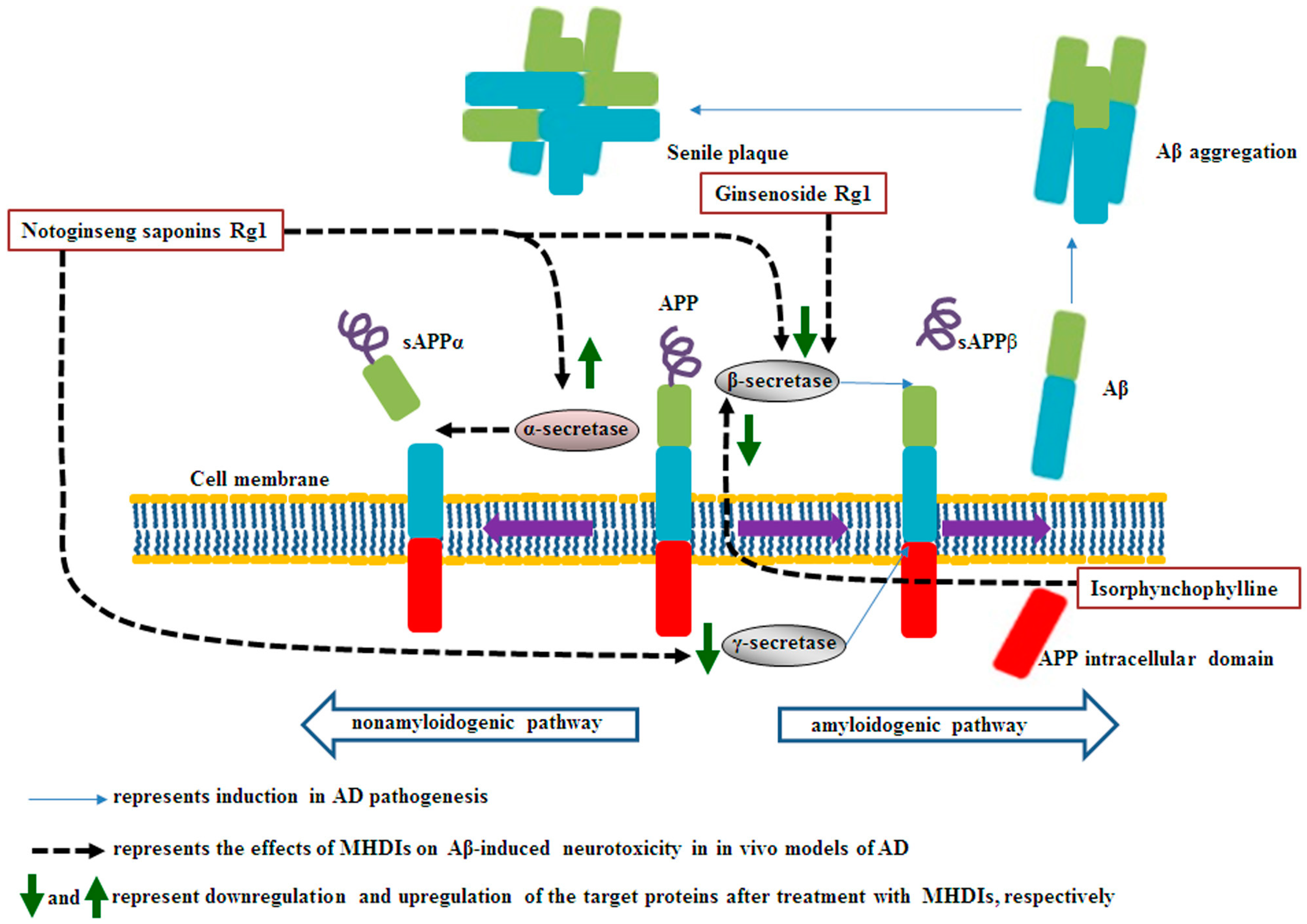
Under physiological conditions, the transmembrane glycoprotein amyloid precursor protein (APP) plays a major role in central nervous system maturation and cell contact and adhesion. However, APP overexpression can cause the production of neurotoxic derivatives, closely related to Alzheimer’s disease (AD) development [32,33]. APP can be cleaved by α-secretase to initiate the nonamyloidogenic cascade preventing Aβ accumulation. Aβ peptides are produced from APP cleavage through the activation of β- and γ-secretases in the brain regions, particularly in the temporal and frontal lobes during the early AD stages.
