity or progenitor cell fate in the central nervous system, promoting self-repair of the brain [26]. It also appears that constitutive release of 2-arachidonoylglycero by late oligodendrocyte progenitors allows oligodendrocyte maturation by activating CB receptors and downstream ERK pathway [27]. In skin, ECS activity maintains the cutaneous homeostasis through the regulation of skin cell proliferation, survival, and differentiation [28]. Locally produced AEA inhibits the cellular growth and the differentiation of cultured NHEK and HaCaT keratinocytes, as well as inducing apoptosis of human HaCaT keratinocytes [28]. CB1 activity is higher in differentiated skin layers [29]. In human cultured hair follicles, AEA but not 2-AG inhibit elongation and proliferation of hair shaft and induce intraepithelial apoptosis in a CB1-dependent manner [30]. Both AEA and 2-AG induce apoptosis of human sebaceous-gland-derived SZ95 sebocytes in a CB2-dependent manner [31]. In the immune system, the central role is played by CB2 receptors that are mainly expressed by cells (T and B lymphocytes) and peripheral tissues of the immune system (spleen and thymus) where it regulates immune suppression, apoptosis, and cell migration [32]. In in vitro studies, it was demonstrated that anandamide inhibits mitogen-induced proliferation of T cells [33], while inhibiting the chemokine SDF-1-induced migration of CD8+ T cells [34]. In contrast, 2-AG, but not anandamide, induced CB2-dependent migration in natural killer cell line KHYG-1 cells [35]. In B cells, 2-AG chemo-attracts naïve B cells and marginal zone B cells and inhibits the function of activated B cells, while 2-AG and anandamide suppress the migration of neutrophils [36]. Additionally, anandamide induces the apoptosis of murine bone-marrow-derived DCs (BMDCs) in a CB1- and CB2-dependent manner [37].2.3. Changes in the ECS with Age
Cancer can be considered an age-associated disease, due to the accumulation of cellular and DNA damage. From this perspective, it is interesting to understand what happens to ECS with age. In general, information about age-related changes in the ECS is scarce. Most of the data are related to changes in the central nervous system, and even then, the data are very contradictory. In general, it is believed that the activity of ECS declines with age [13]. In rats, in one study, a general decrease in the expression of CB1 and a decrease in density of the receptors in various brain areas with age was observed [38], while in another study—in which only redistribution of the receptors was noted– they were reduced in the postrhinal, but elevated in the entorhinal and temporal cortices in old animals [39] (Table 1). In mice, no changes in the receptor density in most brain regions was found with age, but instead, a significantly reduced receptor/Gi protein coupling was observed [40]. In one study on humans, CB1 expression increased, predominantly in females, most drastically in the basal ganglia, the lateral temporal cortex, and in the hippocampus [41], while another study reported no change [39]. As for endocannabinoids, the picture is not clear either—some studies suggested a decrease, while others found no difference in different brain regions of young and old animals. [13]. However, animals lacking FAAH—the enzyme degrading anandamide showed less pronounced features of aging—decreased expression of pro-inflammatory genes and decreased decline in cardiac function [42] (Table 1).| Tissues/Organs | Endocannabinoids | Receptors | Metabolizing Enzymes |
|---|---|---|---|
| Skin | No reliable data | ↓ in CB1 expression [13] | FAAH tends to ↓ with age [45][43] |
| Lung | 2-AG ↓ and AEA ↑ in mice [46][44] | No reliable data | No reliable data |
| Brain | From no change [40] to a ↓ in AEA [47][45] ↓ in 2-AG levels in mice [48][46] |
From ↑ in humans [41] to no change [39] to a ↓ [38,49][38][47] in mice/rats in CB1 expression, brain area-specific | ↓ FAAH activity in rats [50][48] ↑ in MAGL levels in mice [48][46] |
| Blood | Small ↑ in 2-AG and AEA in mice [46][44] | No reliable data | No reliable data |
3. Effect of Cannabinoids on Various Hallmarks of Cancer
