Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Bartłomiej Barczyński and Version 2 by Dean Liu.
The eubiotic vaginal microbiota of reproductive-aged women is composed mostly of various Lactobacillus species (spp.), which exert protective effects via the production of lactic acid, bacteriocins, polysaccharides, peptidoglycans, and hydrogen peroxide (H2O2), lowering pH, raising the viscosity of cervicovaginal mucus, and hampering both the adhesion of cells to epithelial tissue and the entry of HPV. The depletion of beneficial microorganisms could increase the risk of sexually transmitted infections. Emerging therapies involve mucosal, intranasal vaccines, which trigger systemic and mucosal immune responses, thus protecting against HPV-induced tumours.
- cervical cancer
- endometrial cancer
- dysbiosis
- Lactobacillus
1. The Microbial Environment of the Vagina and Upper Reproductive Tract
The female reproductive tract is inhabited by various coexisting microorganisms, which influence health or disease states [1][30]. The vaginal microbiota in healthy women of reproductive age is not diverse and usually comprises one or few Lactobacillus spp. [2][3][26,31]. During eubiosis, the vaginal microbiota of reproductive-aged women is primarily composed of various Lactobacillus spp., including Lactobacillus gasseri, Lactobacillus crispatus, Lactobacillus jensenii, and Lactobacillus iners [3][4][5][31,32,33]. Studies have shown that the depletion of beneficial microorganisms could be associated with a higher risk of sexually transmitted infections, pelvic inflammatory disease, preterm births, and spontaneous miscarriages [6][34]. According to studies, the profile of each female vaginal microbiome can be classified into six community state types (CSTs) [7][8][3,35]. Lactobacillus, especially L. crispatus, L. gasseri, L iners, and L. jensenii, is predominant in CST-I, II, and III, Streptococcus and Prevotella dominate in CST IV-A, and Atopobium is highly prevalent in CST IV-B. The presence of bacteria belonging to CST-IV is frequently associated with bacterial vaginosis. The aforementioned Lactobacillus species appear to be adapted for dominance in the vaginal niche since other types of Lactobacillus are not observed there [9][10][36,37]. The explanation of this phenomenon is unknown; however, it could be related to evolutionary issues [11][38]. The predominance of vaginal Lactobacillus spp. protects this microenvironment against the invasion of pathogens. It has been observed that Gardnerella vaginalis can also be dominant in the vaginal microbiome. The vaginal microbiome that is non-Lactobacillus-dominant appears to be more frequent in Hispanic and Black women (30–40%) compared to White and Asian women (10–20%) [12][13][14][39,40,41]. Ethnic and racial disparities can stem from different environmental and socioeconomic factors as well as diverse behaviour, e.g., sexual and hygiene-related [15][42]. However, some reports have indicated that at least one Lactobacillus can be related to disease states. For example, L. iners was identified in females with disorders of the vaginal environment [16][17][18][43,44,45]. The presence of L. iners-dominant vaginal microbiome is frequently observed during the transition to the non-Lactobacillus-dominant communities [19][46]. The vaginal microbiome can be affected by numerous factors, including infections with HPV and other STIs, sexual activity, lubricant use, the number of sexual partners, contraception use, hygiene practices, access to health care, diet and nutrition (fat-rich diet and high glycaemic load), smoking, physical activity, obesity, and alcohol consumption. Age; genetic and epigenetic factors; hormone levels; pregnancy; immune system impairment; stress; and exposure to xenobiotics, carcinogens, toxins, and antibiotics also influence its composition [20][21][22][47,48,49]. The vaginal microbiota profile depends on ethnicity; the Lactobacillus species are more prevalent in Caucasian and Asian women compared to Hispanic and Black women [7][3]. The ethnic differences in microbiota can be associated with either genetic factors affecting mucosal immunity and metabolic pathways or hygiene practices [7][3]. The gut microbiome