Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Peng Gao and Version 2 by Rita Xu.
Amino acids (AAs) are indispensable building blocks of diverse bio-macromolecules as well as functional regulators for various metabolic processes. The fact that cancer cells live with a voracious appetite for specific AAs has been widely recognized. Glioma is one of the most lethal malignancies occurring in the central nervous system. The reprogrammed metabolism of AAs benefits glioma proliferation, signal transduction, epigenetic modification, and stress tolerance. Metabolic alteration of specific AAs also contributes to glioma immune escape and chemoresistance. For clinical consideration, fluctuations in the concentrations of AAs observed in specific body fluids provides opportunities to develop new diagnosis and prognosis markers.
- amino acid
- glioma
- metabolomics
- metabolism
1. Introduction
Amino acids (AAs) are the exclusive building blocks for proteins in both the eukaryotes and the prokaryotes. It is estimated that over 500 AAs could be found in different forms of lives [1], but only 20 AAs were implicated in protein synthesis. The proteogenic AAs share a similar chemical structure with a carboxyl group and an amino group, together with a hydrogen atom, bonded to the α-carbon atom. The physical and chemical properties of different AAs are determined by the relevant side chain groups which are also covalently linked to the α-carbon atom. Owning to the chirality of the α-carbon atom, each AA has two optical isomers in theory. Interestingly, the 20 proteogenic AAs are all L-enantiomers. Previously, it was believed that AAs of D-enantiomers were absent in mammals. However, mounting evidence demonstrated that D-AAs could also be found in higher animals. For example, D-serine and D-aspartate were abundant in the central nervous system (CNS) of some vertebrates [2][3][2,3]. This review would pay more attention to L-AAs.
Except for protein synthesis, AAs have diverse functions under different circumstances [4]: (a) Some AAs could be converted to corresponding α-ketonic acids by aminotransferases. These α- ketonic acids could be catalyzed to glucose, fats or ketone bodies to be utilized for various purposes. The relevant AAs are called ketogenic or glycogenic AAs. (b) Some AAs behave as neurotransmitters, precursors for many hormones or other bioactive molecules. For example, glutamate is an excitatory AA in the CNS [5]. One of the enzymatic products of glutamate, the γ-aminobutyric acid, is an inhibitory neurotransmitter [6]. (c) Glycine, serine, methionine and histidine could provide one-carbon units, promoting purine, pyrimidine, and the redox regulator biosynthesis [7][8][7,8]. (d) Some AAs have been utilized as potential biomarkers to aid diagnosis and prognosis. For instance, elevated circulating branched-chain AAs (BCAAs) indicate a higher risk of pancreatic cancer [9]. Individuals with elevated serum concentrations of 9 AAs showed a higher incidence of Type 2 diabetes [10]. (e) AAs could behave as epigenetic modifiers to regulate cell phenotypes. Sarcosine played roles in prostate cancer diagnosis and stratification [11][12][11,12], and this AA also brought about increased methylated CpG island landscapes in several prostate cancer cell lines [13]. Collectively, AAs are multi-functional molecules.
Metabolites directly affect and regulate cellular phenotypes [14][15][14,15]. Cell transformation (tumorigenesis) is accompanied by metabolic reprogramming [16]. A half-century ago, Warburg published his profound work pointing out that tumor cells consumed a large part of glucose through glycolysis even under the circumstance of sufficient oxygen supply [17][18][17,18]. Several lines of evidence demonstrated that metabolic reprogramming is involved in diverse metabolic pathways when the cell transformed [19].
Glioma is one of the lethal malignancies in the CNS and accounts for one-third of the primary brain tumors [20]. The World Health Organization primarily stratified gliomas into four histopathologic grades. The higher the grade, the less favorable the prognosis [21]. The exact pathological mechanisms of gliomas are still elusive. A deep insight into the metabolic adaptation of gliomas is of benefit to exploring new therapeutic solutions.
2. Glioma AA Metabolism Adapted to Proliferation