- Aβ Accumulation
- Alzheimer’s disease
- Aβ plague
- medicinal herb
- oxidative stress
1. Medicinal Herbs and their Derived Ingredients (MHDIs)-Mediated Suppression of Aβ Accumulation
1.1. Effects of (Medicinal Herbs and Their Derived Tngredients) MHDIs on Aβ Accumulation through α-, β-, and γ-Secretase Activity Regulation
1.2. Summary
| Major Ingredients | Isolated from Medicinal Herbs | Anti-Aβ Accumulation Activities | Models | Reference |
|---|---|---|---|---|
| Notoginseng saponin Rg1 | Panax notoginseng | α-secretase↑, β- secretase↓, γ-secretase↓ | 28 days after Aβ1–42-induced AD | [35][1] |
| Ginsenoside Rg1 | Bcl-2↑, MAP-2↑, NeuN↑, Bax↓, β-secretase↓ | 6 weeks after Aβ25–35-induced AD | [38][2] | |
| Isorphynchophylline | Uncaria tomentosa | BACE-1↓, presenilin 1↓, p-APP (Thr668) ↓ | 129 days in TgCRND8 transgenic mice | [42][7] |
Bcl-2, B-cell lymphoma 2; MAP-2, microtubule-associated protein 2; NeuN, neuronal nuclei, Bax, Bcl-2-associated x protein.
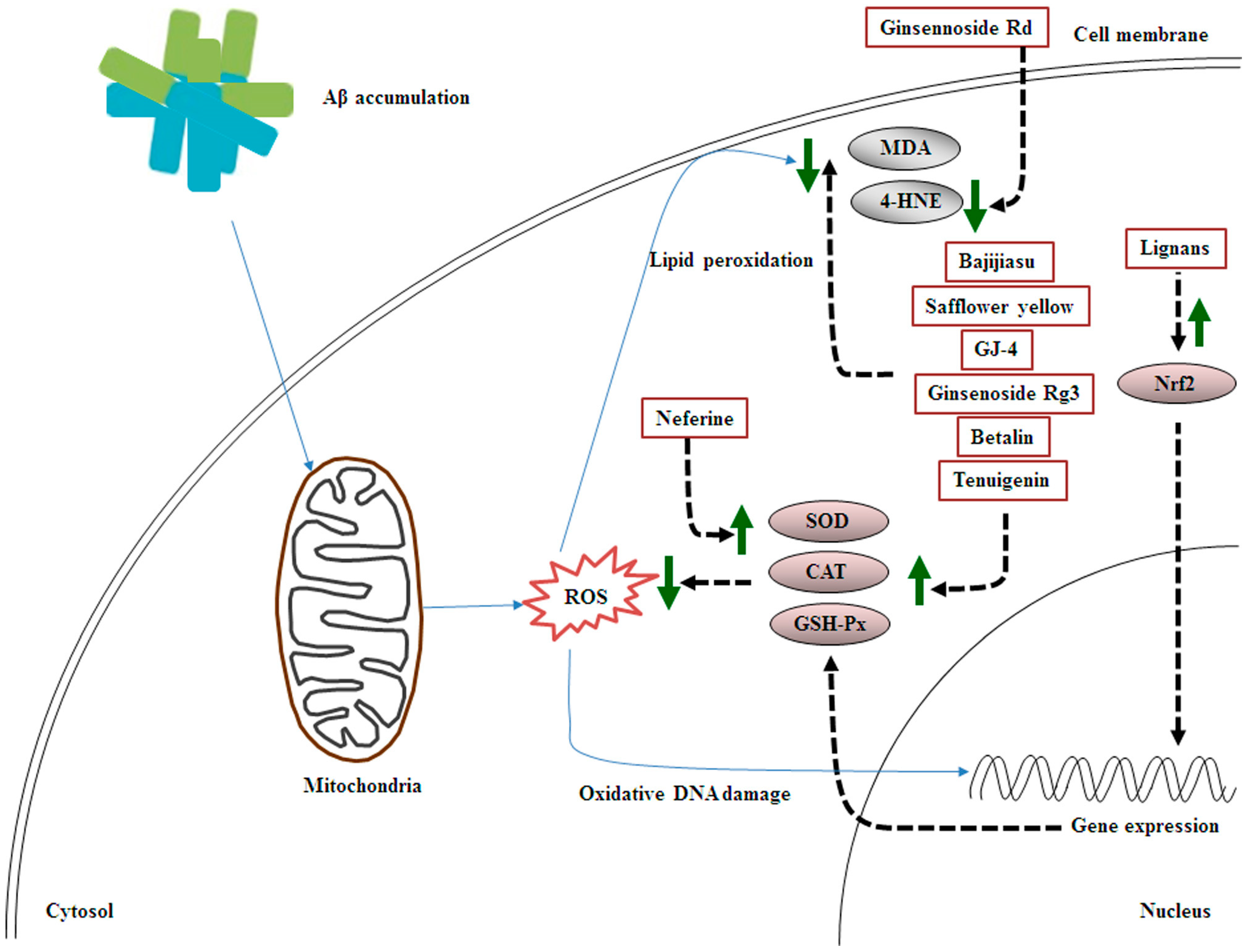
2. MHDI-Mediated Inhibition of Aβ-Induced Oxidative Stress
2.1. Involvement of Decreased Antioxidant Status and Increased Lipid Peroxidation in Aβ-Induced Oxidative Stress
2.2. Effects of MHDIs on Aβ-Induced Oxidative Stress through Antioxidant Activity and Lipid Oxidation Regulation
2.3. Summary
| Major Ingredients | Isolated from Medicinal Herbs | Antioxidative Stress Activities | Models | References |
|---|---|---|---|---|
| Ginsennoside Rd | Panax ginseng | 4-HNE↓ | 5 days after Aβ1–40-induced AD | [50][17] |
| Lignans | Schisandra chinensis Baill | kynurenic acid↑, Nrf2↑ | 28 days after Aβ25–35-induced AD | [53][20] |