has been demonstrated to indirectly influence the abundance of Lactobacillus in the vaginal microenvironment via the modulation of oestrogen release, which may imply the existence of a gut–vaginal axis [23][24][25][50,51,52]. β-glucuronidase and β-glucosidase secreted by microorganisms attach to oestrogen, thus leading to its enhanced reabsorption into the circulation [26][27][53,54]. In turn, unbound oestrogen reaches the female reproductive tract where it activates intracellular signalling associated with increased glycogen syntheses, thickening of the genital epithelium, and the production of mucus. Thus, females’ hormones, including oestrogen and progesterone, modulate vaginal colonisation with Lactobacillus spp. Higher levels of these hormones are associated with lower vaginal microbiota diversity and the dominance of Lactobacillus [28][29][55,56]. The relationship between oestrogen levels and the amount of vaginal Lactobacillus is mirrored by the finding of reduced Lactobacillus abundance in females before menstruation, i.e., when oestrogen levels are significantly reduced [30][31][57,58]. In this period of decreased oestrogen levels, some species become enriched, while others are depleted in the vaginal environment [8][35]. Temporal oestrogen deficiency may cause vaginal atrophy, which is partly responsible for higher bacterial diversity [8][35]. The decrease in the lactic acid bacteria pool is associated with the predominance of anaerobic bacteria and the subsequent risk of cervical cancer development. Though the mechanisms underlying hormone-related microbial composition of the vagina are not fully understood, it has been suggested that the dominance of Lactobacillus spp. may be associated with the oestrogen-driven maturation of vaginal epithelium, the production of α-amylase, and the accumulation of glycogen [32][59]. The degradation of glycogen by α-amylase to simple products such as maltose, maltotriose, maltotetraose, and α-dextrins promotes Lactobacillus growth and colony formation [33][60]. The use of synthetic hormones, e.g., contraceptives, has also been reported to decrease the incidence or recurrence of bacterial vaginosis [34][61]. In turn, smoking, sexual intercourse, and vaginal douching appear to diminish the abundance of L. crispatus, increase species diversity, and enhance the risk of bacterial vaginosis [35][36][37][62,63,64]. Data concerning common microbiota inhabiting the uterus, fallopian tubes, or ovaries are limited due to problems with its assessment [38][65]. The microbiota of the female upper reproductive tract was found to be very different from that of the vagina in composition and quantity [15][42]. Chen et al. [39][66] suggested that the number of bacteria in the uterus could be ~10,000-fold lower compared to the number of bacteria in the vagina. However, this estimation could be inexact due to the high risk of cross-contamination with bacteria from the lower part of the tract during transcervical collection. Moreover, it has been suggested that upper reproductive tract microbiota are more diverse compared to that of the lower tract; however, genuine members have not been identified since various studies indicated different microbiota compositions [39][66]. Lactobacillus species were also found in the upper tract, but their abundance gradually reduced with its withdrawal from the vagina and cervix. Numerous studies revealed that various body sites can serve as possible reservoirs of genital microorganisms. For example, common vaginal bacteria, including Lactobacillus, Gardnerella, Sneathia, Prevotella, Atopobium, Gemella, Peptoniphilus, and Finegoldia, are normally found in the urinary tract in both women and men [40][41][42][67,68,69]. Thomas-White et al. [42][69] observed that vaginal and bladder microbiota displayed comparable functional capacities, which differed from gut microbiota. The presence of Lactobacillus spp. in the bladder and the vagina could exert a protective effect against invading uropathogens. Moreover, the co-colonisation of both the vagina and rectum with vaginal Lactobacillus species, including L. crispatus, L. jensenii, L. iners, and L. gasseri, was associated with the lowest prevalence of