Various in vitro and in vivo experiments have shown that cannabinoids can target almost every hallmark of cancer (Figure 2) [186][51]. They inhibit proliferation, reduce inflammation, stimulate apoptosis, and inhibit tumor invasiveness, angiogenesis, and metastasis [187,188,189,190][52][53][54][55]. One of the most important effects of cannabinoids, besides their antitumor ability, is that they are less likely to affect non-transformed normal cells surrounding tumors, and they may even have protective effects. For instance, cannabinoids may induce cell death in glioma cells while protecting normal astroglial and oligodendroglial cells from apoptosis via CB1 receptors [187][52]. Studies on animals show the protective effects of cannabinoids against certain types of tumors. For example, a dose-dependent decrease in the incidence of hepatic adenomas and hepatocellular carcinomas in mice that were given THC over 2 years was noted. Additionally, lower incidence rates of benign tumors in mammary glands, uterus, testis, and pancreas were seen in tested rats [191][56].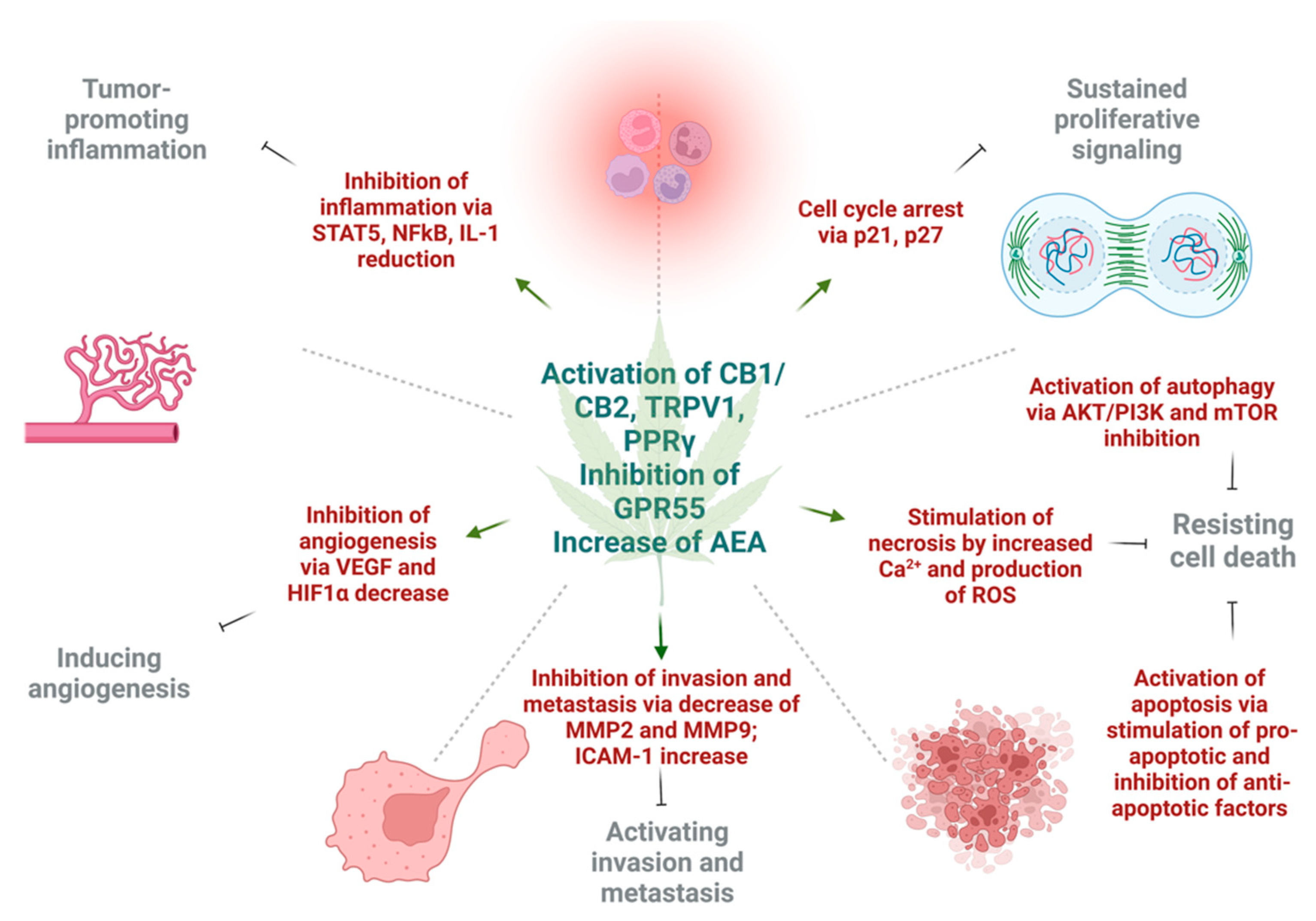
3.1. Induction of Autophagy and Apoptosis
Autophagy and apoptosis are two essential mechanisms of regulation of uncontrolled growth. Autophagic activity of cannabinoids observed in several major cancers [192,193][57][58] is partially dependent on the CB1 or CB2 receptor. Mice deficient in CB1 receptor exhibit altered autophagosomal activity [13], while endocannabinoid palmitoylethanolamide increased the phagocytosis of murine microglial cells [194][59]. Additionally, the experimental study using delta-9-THC and a synthetic agonist decreased the cell viability of hepatocellular carcinoma xenografts in nude mice via the CB2 receptors. The anti-cancer effect was explained by activating the endoplasmic reticulum stress response, which leads to macro-autophagy and eventually apoptosis [195][60]. Studies on small-cell lung carcinoma [61] and breast cancer cells [67][62] supported the idea that CB1 and CB2 receptors may be potential targets to achieve apoptosis. The preclinical models of breast cancer showed evidence that CBD may induce apoptosis in estrogen-dependent and estrogen-independent breast cancer cells with little or no effect on normal mammary cells. Surprisingly, this was CB1-, CB2-, and vanilloid receptor-independent [176][63]. The well-established antineoplastic mechanisms of cannabinoids are alterations in ceramide de novo synthesis. In cancer cells, increased ceramide levels, a neutral lipid backbone of complex sphingolipids, can occur under chemotherapy, radiation, and stimulation of CB receptors [58,196][64][65]. As a result, ceramide activates endoplasmic reticulum stress response and causes inhibition of global translation of proteins. At the same time, there is an activation of C/EBP homology protein (CHOP) which can stimulate proapoptotic proteins BAD and BAX [197][66]. Moreover, cannabinoids can cause downregulation of AKT, which may have a variety of intracellular effects. Low AKT leads to activation of autophagy via the mTOR pathway, cell cycle arrest through p21, and activation of caspase 9 and 3, which eventually ends in apoptotic cell death [58,108,155,198,199,200][64][67][68][69][70][71]. Activation of CB1 and CB2 receptors by synthetic cannabinoid agonists could stimulate apoptosis via ceramide synthesis and TNF-receptor activation [58][64]. Another group showed that activation of CB1 receptors in different CRC cell lines causes inhibition of major cancer survival pathways such as RAS/MAPK, ERK1, and PI3K/AKT [155][68]. Additionally, CBD, a partial agonist of CB1/CB2 receptors and antagonist of GPR55, may suppress mTOR/AKT signaling and activate proapoptotic NOXA in CRC cells [201][72]. Moreover, CBD suppressed the production of inhibitors of apoptosis, such as survivin and c-FLIP in colon cancer cells [197][66].3.2. Reduction of Inflammation and Inhibition of Proliferation
Inflammation is a large component of carcinogenesis. ECS plays a central in the regulation of function of immune system and control of inflammation. Similarly, many phytocannabinoids exert strong anti-inflammatory effects upon local [202][73] or systemic [203][74] application. Cannabinoids inhibited proliferation by suppressing the AKT/PKB prosurvival pathway causing cell cycle arrest in G1/S phase. This was shown in multiple cancers, including melanoma, breast, gastric, lung, and liver carcinomas [93,151,159,160,204,205][75][76][77][78][79][80]. In a breast cancer model, cannabinoids were able to induce cell cycle arrest via inhibition of cyclin dependent kinase 1 (CDK1), induction of p21 and p27, a decrease in cyclin A and E levels, degradation of CDC25A, and finally, inactivation of CDK2 [206,207][81][82]. In the study on head and neck squamous cell carcinoma, cannabinoids were able to stimulate dual specificity phosphatase 1 (DUSP1), which is a negative regulator of MAPK [208][83]. DUSP1 is one of the central mediators in the resolution of inflammation in cells. Moreover, the levels of cyclin dependent kinase inhibitor, p21, as well as growth arrest and DNA damage-inducible protein α (GADD45A) were activated, resulting in cannabinoid’s antiproliferative effects. In