| Bajijiasu | Morinda officinalis | SOD↑, CAT↑, GSH-Px↑, MDA↓ | 25 days after Aβ25–35-induced AD | [23] |
| Safflower yellow | Carthamus tinctorius | SOD↑, GSH-Px↑, MDA↓ | 28 days after Aβ1–42-induced AD | [55][24] |
| GJ-4 | Gardenia jasminoides J. Ellis | SOD↑, MDA↓, iNOS↓, COX-2↓, PGE2↓, TNF-α↓ | 10 days after Aβ25–35-induced AD | [56][25] |
| Tenuigenin | Polygala tenuifolia Willd | SOD↑, GSH-Px↑, MDA↓, 4-HNE↓ p-tau (Ser396) ↓, p-tau (Thr181) ↓ |
28 days after STZ-induced AD | [47][14] |
| Ginsenoside Rg3 | P. ginseng C. A. Meyer | SOD↑, CAT↑, GSH-Px↑, MDA↓, | 60 days after D-galactose-induced AD | [57][26] |
| Neferine | Nelumbo nucifera | SOD↑, CAT↑, GSH-Px↑ | 4 days after AlCl3-induced AD | [58][27] |
| Rhodiola crenulata | GSH-Px↓, arachidonic acid↓ | 28 days after Aβ1–42-induced AD | [31][21] | |
| Betalin | Beta vulgaris L. | SOD↑, CAT↑, GSH-Px↑, MDA↓, | 28 days after AlCl3-induced AD | [48][15] |
iNOS, inducible nitric oxide synthase; COX-2, cyclooxygenase-2; PGE2, prostaglandin E2; TNF-α, tumor necrosis factor-α.
References
- Liu, S.Z.; Cheng, W.; Shao, J.W.; Gu, Y.F.; Zhu, Y.Y.; Dong, Q.J.; Bai, S.Y.; Wang, P.; Lin, L. Notoginseng Saponin Rg1 Prevents Cognitive Impairment through Modulating APP Processing in Abeta1-42-injected Rats. Curr. Med. Sci. 2019, 39, 196–203.
- Guo, Y.; Wang, L.; Lu, J.; Jiao, J.; Yang, Y.; Zhao, H.; Liang, Z.; Zheng, H. Ginsenoside Rg1 improves cognitive capability and affects the microbiota of large intestine of tree shrew model for Alzheimer’s disease. Mol. Med. Rep. 2021, 23, 291.