bacterial vaginosis [43][44][70,71]. Therefore, it was suggested that the rectum might be a vital reservoir for vaginal lactobacilli. The presence of the vaginal microbiome’s members on male penile skin, in semen, and in urine specimens may imply that sexual partners can exchange microbiota residing in their urogenital tracts [45][72]. According to some studies, the composition of endometrial microbiota may affect implantation, pregnancy, and live birth rates [46][73]. Lactobacillus-dominated endometrial fluid and vaginal aspirate correlate with better outcomes. Uterine microbiota was suggested to exert an impact on the immune environment during conception [47][74]. Modifications of microbial composition in the endometrial fluid can elicit an inflammatory response within the endometrium, thus lowering the probability of embryo implantation success [48][75].2. The Role of Lactobacillus in the Female Reproductive Tract
In contrast to many parts of the body in which great microbial diversity appears to be beneficial, in the vagina, a higher diversity of microbiota frequently results in dysbiosis and the development of disease states. Many studies have demonstrated that vaginal microbiota, including Lactobacillus, is involved in the protection of the reproductive tract and gastrointestinal tract against opportunistic infections [49][50][1,7]. The ability of Lactobacillus to produce lactic acid via the fermentation of glucose (glycolysis) supports vaginal eubiosis, as this organic acid helps preserve the vaginal acidic environment [51][76]. The acidic environment constrains the growth of some potentially pathogenic species, including C. trachomatis, G. vaginalis, and Neisseria gonorrhoeae [4][52][53][54][32,77,78,79]. Vaginal pH exceeding 5.0 was found to increase the risk of HPV in premenopausal women by 10–20% [55][80]. This finding could be partly explained by the fact that the HPV protein crucial for viral transformation, E5, is vulnerable to low pH [56][81]. Moreover, it offers optimal conditions for the metabolic functioning of cervical and vaginal cells [57][82]. Apart from affecting the pH of the environment, the chemical structure of lactic acid itself may modulate the HPV infection and the development of squamous intraepithelial lesions [7][3]. As a chiral molecule, lactic acid can be produced in the form of D- and L-isomers. Studies demonstrated that high levels of D-lactic acid could protect against Chlamydia infection and upper reproductive tract infections via the modulation of extracellular matrix metalloproteinase inducer (EMMPRIN) production in vaginal epithelial cells [58][59][83,84]. A higher L-lactate-to-D-lactate ratio is associated with the enhanced expression of EMMPRIN as well as the activation of matrix metalloproteinase 8 (MMP-8), eventually resulting in impaired cervical integrity and the easier entry of HPV into basal keratinocytes [58][83]. Nunn et al. [60][85] revealed that the predominance of L. crispatus and relatively high levels of D-lactic acid could increase the viscosity of cervicovaginal mucus, resulting in viral particle trapping. Lactic acid also limits the cytotoxicity of natural killer (NK) cells, diminishes the synthesis of pro-inflammatory cytokine IL-12, and promotes the release of anti-inflammatory interleukin-10 (IL-10) [61][62][86,87]. Apart from lactic acid, beneficial microbiota can also release other antimicrobial peptides, including bacteriocins and hydrogen peroxide (H2O2) [63][64][88,89]. Bacteriocins exert direct bactericidal effects, but they can also modulate the inflammatory immune response and mediate acquired immune response [49][1]. They possess anti-tumour properties resulting from cytotoxicity and the stimulation of cell lysis. Gassericin (bacteriocin), produced by L. gasseri as well as other strains of L. crispatus and Lactobacillus reuteri, acts on Gram-negative and Gram-positive bacteria [65][66][90,91]. Apart from bacteriocins, some bacteria (e.g., Lactobacillus) can also release biosurfactants, which modify surface tension, therefore hampering bacterial adhesion, biofilm formation, and the excessive growth of pathogenic anaerobes [67][92]. Lactobacillus epithelium adhesin (LEA), produced by