human gastric cancer model, CBD upregulated ATM and p21, which caused a decrease in CDK2 and CCNE, resulting in cell arrest in G0/G1 stage [209][84]. In a xenograft model of human glioma, CBD was able to reduce the activity of 5-lipoxygenase, an enzyme that catalyzes synthesis of leukotrienes (LTs) and mediators of inflammation; a decrease in 5-lipoxygenase activity caused inhibition of LTB4 and had antiproliferative effect [210][85]. The eicosanoid system, which contains pro- and anti-inflammatory molecules, plays an important role in cannabinoid-induced tumor cell apoptosis. The addition of R(+)methanandamide to the glioma cells activated de novo ceramide synthesis, which eventually led to COX-2 expression with subsequent production of PGE2 that had proapoptotic effect [211,212][86][87]. It was shown that proapoptotic effects of eicosanoids was PPARγ receptor-dependent [213,214,215][88][89][90]. On the other hand, it was shown that the micromolar concentrations of THC, CB1 agonist—arachidonyl-2-chloroethylamine (ACEA), and CB2 agonist HU308 stimulated the proliferation of cancer cells, which can be explained by transactivation of EGFR [169,171,216,217][91][92][93][94]. The chemoprotective effect of CBD was also shown on colorectal cancer in mice. Adding CBD prevented premalignant and malignant lesions development in the azoxymethane model of colon cancer [218][95]. The effect was explained by DNA protection against oxidative damage, increased levels of endocannabinoids, and decreased cell proliferation [218][95]. The antiproliferative action was CB1 dependent [219][96].3.3. Inhibition of Angiogenesis, Tumor Invasiveness, and Metastasis
There were multiple reports showing the inhibitory effects of cannabinoids on cancer cell migration, invasion, and metastasis [62,74,220][97][98][99]. CBD was shown to inhibit the invasiveness of lung cancer cell lines by inhibiting ICAM-1 [190][55]. As some experiments indicated, the induction of tissue inhibitor of metalloproteinase-1 (TIMP-1), and ICAM-1 by THC, Met-AEA and CBD had significant anti-invasive effects [105,190,221][55][100][101]. The action of TIMP-1 is achieved via reduction of collagen-degrading enzymes, MMP-2 and MMP-9, that promote cancer cell invasiveness [222][102]. Another way in which cannabinoids are diminishing tumor aggressiveness is inhibition of epithelial-to-mesenchymal transition. A study that involved 2-methyl-2′-F-anandamide (Met-F-AEA) showed a significant reduction in β-catenin, vimentin, N-cadherin, and fibronectin, which are considered mesenchymal markers in tumor invasion. Moreover, Met-F-AEA decreased the levels of EMT markers such as Snail1, Slug, and Twist [223][103]. Other studies showed that CBD may reverse an IL-1β-induced EMT in breast cancer cells [224][104], or TGF-β-induced reorganization of F-actin, which also corresponds to EMT in lung cancer cells [60][105]. Cannabinoids may inhibit the invasion and metastasis of cancer cells through downregulation of vascular endothelial growth factor (VEGF), matrix metalloproteinase 2, matrix metalloproteinase 9, E-cadherin, cyclooxygenase 2 (COX-2), and hypoxia-inducible factor α [225,226,227][106][107][108].References
- McPartland, J.M.; Matias, I.; Di Marzo, V.; Glass, M. Evolutionary origins of the endocannabinoid system. Gene 2006, 370, 64–74.
- Cristino, L.; Becker, T.; Di Marzo, V. Endocannabinoids and energy homeostasis: An update. BioFactors 2014, 40, 389–397.
- Rodríguez-Valentín, R.; Torres-Mejía, G.; Martínez-Matsushita, L.; Angeles-Llerenas, A.; Gómez-Flores-Ramos, L.; Wolff, R.K.; Baumgartner, K.B.; Hines, L.M.; Ziv, E.; Flores-Luna, L.; et al. Energy homeostasis genes modify the association between serum concentrations of IGF-1 and IGFBP-3 and breast cancer risk. Sci. Rep. 2022, 12, 1837.
- Khandekar, M.J.; Cohen, P.; Spiegelman, B.M. Molecular mechanisms of cancer development in obesity. Nat. Cancer 2011, 11, 886–895.
- Kunos, G.; Osei-Hyiaman, D.; Liu, J.; Godlewski, G.; Bátkai, S. Endocannabinoids and the control of energy homeostasis. J. Biol. Chem. 2008, 283, 33021–33025.
- Loeb, L.A.; Loeb, K.R.; Anderson, J.P. Multiple mutations and cancer. Proc. Natl. Acad. Sci. USA 2003, 100, 776–781.
- Battista, N.; Bari, M.; Maccarrone, M. Endocannabinoids and Reproductive Events in Health and Disease. Endocannabinoids 2015, 231, 341–365.
- Kozakiewicz, M.L.; Grotegut, C.A.; Howlett, A.C. Endocannabinoid System in Pregnancy Maintenance and Labor: A Mini-Review. Front. Endocrinol. 2021, 12, 699951.
- Nghdawagsb, E.F. The endocannabinoid system during development: Emphasis on perinatal events and delayed effects. In Vitamins and Hormones; Elsevier: Amsterdam, The Netherlands, 2009; pp. 139–158.
