Mycobacterium avium subspecies paratuberculosis (MAP) is an environmentally hardy pathogen of ruminants that is transmitted via the fecal-oral route. Transition from subclinical to clinical infection is a dynamic process led by MAP, which survives and replicates in host macrophages. Hallmark clinical symptoms include granulomatous enteritis, watery diarrhea, and significant loss of body condition. Clinical stage disease is accompanied by dysfunctional immune responses and a reduction in circulating vitamin D3. The immunomodulatory role of vitamin D3 in infectious disease has been well established in humans, particularly in Mycobacterium tuberculosis infection. However, significant species differences exist between the immune system of humans and bovines, including effects induced by vitamin D3.
- vitamin D
- Mycobacterium avium subsp. paratuberculosis
- cattle
- macrophage
- PBMC
- endosomal trafficking
1. Johne’s Disease Overview
2. Transmission
MAP infection rates in young calves increase with exposure to larger numbers of MAP, and infection is most commonly established via the fecal–oral route for upwards of a year following birth [11][12]. Transmission of MAP through colostrum and milk by infected dams has been shown to occur and detection rates, along with bacterial burden, appear to coincide with the level of fecal shedding by the dam [13]. Further, rates of in utero transmission also appear to relate to disease severity and MAP burden in the feces [14][15]. These routes of infection represent important steps in biosafety protocol during efforts of herd infection control, and largely revolve around maintaining clean, dry housing environments. Fecal shedding patterns are intermittent in animals naturally infected with MAP categorized in the subclinical stage of disease. Dynamics of fecal shedding have been shown to be largely different between experimentally and naturally infected cattle, with experimentally infected animals shedding after one year and alternating between positive and negative status several times while also fluctuating between high and low levels [16]. Most of these animals reached high levels of shedding, which could be impacted by MAP infection dose and administration route. This sentudry also showed those animals that were naturally infected at a young age had a propensity to begin shedding approximately 3 years following infection, had little to no switching between shedding status, and those that did shed intermittently had a reduced chance of becoming a high shedder compared to those who shed MAP consistently. Other reports have also shown similar disease outcomes related to shedding level and frequency [17]. Additionally, naturally infected cattle that began shedding high levels frequently continued to do so until being removed from the herd [16]. Cattle that progress to clinical stage paratuberculosis show symptoms including watery diarrhea, weight loss, and in severe cases submandibular edema. Gross pathology shows marked inflammation of the intestinal wall, which impairs nutrient absorption from the diet [18]. This can lead to hypoproteinemia, reducing oncotic pressure of capillary vessels allowing fluid to leak from the blood vessels and accumulate beneath the jaw in grazing animals. The clinical stage is often accompanied by shedding of high levels of MAP and a significant reduction in milk production for lactating animals [19].3. Immune Responses to MAP
Macrophage phenotype and resulting effector functions are influenced by the cell’s cytokine milieu, pathogen signaling, and immune cell interactions. Phenotypic differences can be broadly represented by two groups according to cell surface receptors and cytokines produced [20]. A pro-inflammatory T helper 1 (Th1) cytokine profile, namely including IFN-γ, promotes a host defense (M1) macrophage phenotype with bactericidal activity that contributes to pathogen elimination. These cells have been found to produce high levels of nitric oxide and express high levels of CD80 and CD86 [21][22]. Binding of CD40 on the macrophage with CD40L (CD154) on activated T cells is essential for T cell proliferation and induction of pro-inflammatory responses including IL-12 expression and nitric oxide production [23][24][25]. In bovine monocyte-derived macrophages (MDMs) infected with MAP, defects in CD40-CD40L signaling resulted in abrogated gene expression for IL-12p40 and inducible nitric oxide synthase [26]. In contrast, uninfected MDMs were able to upregulate these responses in addition to increased IL-6, TNF-α, IL-8, and IL-10. The subclinical stage of disease is often associated with greater capacity to produce pro-inflammatory cytokines such as IFN-γ and TNF-α [27][28]. In the first year of infection with MAP, it has been shown CD4+CD45RO+ memory T cells are the main antigen responders, and significantly upregulate activation markers CD25 and CD26 following in vitro stimulation [29]. This sentudry also observed that while CD8+CD45RO+ comprised approximately 30% of CD8+ T cells in the first year, they did not begin to have significant responses to in vitro stimulation until 18 months post infection, at which time they increased their expression of activation markers CD25 and CD26 [29]. Importantly, prior activation of the macrophage can have a profound effect on intracellular viability of the invading bacterium, which has