L. crispatus, prevents the pilus-mediated adhesion of G. vaginalis [68][93]. The aforementioned bacteriocins and biosurfactants have also been demonstrated to disturb viral infiltration [69][94]. Moreover, both bacteriocin and surface-active components can constrain the synthesis of tumourigenic substances [70][95]. A higher rate of bacterial vaginosis was reported in females with decreased vaginal levels of bacteria capable of producing H2O2 [71][96]. The release of a variety of antimicrobial peptides (AMPs) into the uterine cavity poses a vital defence mechanism, protecting epithelial tissues against proteolytic enzymes secreted by pathogens [72][73][97,98]. Some studies have suggested that hypoxia could also promote the development of bacterial vaginosis since, in such conditions, bacteria are not able to produce H2O2 in a sufficient amount to inhibit pathogenic bacteria growth [74][75][99,100]. The interaction of commensal bacteria with endometrial epithelial cells was found to form an antimicrobial barrier against pathogens [76][101]. The presence of Lactobacillus in the vagina is associated with protection against the adherence of pathogenic bacteria to the epithelial tissue. These bacteria compete against pathogenic microorganisms for territories and nutrients [77][102]. Lactobacillus that occupies the vaginal epithelial cells (VECs) has been found to prevent the conglutination of invasive pathogenic bacteria, thus hampering the initiation of malignant tumours [78][79][103,104]. Lactobacillus was demonstrated to hinder the proliferation of malignant tumours via the secretion of phosphorylated polysaccharides, exopolysaccharides, and peptidoglycans [62][80][87,105]. Moreover, these bacteria can stimulate nitric oxide (NO) production by macrophages and impair energy metabolism in cancer cells [81][106]. Commensal bacteria stimulate the production of neutral, stable mucous by endometrial cells as well as preserve tight junctions [38][82][65,107]. An intact epithelial barrier is crucial for protection against the penetration and colonisation of opportunistic microorganisms. Furthermore, commensal bacteria can modify immune responses at the cellular level [76][101]. Studies have demonstrated that Lactobacillus enhances the proliferation and differentiation of thymus-derived cells (T cells) and ameliorates the immunological recognition and proliferation of B cells [83][84][108,109]. The adhesion of Lactobacillus and the absorption of nutrients have been demonstrated to trigger the complement system, which subsequently regulates microbial growth [85][110].
Motevaseli et al. [86][111] demonstrated that vaginal lactobacilli (L. gasseri and L. crispatus) could exert cytotoxic impact on cervical tumour cells, however, normal cells remained unaffected. Moreover, they observed that this effect was independent of lactic acid and pH. Studies have demonstrated the antimetastatic and antiproliferative properties of Lactobacillus, its subgenera, and its supernatants [62][87]. Via the modulation of HPV oncogenes, Lactobacillus was shown to limit cervical cancer cell viability. Another study has implied that L. crispatus is highly resistant to the co-colonisation of other bacteria and the transition into CST IV [19][46]. These bacteria are rarely found to coexist with other species. Furthermore, females with these bacteria have the lowest vaginal pH and are not susceptible to infections with bacterial STIs, HPV, herpes simplex virus-2 (HSV-2), or HIV [3][87][31,112]. Since bacterial vaginosis promotes the shedding of HIV and HSV-2, it has been suggested that dysbiosis and the reduced abundance of Lactobacillus may support the formation of an environment that induces the persistence of infections and leads to the development of squamous intraepithelial lesions [88][113]. The basic beneficial effects of Lactobacillus in the lower female genital tract are presented in Figure 1.
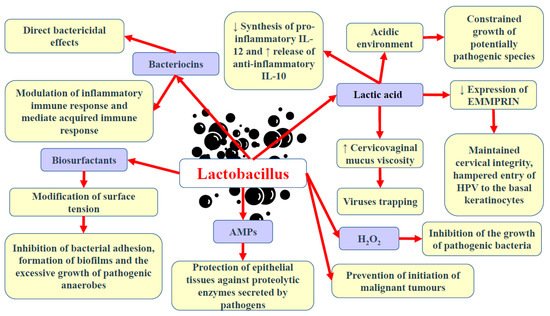
Figure 1. Basic beneficial mechanisms of Lactobacillus in the female genital tract. Abbreviations: ↓—decrease; ↑—increase; AMPs—antimicrobial peptides; H2O2—hydrogen peroxide; EMMPRIN—extracellular matrix metalloproteinase inducer; IL-10 and -12—interleukin-10 and -12.