- Skaper, S.D.; di Marzo, V. Endocannabinoids in nervous system health and disease: The big picture in a nutshell. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3193–3200.
- Pagotto, U.; Marsicano, G.; Cota, D.; Lutz, B.; Pasquali, R. The Emerging Role of the Endocannabinoid System in Endocrine Regulation and Energy Balance. Endocr. Rev. 2005, 27, 73–100.
- Almogi-Hazan, O.; Or, R. Cannabis, the Endocannabinoid System and Immunity—The Journey from the Bedside to the Bench and Back. Int. J. Mol. Sci. 2020, 21, 4448.
- Bilkei-Gorzo, A. The endocannabinoid system in normal and pathological brain ageing. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3326–3341.
- Michela, G.; Giuseppe, C.; Ornella, P.; Patrizia, P.; Daniela, C.; Renza, V.; Romano, L.L. Anandamide-induced apoptosis in Chang liver cells involves ceramide and JNK/AP-1 pathway. Int. J. Mol. Med. 2006, 17, 811–819.
- Noonan, J.; Tanveer, R.; Klompas, A.; Gowran, A.; McKiernan, J.; Campbell, V.A. Endocannabinoids Prevent β-Amyloid-mediated Lysosomal Destabilization in Cultured Neurons. J. Biol. Chem. 2010, 285, 38543–38554.
- Velez-Pardo, C.; Jimenez-Del-Rio, M.; Lores-Arnaiz, S.; Bustamante, J. Protective Effects of the Synthetic Cannabinoids CP55,940 and JWH-015 on Rat Brain Mitochondria upon Paraquat Exposure. Neurochem. Res. 2010, 35, 1323–1332.
- Zaccagnino, P.; D’Oria, S.; Romano, L.L.; Di Venere, A.; Sardanelli, A.M.; Lorusso, M. The endocannabinoid 2-arachidonoylglicerol decreases calcium induced cytochrome c release from liver mitochondria. J. Bioenerg. Biomembr. 2012, 44, 273–280.
- Mackie, K.; Mackie, K. Cannabinoid Receptors: Where They are and What They do. J. Neuroendocr. 2008, 20, 10–14.
- Tóth, K.F.; Ádám, D.; Bíró, T.; Oláh, A. Cannabinoid signaling in the skin: Therapeutic potential of the “c(ut)annabinoid” system. Molecules 2019, 24, 918.
- Baron, E.P. Medicinal Properties of Cannabinoids, Terpenes, and Flavonoids in Cannabis, and Benefits in Migraine, Headache, and Pain: An Update on Current Evidence and Cannabis Science. Headache J. Head Face Pain 2018, 58, 1139–1186.
- del, R.C.; Millán, E.; García, V.; Appendino, G.; DeMesa, J.; Muñoz, E. The endocannabinoid system of the skin. A potential approach for the treatment of skin disorders. Biochem. Pharmacol. 2018, 157, 122–133.
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.M.; Denovan-Wright, E.M. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. J. Cereb. Blood Flow Metab. 2015, 172, 4790–4805.
- Burstein, S. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorganic Med. Chem. 2015, 23, 1377–1385.
- Martínez, V.; Iriondo De-Hond, A.; Borrelli, F.; Capasso, R.; del Castillo, M.D.; Abalo, R. Cannabidiol and other non-psychoactive cannabinoids for prevention and treatment of gastrointestinal disorders: Useful nutraceuticals? Int. J. Mol. Sci. 2020, 9, 3067.
- Fernández-Ruiz, J.; Romero, J.; Ramos, J.A. Endocannabinoids and neurodegenerative disorders: Parkinson’s disease, Huntington’s chorea, Alzheimer’s disease, and others. In Endocannabinoids; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 233–259.
- Elphick, M.R. The evolution and comparative neurobiology of endocannabinoid signalling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3201–3215.
- Gomez, O.; Arevalo-Martin, A.; Garcia-Ovejero, D.; Ortega-Gutierrez, S.; Cisneros, J.A.; Almazan, G.; Sánchez-Rodriguez, M.A.; Molina-Holgado, F.; Molina-Holgado, E. The constitutive production of the endocannabinoid 2-arachidonoylglycerol participates in oligodendrocyte differentiation. Glia 2010, 58, 1913–1927.
- Bíró, T.; Tóth, B.I.; Haskó, G.; Paus, R.; Pacher, P. The endocannabinoid system of the skin in health and disease: Novel perspectives and therapeutic opportunities. Trends Pharmacol. Sci. 2009, 30, 411–420.
- Ständer, S.; Schmelz, M.; Metze, D.; Luger, T.; Rukwied, R. Distribution of cannabinoid receptor 1 (CB1) and 2 (CB2) on sensory nerve fibers and adnexal structures in human skin. J. Dermatol. Sci. 2005, 38, 177–188.
- Telek, A.; Bíró, T.; Bodó, E.; Tóth, B.I.; Borbíró, I.; Kunos, G.; Sardanelli, A.M. Inhibition of human hair follicle growth by endo-and exocannabinoids. FASEB J. 2007, 21, 3534–3541.
- Dobrosi, N.; Tóth, B.I.; Nagy, G.; Dózsa, A.; Géczy, T.; Nagy, L.; Harvey-White, J.; Loureiro, A.I. Endocannabinoids enhance lipid synthesis and apoptosis of human sebocytes via cannabinoid receptor-2-mediated signaling. FASEB J. 2008, 22, 3685–3695.
- Ibsen, M.S.; Connor, M.; Glass, M. Cannabinoid CB1 and CB2 Receptor Signaling and Bias. Cannabis Cannabinoid Res. NLM (Medlin.) 2017, 2, 48–60.
- Schwarz, H.; Blanco, F.J.; Lotz, M. Anadamide, an endogenous cannabinoid receptor agonist inhibits lymphocyte proliferation and induces apoptosis. J. Neuroimmunol. 1994, 55, 107–115.