One of the hallmarks of cancer cell metabolism is their uncontrollable proliferation abilities. This does not mean the malignant cells adopted some metabolic pathways that were unique to tumor cells. Most frequently, cancer cells change certain enzymes’ expression or activities to meet their abnormal catabolic and anabolic requirements [19]. The relevant mechanisms include the mutations of specific enzyme genes, the accumulation of specific activators/inhibitors, the altered post-translational modification activities and the over-activated/inhibited regulation signals. One-carbon unit metabolism is closely linked to many biological processes. The production of ATP, NADPH, lipids and nucleotides greatly relies on one-carbon units, especially the de novo synthesis of purine. Cell proliferation is largely determined by the availability of nucleotides [22]. Jain, M et al. profiled the extracellular consumption and release of 219 metabolites for 60 primary human cancer cell lines including U251 glioblastoma (GBM) cells [23][24][23,24]. They found glycine was consumed massively by rapidly proliferating cells and released by slowly proliferating ones. The uptake of extracellular glycine is mediated by the transporter of GLYT1 [25]. The intracellular biosynthesis of glycine could occur both in the cytosol and the mitochondria. Most cells default to the mitochondria for glycine synthesis [26]. Mitochondrial serine hydroxymethyltransferase 2 (SHMT2) catalyzes the reversible transformation from glycine to serine [27]. SHMT2 was frequently found to be overexpressed in GBM tissues [28][29][28,29]. Isotope tracing analysis indicated mitochondria contribute about two-thirds of the needed glycine to rapidly proliferating glioma cells [24]. The consumed glycine was either converted to one-carbon units to support purine synthesis or incorporated into glutathione (GSH) to clear reactive oxygen species (ROS) [30][31][30,31]. Glycine decarboxylase (GLDC) plays a key role in converting glycine into one-carbon units. The activity of GLDC is regulated by acetylation modification. This post-translational modification of GLDC is inhibited by the mechanistic target of rapamycin complex 1 (mTORC1). Acetylated GLDC is prone to be degraded in the proteasomes and results in impaired pyrimidine synthesis and growth inhibition of gliomas [32]. Many glioma cells exhibit highly expressed mTORC1 and GLDC [32][33][32,33]. γ-Glutamylcyclotransferase (GGCT), one of the key enzymes promoting GSH synthesis, was demonstrated to be highly expressed in glioma cells. Suppression or depletion of GGCT compromised glioma cells but not normal cells’ proliferation [34][35][34,35], implying the different GSH-related vulnerabilities of the normal and the transformed cells. In addition to GGCT, SHMT2 also promoted GSH synthesis [26]. Collectively, glycine confers gliomas a proliferative advantage by providing one-carbon units and reduction substances (Figure 1). The voracious appetite for glycine of gliomas renders glycine a promising imaging tracer to aid glioma aggressiveness evaluation [36]. Additionally, one-carbon metabolism has been identified as one of the potential targets for treating GBM [37].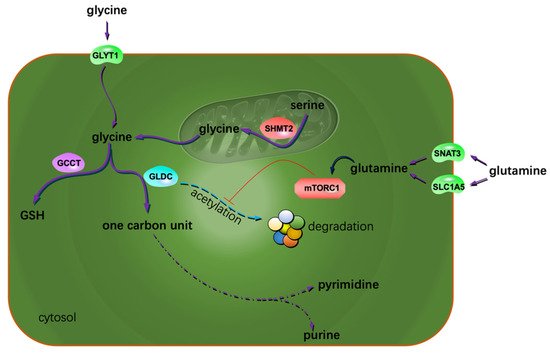
Figure 1. Extra glycine and glutamine sustain glioma proliferation. The possible mechanism is that both the AAs contribute to the metabolism of one carbon units and the reduction substances. The arrow with a blunt end indicates suppression.
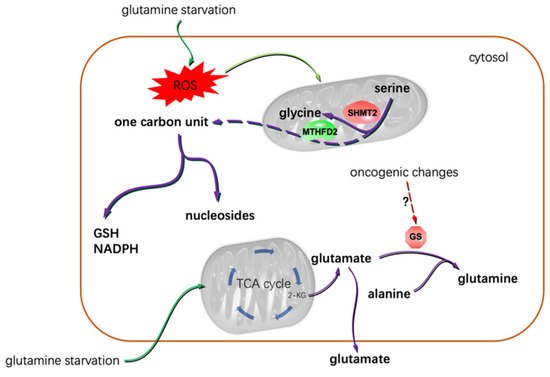
Figure 2. Glutamine starvation resulted in the dependence of glioma cells on serine-based one carbon unit metabolism, and that gliomas showed differential glutamine addiction was most possibly due to the varied oncogenic backgrounds of the cells. Under glutamine deprivation conditions, some glioma cells even excreted glutamate [49]. 2-KG: α-ketoglutarate.
3. Glioma Migration, Invasion, and AA Metabolism
Although the metastasis of glioma is seldom found in patients, the infiltrative growth feature of gliomas is very harmful. Thus, understanding the mechanisms of glioma invasion and migration would benefit patient care. System Xc− is an Na+-independent antiporter. It mediates the exchanging of extracellular cystine and intracellular glutamate [64]. Most gliomas highly express system Xc−. Glutamate excreted by gliomas is cytotoxic to the adjacent normal brain cells. Through this, gliomas create extra space to expand and invade. Lyons S. et al. reported that glutamate could activate Ca2+-permeable α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptors (AMPA-Rs) in the manner of autocrine and/or paracrine signaling. Activation of AMPA-Rs resulted in intracellular Ca2+ oscillation, one of the essential signals to trigger glioma invasion and migration (Figure 3). Most of the glioma tissues highly express AMPA-Rs. Agonists of AMPA-Rs facilitated glioma invasion and migration [65]. Additionally, glutamate showed a high affinity to N-methyl-D-aspartate receptors. Crosstalk between AMPA-Rs and N-methyl-D-aspartate receptors has been demonstrated to synergistically promote glioma invasion, especially in a glutamate-rich microenvironment. BCAA transaminase 1 (BCAT1) initiates the breakdown of BCAAs. Some catabolic products of BCAAs could be used as the carbon skeleton for glutamate synthesis. Inhibition of BCAT1 resulted in a decreased efflux of glutamate and brought about the impaired invasiveness of glioma cells [66]. In conclusion, except for its cytotoxic effects, glutamate could act as a hormone to stimulate invasion and migration (Figure 3) [67].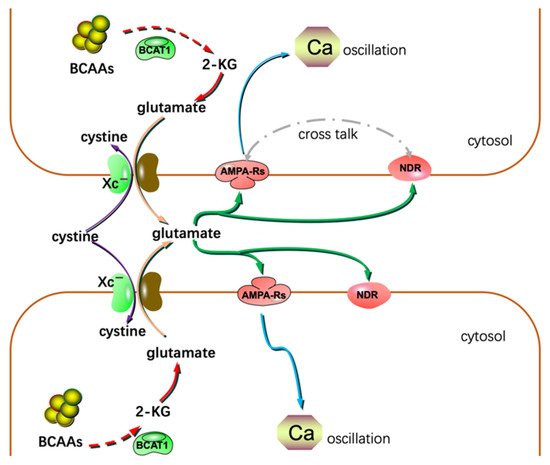
Figure 3. Glutamate could act as a hormone to stimulate invasion and migration in addition to its cytotoxic effects. NDR: N-methyl-D-aspartate receptor; 2-KG: α-ketoglutarate.