- Ali, T.; Yoon, G.H.; Shah, S.A.; Lee, H.Y.; Kim, M.O. Osmotin attenuates amyloid beta-induced memory impairment, tau phosphorylation and neurodegeneration in the mouse hippocampus. Sci. Rep. 2015, 5, 11708.
- Li, J.; Du, Q.; Li, N.; Du, S.; Sun, Z. Alpiniae oxyphyllae Fructus and Alzheimer’s disease: An update and current perspective on this traditional Chinese medicine. Biomed. Pharmacother. 2021, 135, 111167.
- Vassar, R.; Kovacs, D.M.; Yan, R.; Wong, P.C. The beta-secretase enzyme BACE in health and Alzheimer’s disease: Regulation, cell biology, function, and therapeutic potential. J. Neurosci. 2009, 29, 12787–12794.
- Neumann, U.; Ufer, M.; Jacobson, L.H.; Rouzade-Dominguez, M.L.; Huledal, G.; Kolly, C.; Luond, R.M.; Machauer, R.; Veenstra, S.J.; Hurth, K.; et al. The BACE-1 inhibitor CNP520 for prevention trials in Alzheimer’s disease. EMBO Mol. Med. 2018, 10, e9316.
- Li, H.Q.; Ip, S.P.; Yuan, Q.J.; Zheng, G.Q.; Tsim, K.K.W.; Dong, T.T.X.; Lin, G.; Han, Y.; Liu, Y.; Xian, Y.F.; et al. Isorhynchophylline ameliorates cognitive impairment via modulating amyloid pathology, tau hyperphosphorylation and neuroinflammation: Studies in a transgenic mouse model of Alzheimer’s disease. Brain Behav. Immun. 2019, 82, 264–278.
- McDade, E.; Voytyuk, I.; Aisen, P.; Bateman, R.J.; Carrillo, M.C.; De Strooper, B.; Haass, C.; Reiman, E.M.; Sperling, R.; Tariot, P.N.; et al. The case for low-level BACE1 inhibition for the prevention of Alzheimer disease. Nat. Rev. Neurol. 2021, 17, 703–714.
- Xiang, X.; Wang, X.; Jin, S.; Hu, J.; Wu, Y.; Li, Y.; Wu, X. Activation of GPR55 attenuates cognitive impairment and neurotoxicity in a mouse model of Alzheimer’s disease induced by Abeta1-42 through inhibiting RhoA/ROCK2 pathway. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2022, 112, 110423.
- Ahmad, S.; Jo, M.H.; Ikram, M.; Khan, A.; Kim, M.O. Deciphering the Potential Neuroprotective Effects of Luteolin against Abeta1-42-Induced Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 9583.
- Feng, S.T.; Wang, Z.Z.; Yuan, Y.H.; Sun, H.M.; Chen, N.H.; Zhang, Y. Mangiferin: A multipotent natural product preventing neurodegeneration in Alzheimer’s and Parkinson’s disease models. Pharmacol. Res. 2019, 146, 104336.
- Kim, J.H.; Lim, D.K.; Suh, Y.H.; Chang, K.A. Long-Term Treatment of Cuban Policosanol Attenuates Abnormal Oxidative Stress and Inflammatory Response via Amyloid Plaques Reduction in 5xFAD Mice. Antioxidants 2021, 10, 1321.
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell Longev. 2019, 2019, 9613090.
- Huang, X.B.; Chen, Y.J.; Chen, W.Q.; Wang, N.Q.; Wu, X.L.; Liu, Y. Neuroprotective effects of tenuigenin on neurobehavior, oxidative stress, and tau hyperphosphorylation induced by intracerebroventricular streptozotocin in rats. Brain Circ. 2018, 4, 24–32.
- Shunan, D.; Yu, M.; Guan, H.; Zhou, Y. Neuroprotective effect of Betalain against AlCl3-induced Alzheimer’s disease in Sprague Dawley Rats via putative modulation of oxidative stress and nuclear factor kappa B (NF-kappaB) signaling pathway. Biomed. Pharmacother. 2021, 137, 111369.