- Joseph, J.; Niggemann, B.; Zaenker, K.S.; Entschladen, F. Anandamide is an endogenous inhibitor for the migration of tumor cells and T lymphocytes. Cancer Immunol. Immunother. 2004, 53, 723–728.
- Kishimoto, S.; Muramatsu, M.; Gokoh, M.; Oka, S.; Waku, K.; Sugiura, T. Endogenous Cannabinoid Receptor Ligand Induces the Migration of Human Natural Killer Cells. J. Biochem. 2005, 137, 217–223.
- Rahaman, O.; Ganguly, D. Endocannabinoids in immune regulation and immunopathologies. Immunology 2021, 164, 242–252.
- Do, Y.; McKallip, R.J.; Nagarkatti, M.; Nagarkatti, P.S. Activation through Cannabinoid Receptors 1 and 2 on Dendritic Cells Triggers NF-κB-Dependent Apoptosis: Novel Role for Endogenous and Exogenous Cannabinoids in Immunoregulation. J. Immunology. 2004, 173, 2373–2382.
- Berrendero, F.; Romero, J.; García-Gil, L.; Suarez, I.; De la Cruz, P.; Ramos, J.; Fernández-Ruiz, J. Changes in cannabinoid receptor binding and mRNA levels in several brain regions of aged rats. Biochim. et Biophys. Acta (BBA)-Mol. Basis Dis. 1998, 1407, 205–214.
- Liu, P.; Bilkey, D.K.; Darlington, C.L.; Smith, P.F. Cannabinoid CB1 receptor protein expression in the rat hippocampus and entorhinal, perirhinal, postrhinal and temporal cortices: Regional variations and age-related changes. Brain Res. 2003, 979, 235–239.
- Wang, L.; Liu, J.; Harvey-White, J.; Zimmer, A.; Kunos, G. Endocannabinoid signaling via cannabinoid receptor 1 is involved in ethanol preference and its age-dependent decline in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 1393–1398.
- Van Laere, K.; Goffin, K.; Casteels, C.; Dupont, P.; Mortelmans, L.; de Hoon, J.; Bormans, G. Gender-dependent increases with healthy aging of the human cerebral cannabinoid-type 1 receptor binding using MK-9470 PET. Neuroimage 2008, 39, 1533–1541.
- Bátkai, S.; Rajesh, M.; Mukhopadhyay, P.; Haskó, G.; Liaudet, L.; Cravatt, B.F.; Csiszár, A.; Ungvári, Z.; Pacher, P. Decreased age-related cardiac dysfunction, myocardial nitrative stress, inflammatory gene expression, and apoptosis in mice lacking fatty acid amide hydrolase-1) expression and monocyte-endothelial adhesion in Downloaded from. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, 909–918.
- Robben, M.; Nasr, M.S.; Das, A.; Huber, M.; Jaworski, J.; Weidanz, J.; Luber, J.M. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 28, D20–D26.
- Pati, S.; Krishna, S.; Lee, J.H.; Ross, M.K.; de La Serre, C.B.; Harn, D.A.; Wagner, J.; Filipov, N.M.; Cummings, B.S. Effects of high-fat diet and age on the blood lipidome and circulating endocannabinoids of female C57BL/6 mice. Biochim. et Biophys. Acta BBA-Mol. Cell Biol. Lipids 2017, 1863, 26–39.
- Maccarrone, M.; Valverde, O.; Barbaccia, M.L.; Castañé, A.; Maldonado, R.; Ledent, C.; Parmentier, M.; Finazzi-Agrò, A. Age-related changes of anandamide metabolism in CB1cannabinoid receptor knockout mice: Correlation with behaviour. Eur. J. Neurosci. 2002, 15, 1178–1186.
- Piyanova, A.; Lomazzo, E.; Bindila, L.; Lerner, R.; Albayram, O.; Ruhl, T.; Lutz, B.; Zimmer, A.; Bilkei-Gorzo, A. Age-related changes in the endocannabinoid system in the mouse hippocampus. Mech. Ageing Dev. 2015, 150, 55–64.
- Romero, J.; Berrendero, F.; Garcia-Gil, L.; de la Cruz, P.; Ramos, J.; Fernandez-Ruiz, J. Loss of cannabinoid receptor binding and messenger RNA levels and cannabinoid agonist-stimulated guanylyl-5′-O-(thio)-triphosphate binding in the basal ganglia of aged rats. Neuroscience 1998, 84, 1075–1083.
- Lee, T.T.-Y.; Hill, M.N.; Hillard, C.J.; Gorzalka, B.B. Temporal changes inN-acylethanolamine content and metabolism throughout the peri-adolescent period. Synapse 2012, 67, 4–10.
- Correia-Sá, I.B.; Carvalho, C.M.; Serrão, P.V.; Loureiro, A.I.; Fernandes-Lopes, C.; Marques, M.; Vieira-Coelho, M.A. A new role for anandamide: Defective link between the systemic and skin endocannabinoid systems in hypertrophic human wound healing. Sci. Rep. 2020, 10, 3.
- Leal, E.C.; Moura, L.I.; Pirzgalska, R.M.; Marques-da-Silva, D.; Ledent, C.; Köfalvi, A.; Carvalho, E. Diabetes and Cannabinoid CB1 receptor deficiency promotes similar early onset aging-like changes in the skin. Exp. Gerontol. 2021, 15, 154.
- Pyszniak, M.; Tabarkiewicz, J.; Łuszczki, J.J. Endocannabinoid system as a regulator of tumor cell malignancy–biological pathways and clinical significance. OncoTargets Ther. 2016, 9, 4323–4336.
- Guzmán, M. Cannabinoids: Potential anticancer agents. Nature Reviews Cancer. Eur. Assoc. Cardio-Thorac. Surg. 2003, 3, 745–755.
- Blázquez, C.; Casanova, M.L.; Planas, A.; del Pulgar, T.G.; Villanueva, C.; Fernández-Aceñero, M.J.; Aragones, J.; Huffman, J.W.; Jorcano, J.L.; Guzman, M. Inhibition of tumor angiogenesis by cannabinoids. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biology. 2003, 17, 529–531.
- Vaccani, A.; Massi, P.; Colombo, A.; Rubino, T.; Parolaro, D. Cannabidiol inhibits human glioma cell migration through a cannabinoid receptor-independent mechanism. J. Cereb. Blood Flow Metab. 2005, 144, 1032–1036.
- Ramer, R.; Bublitz, K.; Freimuth, N.; Merkord, J.; Rohde, H.; Haustein, M.; Borchert, P.; Schmuhl, E.; Linnebacher, M.; Hinz, B. Cannabidiol inhibits lung cancer cell invasion and metastasis via intercellular adhesion molecule-1. FASEB J. 2011, 26, 1535–1548.
- National Toxicology Program: NTP toxicology and carcinogenesis studies of 1-trans-delta(9)-tetrahydrocannabinol (CAS No. 1972-08-3) in F344 rats and B6C3F1 mice (gavage studies). NTP Tech. Rep. 1996, 446, 1–317.
- Donadelli, M.; Dando, I.; Zaniboni, T.; Costanzo, C.; Pozza, E.D.; Scupoli, M.; Scarpa, A.; Zappavigna, S.; Marra, M.; Abbruzzese, A.; et al. Gemcitabine/cannabinoid combination triggers autophagy in pancreatic cancer cells through a ROS-mediated mechanism. Cell Death Dis. 2011, 2, e152.
- Salazar, M.; Carracedo, A.; Salanueva, J.; Hernández-Tiedra, S.; Lorente, M.; Egia, A.; Vázquez, P.; Blázquez, C.; Torres, S.; García, S.; et al. Cannabinoid action induces autophagy-mediated cell death through stimulation of ER stress in human glioma cells. J. Clin. Investig. 2009, 119, 1359–1372.
- Redlich, S.; Ribes, S.; Schütze, S.; Czesnik, D.; Nau, R. Palmitoylethanolamide stimulates phagocytosis of Escherichia coli K1 and Streptococcus pneumoniae R6 by microglial cells. J. Neuroimmunol. 2012, 244, 32–34.
- Vara, D.; Salazar, M.; Olea-Herrero, N.; Guzmán, M.; Velasco, G.; Díaz-Laviada, I. Anti-tumoral action of cannabinoids on hepatocellular carcinoma: Role of AMPK-dependent activation of autophagy. Cell Death Differ. 2011, 18, 1099–1111.
- Preet, A.; Qamri, Z.; Nasser, M.W.; Prasad, A.; Shilo, K.; Zou, X.; Groopman, J.E.; Ganju, R.K. Cannabinoid receptors, CB1 and CB2, as novel targets for inhibition of non-small cell lung cancer growth and metastasis. Cancer Prev. Res. 2011, 4, 65–75.
- Nasser, M.W.; Qamri, Z.; Deol, Y.S.; Smith, D.; Shilo, K.; Zou, X.; Ganju, R.K. Crosstalk between Chemokine Receptor CXCR4 and Cannabinoid Receptor CB2 in Modulating Breast Cancer Growth and Invasion. PLoS ONE 2011, 6, e23901.
- Shrivastava, A.; Kuzontkoski, P.M.; Groopman, J.E.; Prasad, A. Cannabidiol Induces Programmed Cell Death in Breast Cancer Cells by Coordinating the Cross-talk between Apoptosis and Autophagy. Mol. Cancer Ther. 2011, 10, 1161–1172.
- Cianchi, F.; Papucci, L.; Schiavone, N.; Lulli, M.; Magnelli, L.; Vinci, M.C.; Messerini, L.; Manera, C.; Ronconi, E.; Romagnani, P.; et al. Cannabinoid Receptor Activation Induces Apoptosis through Tumor Necrosis Factor α–Mediated Ceramide De novo Synthesis in Colon Cancer Cells. Clin. Cancer Res. 2008, 14, 7691–7700.
- Sánchez, C.; Galve-Roperh, I.; Rueda, D.; Guzmán, M. Involvement of Sphingomyelin Hydrolysis and the Mitogen-Activated Protein Kinase Cascade in the Δ9-Tetrahydrocannabinol-Induced Stimulation of Glucose Metabolism in Primary Astrocytes. Mol. Pharmacol. 1998, 54, 834–843.
- Ogretmen, B.; Hannun, Y.A. Biologically active sphingolipids in cancer pathogenesis and treatment. Nat. Cancer 2004, 4, 604–616.
- Wang, D.; Wang, H.; Ning, W.; Backlund, M.G.; Dey, S.K.; DuBois, R.N. Loss of Cannabinoid Receptor 1 Accelerates Intestinal Tumor Growth. Cancer Res. 2008, 68, 6468–6476.
- Greenhough, A.; Patsos, H.A.; Williams, A.C.; Paraskeva, C. The cannabinoid Δ9-tetrahydrocannabinol inhibits RAS-MAPK and PI3K-AKT survival signalling and induces BAD-mediated apoptosis in colorectal cancer cells. Int. J. Cancer 2007, 121, 2172–2180.
- Pellerito, O.; Notaro, A.; Sabella, S.; De Blasio, A.; Vento, R.; Calvaruso, G.; Giuliano, M. WIN induces apoptotic cell death in human colon cancer cells through a block of autophagic flux dependent on PPARγ down-regulation. Apoptosis 2014, 19, 985.
- Pertwee, R.G.; Cascio, M.G. Known Pharmacological Actions of Delta-9-Tetrahydrocannabinol and of Four Other Chemical Constituents of Cannabis that Activate Cannabinoid Receptors. In Handbook of Cannabis; Oxford University Press: Oxford, UK, 2014; Volume 115–136, p. 6.
- Velasco, G.; Sánchez, C.; Guzmán, M. Towards the use of cannabinoids as antitumour agents. Nat. Rev. Cancer 2012, 12, 436–444.