- Xiong, Z.; Liu, C.; Wang, F.; Li, C.; Wang, W.; Wang, J.; Chen, J. Protective effects of breviscapine on ischemic vascular dementia in rats. Biol. Pharm. Bull. 2006, 29, 1880–1885.
- Liu, J.; Yan, X.; Li, L.; Zhu, Y.; Qin, K.; Zhou, L.; Sun, D.; Zhang, X.; Ye, R.; Zhao, G. Ginsennoside rd attenuates cognitive dysfunction in a rat model of Alzheimer’s disease. Neurochem. Res. 2012, 37, 2738–2747.
- Ashabi, G.; Alamdary, S.Z.; Ramin, M.; Khodagholi, F. Reduction of hippocampal apoptosis by intracerebroventricular administration of extracellular signal-regulated protein kinase and/or p38 inhibitors in amyloid beta rat model of Alzheimer’s disease: Involvement of nuclear-related factor-2 and nuclear factor-kappaB. Basic Clin. Pharmacol. Toxicol. 2013, 112, 145–155.
- Ferreira, F.S.; Biasibetti-Brendler, H.; Pierozan, P.; Schmitz, F.; Berto, C.G.; Prezzi, C.A.; Manfredini, V.; Wyse, A.T.S. Kynurenic Acid Restores Nrf2 Levels and Prevents Quinolinic Acid-Induced Toxicity in Rat Striatal Slices. Mol. Neurobiol. 2018, 55, 8538–8549.
- Zhou, Y.; Wei, M.; Fan, M.; Liu, Z.; Wang, A.; Liu, Y.; Men, L.; Pi, Z.; Liu, Z.; Song, F. Pharmacokinetic and metabolomics approach based on UHPLC-MS to evaluate therapeutic effect of lignans from S. Chinensis in alzheimer’s disease. J. Chromatogr. B 2021, 1178, 122859.
- Li, X.; Wang, Y.; Su, M.; Chu, X.; Li, S.; Yue, Y.; Zhang, X.; Wang, J.; Han, F. Brain metabolomics study for the protective effects of Rhodiola crenulata extract on Alzheimer’s disease by HPLC coupled with Fourier transform-ion cyclotron resonance mass spectrometry. J. Sep. Sci. 2020, 43, 3216–3223.
- Duffy, S.L.; Lagopoulos, J.; Hickie, I.B.; Diamond, K.; Graeber, M.B.; Lewis, S.J.; Naismith, S.L. Glutathione relates to neuropsychological functioning in mild cognitive impairment. Alzheimers Dement. 2014, 10, 67–75.
- Chen, D.L.; Zhang, P.; Lin, L.; Zhang, H.M.; Deng, S.D.; Wu, Z.Q.; Ou, S.; Liu, S.H.; Wang, J.Y. Protective effects of bajijiasu in a rat model of Abeta(2)(5)(-)(3)(5)-induced neurotoxicity. J. Ethnopharmacol. 2014, 154, 206–217.
- Ma, Q.; Ruan, Y.Y.; Xu, H.; Shi, X.M.; Wang, Z.X.; Hu, Y.L. Safflower yellow reduces lipid peroxidation, neuropathology, tau phosphorylation and ameliorates amyloid beta-induced impairment of learning and memory in rats. Biomed. Pharmacother. 2015, 76, 153–164.
- Zang, C.X.; Bao, X.Q.; Li, L.; Yang, H.Y.; Wang, L.; Yu, Y.; Wang, X.L.; Yao, X.S.; Zhang, D. The Protective Effects of Gardenia jasminoides (Fructus Gardenia) on Amyloid-beta-Induced Mouse Cognitive Impairment and Neurotoxicity. Am. J. Chin. Med. 2018, 46, 389–405.
- Zhang, Y.; Yang, X.; Wang, S.; Song, S. Ginsenoside Rg3 Prevents Cognitive Impairment by Improving Mitochondrial Dysfunction in the Rat Model of Alzheimer’s Disease. J. Agric. Food Chem. 2019, 67, 10048–10058.
- Yin, S.; Ran, Q.; Yang, J.; Zhao, Y.; Li, C. Nootropic effect of neferine on aluminium chloride-induced Alzheimer’s disease in experimental models. J. Biochem. Mol. Toxicol. 2020, 34, e22429.