- Jeong, S.; Yun, H.K.; Jeong, Y.A.; Jo, M.J.; Kang, S.H.; Kim, J.L.; Kim, D.Y.; Park, S.H.; Kim, B.R.; Na, Y.J.; et al. Cannabidiol-induced apoptosis is mediated by activation of Noxa in human colorectal cancer cells. Cancer Lett. 2019, 447, 12–23.
- Tubaro, A.; Giangaspero, A.; Sosa, S.; Negri, R.; Grassi, G.; Casano, S.; Della Loggia, R.; Appendino, G.B. Comparative topical anti-inflammatory activity of cannabinoids and cannabivarins. Fitoterapia 2010, 81, 816–819.
- Anil, S.M.; Peeri, H.; Koltai, H. Medical Cannabis Activity Against Inflammation: Active Compounds and Modes of Action. Front. Pharmacol. 2022, 13, 908198.
- Blázquez, C.; Carracedo, A.; Barrado, L.; Real, P.J.; Fernández-Luna, J.L.; Velasco, G.; Malumbres, M.; Guzmán, M. Cannabinoid receptors as novel targets for the treatment of melanoma. FASEB J. 2006, 20, 2633–2635.
- Caffarel, M.M.; Andradas, C.; Mira, E.; Pérez-Gómez, E.; Cerutti, C.; Moreno-Bueno, G.; Flores, J.M.; García-Real, I.; Palacios, J.; Mañes, S.; et al. Cannabinoids reduce ErbB2-driven breast cancer progression through Akt inhibition. Mol. Cancer 2010, 9, 196.
- Rao, M.; Chen, D.; Zhan, P.; Jiang, J. MDA19, a novel CB2 agonist, inhibits hepatocellular carcinoma partly through inactivation of AKT signaling pathway. Biol. Direct 2019, 14, 9.
- Boyacıoğlu, C.; Bilgiç, E.; Varan, C.; Bilensoy, E.; Nemutlu, E.; Sevim, D.; Kocaefe, L.; Korkusuz, P. ACPA decreases non-small cell lung cancer line growth through Akt/PI3K and JNK pathways in vitro. Cell Death Dis. 2021, 12, 1–14.
- Xian, X.-S.; Park, H.; Cho, Y.K.; Lee, I.S.; Kim, S.W.; Choi, M.-G.; Chung, I.-S.; Han, K.-H.; Park, J.M. Effect of a synthetic cannabinoid agonist on the proliferation and invasion of gastric cancer cells. J. Cell. Biochem. 2010, 110, 321–332.
- Hinz, B.; Ramer, R. Cannabinoids as anticancer drugs: Current status of preclinical Res. Br. J. Cancer 2022, 127, 1–13.
- Caffarel, M.M.; Sarrió, D.; Palacios, J.; Guzmán, M.; Sanchez, C. Δ9-Tetrahydrocannabinol Inhibits Cell Cycle Progression in Human Breast Cancer Cells through Cdc2 Regulation. Cancer Res. 2006, 66, 6615–6621.
- Laezza, C.; Pisanti, S.; Crescenzi, E.; Bifulco, M. Anandamide inhibits Cdk2 and activates Chk1 leading to cell cycle arrest in human breast cancer cells. FEBS Lett. 2006, 580, 6076–6082.
- Go, Y.Y.; Kim, S.R.; Kim, D.Y.; Chae, S.W.; Song, J.J. Cannabidiol enhances cytotoxicity of anti-cancer drugs in human head and neck squamous cell carcinoma. Sci. Rep. 2020, 1, 10.
- Zhang, X.; Qin, Y.; Pan, Z.; Li, M.; Liu, X.; Chen, X.; Qu, G.; Zhou, L.; Xu, M.; Zheng, Q.; et al. Cannabidiol Induces Cell Cycle Arrest and Cell Apoptosis in Human Gastric Cancer SGC-7901 Cells. Biomolecules 2019, 9, 302.
- Massi, P.; Valenti, M.; Vaccani, A.; Gasperi, V.; Perletti, G.; Marras, E.; Fezza, F.; Maccarrone, M.; Parolaro, D. 5-Lipoxygenase and anandamide hydrolase (FAAH) mediate the antitumor activity of cannabidiol, a non-psychoactive cannabinoid. J. Neurochem. 2008, 104, 1091–1100.
- Ramer, R.; Weinzierl, U.; Schwind, B.; Brune, K.; Hinz, B. Ceramide Is Involved in R()-Methanandamide-Induced Cyclooxygenase-2 Expression in Human Neuroglioma Cells. Mol. Pharmacol. 2003, 64, 1189–1198.
- Hinz, B.; Ramer, R.; Eichele, K.; Weinzierl, U.; Brune, K. Up-Regulation of Cyclooxygenase-2 Expression Is Involved in R(+)-Methanandamide-Induced Apoptotic Death of Human Neuroglioma Cells. Mol. Pharmacol. 2004, 66, 1643–1651.
- Eichele, K.; Ramer, R.; Hinz, B. R(+)-Methanandamide-Induced Apoptosis of Human Cervical Carcinoma Cells Involves A Cyclooxygenase-2-Dependent Pathway. Pharm. Res. 2008, 26, 346–355.
- Eichele, K.; Ramer, R.; Hinz, B. Decisive role of cyclooxygenase-2 and lipocalin-type prostaglandin D synthase in chemotherapeutics-induced apoptosis of human cervical carcinoma cells. Oncogene 2007, 27, 3032–3044.
- Ramer, R.; Heinemann, K.; Merkord, J.; Rohde, H.; Salamon, A.; Linnebacher, M.; Hinz, B. COX-2 and PPAR-γ Confer Cannabidiol-Induced Apoptosis of Human Lung Cancer Cells. Mol. Cancer Ther. 2013, 12, 69–82.
- Martínez-Martínez, E.; Martín-Ruiz, A.; Martín, P.; Calvo, V.; Provencio, M.; García, J.M. CB2 cannabinoid receptor activation promotes colon cancer progression via AKT/GSK3β signaling pathway. Oncotarget 2016, 7, 68781–68791.
- Liu, C.; Sadat, S.H.; Ebisumoto, K.; Sakai, A.; Panuganti, B.A.; Ren, S.; Goto, Y.; Haft, S.; Fukusumi, T.; Ando, M.; et al. Cannabinoids Promote Progression of HPV-Positive Head and Neck Squamous Cell Carcinoma via p38 MAPK Activation. Clin. Cancer Res. 2020, 26, 2693–2703.
- Hart, S.; Fischer, O.M.; Ullrich, A. Cannabinoids Induce Cancer Cell Proliferation via Tumor Necrosis Factor-Converting Enzyme (TACE/ADAM17)-Mediated Transactivation of the Epidermal Growth Factor Receptor. Cancer Res. 1943, 64, 1943–1950.
- Miyato, H.; Kitayama, J.; Yamashita, H.; Souma, D.; Asakage, M.; Yamada, J.; Nagawa, H. Pharmacological Synergism Between Cannabinoids and Paclitaxel in Gastric Cancer Cell Lines. J. Surg. Res. 2009, 155, 40–47.
- Aviello, G.; Romano, B.; Borrelli, F.; Capasso, R.; Gallo, L.; Piscitelli, F.; Di Marzo, V.; Izzo, A.A. Chemopreventive effect of the non-psychotropic phytocannabinoid cannabidiol on experimental colon cancer. Klin. Wochenschr. 2012, 90, 925–934.
- Romano, B.; Borrelli, F.; Pagano, E.; Cascio, M.G.; Pertwee, R.G.; Izzo, A.A. Inhibition of colon carcinogenesis by a standardized Cannabis sativa extract with high content of cannabidiol. Phytomedicine 2013, 21, 631–639.
- Winkler, K.; Ramer, R.; Dithmer, S.; Ivanov, I.; Merkord, J.; Hinz, B. Fatty acid amide hydrolase inhibitors confer anti-invasive and antimetastatic effects on lung cancer cells. Oncotarget 2016, 7, 15047–15064.
- Nithipatikom, K.; Endsley, M.P.; Isbell, M.A.; Falck, J.R.; Iwamoto, Y.; Hillard, C.J. 2-Arachidonoylglycerol: A Novel Inhibitor of Androgen-Independent Prostate Cancer Cell Invasion. Cancer Res. 2004, 64, 8826–8830.
- Ma, C.; Wu, T.-T.; Jiang, P.-C.; Li, Z.-Q.; Chen, X.-J.; Fu, K.; Wang, W.; Gong, R. Anti-carcinogenic activity of anandamide on human glioma in vitro and in vivo. Mol. Med. Rep. 2015, 13, 1558–1562.
- Ramer, R.; Hinz, B. Inhibition of Cancer Cell Invasion by Cannabinoids via Increased Expression of Tissue Inhibitor of Matrix Metalloproteinases-1. JNCI J. Natl. Cancer Inst. 2008, 100, 59–69.
- Ramer, R.; Merkord, J.; Rohde, H.; Hinz, B. Cannabidiol inhibits cancer cell invasion via upregulation of tissue inhibitor of matrix metalloproteinases-1. Biochem. Pharmacol. 2010, 79, 955–966.
- Cruz-Munoz, W.; Khokha, R. The Role of Tissue Inhibitors of Metalloproteinases in Tumorigenesis and Metastasis. Crit. Rev. Clin. Lab. Sci. 2008, 45, 291–338.
- Laezza, C.; D’Alessandro, A.; Paladino, S.; Malfitano, A.M.; Proto, M.C.; Gazzerro, P.; Pisanti, S.; Santoro, A.; Ciaglia, E.; Bifulco, M. Anandamide inhibits the Wnt/β-catenin signalling pathway in human breast cancer MDA MB 231 cells. Eur. J. Cancer 2012, 48, 3112–3122.
- García-Morales, L.; Castillo, A.M.; Ramírez, J.T.; Zamudio-Meza, H.; Domínguez-Robles, M.D.C.; Meza, I. CBD Reverts the Mesenchymal Invasive Phenotype of Breast Cancer Cells Induced by the Inflammatory Cytokine IL-1β. Int. J. Mol. Sci. 2020, 21, 2429.
- Milian, L.; Mata, M.; Alcacer, J.; Oliver, M.; Sancho-Tello, M.; De Llano, J.J.M.; Camps, C.; Galbis, J.; Carretero, J.; Carda, C. Cannabinoid receptor expression in non-small cell lung cancer. Effectiveness of tetrahydrocannabinol and cannabidiol inhibiting cell proliferation and epithelial-mesenchymal transition in vitro. PLoS ONE 2020, 15, e0228909.
- Pagano, E.; Borrelli, F.; Orlando, P.; Romano, B.; Monti, M.; Morbidelli, L.; Aviello, G.; Imperatore, R.; Capasso, R.; Piscitelli, F.; et al. Pharmacological inhibition of MAGL attenuates experimental colon carcinogenesis. Pharmacol. Res. 2017, 119, 227–236.
- Thapa, D.; Kang, Y.; Park, P.-H.; Noh, S.K.; Lee, Y.R.; Han, S.S.; Ku, S.K.; Jung, Y.; Kim, J.-A. Anti-tumor Activity of the Novel Hexahydrocannabinol Analog LYR-8 in Human Colorectal Tumor Xenograft Is Mediated through the Inhibition of Akt and Hypoxia-Inducible Factor-1α Activation. Biol. Pharm. Bull. 2012, 35, 924–932.
- Solinas, M.; Massi, P.; Cinquina, V.; Valenti, M.; Bolognini, D.; Gariboldi, M.; Monti, E.; Rubino, T.; Parolaro, D. Cannabidiol, a Non-Psychoactive Cannabinoid Compound, Inhibits Proliferation and Invasion in U87-MG and T98G Glioma Cells through a Multitarget Effect. PLoS ONE 2013, 8, e76918.