been shown in IFN-γ pretreatment of macrophage in vitro [30][31]. Stimulation with a pro-inflammatory mediator after infection, however, does not confer the same protective effects. Furthermore, physiological responses such as polarization and signaling of T cells following antigen presentation rely on a variety of factors including antigen dose, its affinity for the T cell receptor (TCR), duration of binding, and production of anti-inflammatory IL-4 or pro-inflammatory IFN-γ [32][33]. The resolution and repair (M2) macrophage facilitates an environment in which a T helper 2 (Th2) cytokine profile predominates, defined by an anti-inflammatory response with increased IL-10 along with high levels of CD163 [34][35][36]. In the case of bovine paratuberculosis, the greater proportion of M2 macrophages present in the intestinal tissue of cows in clinical stage of disease ultimately hinders MAP clearance and disease resolution within the host [37][38]. This stage is characterized by a reduction in IFN-γ production, a cytokine which is protective for the host and functions by activating macrophages [28]. While the specific mechanisms are unresolved in this transition of immune function from subclinical to clinical cows, it has been shown that CD4+CD25- naïve T cells are largely unresponsive to MAP antigen and do not develop a regulatory T cell cytokine profile [39]. MAP-induced Th1 cytokine gene expression in these CD4+CD25- T cells was observed to be reduced in subclinical cows as well, but to a lesser extent. Furthermore, IL-10 facilitates reduction in IFN-γ and IL-12 and, has also been shown to reduce major histocompatibility complex II (MHCII) on monocytes, collectively resulting in reduced cell activation and antigen presentation [36][40][41]. Signaling through TLR2 has been implicated in facilitating pathogenic mycobacteria’s ability to upregulate IL-10 [36][42]. T cell subsets largely contributing to IL-10 production are CD4+CD25+ regulatory cells, and in the absence of CD25+ cells IFN-γ production is significantly enhanced [43]. Fresh, unstimulated PBMCs from cows with clinical paratuberculosis have been shown to have significantly lower CD25 expression compared to subclinical and control cows but following MPS activation no differences among infection status groups were observed [44]. When T cell subsets were parsed out, MPS stimulated PBMCs from subclinical and clinical cows had significantly higher CD4+CD25+ T cells, and this observation has been replicated [45]. An additional T cell subset found in significant proportions in the bovine is the γδ T cell. This subset can constitute up to 60% of all T cells, and are speculated to bridge responses between innate and adaptive immunity. They can express cytokines such as IL-2, IL-10, IL-12, IL-15, and IFN-γ [46] and have been found to have regulatory function through spontaneous IL-10 secretion which has negative effects on proliferation for CD4+ and CD8+ T cells [47]. In the presence of Mycobacterium bovis (M. bovis) infected dendritic cells, γδ T cells upregulate production of pro-inflammatory IFN-γ and IL-12 [48]. Additionally, γδ T cells have been reported to be significantly lower in cattle with clinical paratuberculosis [44][49]. Furthermore, investigation of γδ T cell distribution in granulomatous tissue in the bovine has shown significantly higher amount of these cells localized to late stage lesions in naïve calves experimentally infected with MAP when compared to vaccinated calves, which also had higher lesion scores [50]. This experiment begs the question of whether γδ T cells are lower in clinical cows due to them exiting the periphery to assist in controlling bacteria at the site of infection. Granulomatous lesions in paratuberculosis are largely unorganized and resemble that of Type II lepromatous granulomas, with disordered structure and macrophages containing high amounts of bacteria [51][52]. Granuloma formation at the site of infection is dependent upon pro-inflammatory TNF-α expression, and blocking TNF-α expression can result in downregulation of IFN-γ, IL-12, IL-10, IL-17, and nitric oxide production [53]. In M. bovis infected cattle, antigen-specific responses by T cells show CD4+ T cells are significant producers of IL-22, while γδ T cells are the main source of IL-17A but can also concurrently produce IL-22 [54]. IL-17A has been associated with early responses in mycobacterial infections [55] and can be produced by CD4+ and CD8+ T cells outside of antigen presenting cell stimulation [56].4. Vitamin D
4.1. History
The story of vitamin D began in the early 1900′s during a time when vitamin deficiencies were more common, and their underlying root cause elusive to physicians. Accessory dietary requirements that were shown to prevent a variety of clinical manifestations led scientists to subscribe to the concept of “vital amines” [57][58][59]. As a result, conventional wisdom of simply balancing dietary proportions of protein, carbohydrates, fats, and salts began to evolve. Soon after the discovery of Vitamins A, B, and C, McCollum et al. discovered that feeding cod liver, oxidized or not, healed rickets [60]. He also made the observation that developing clinical symptoms of rickets in preparation for disease resolution experiments takes significantly longer in the summer than the winter, one of the first hints that sunlight is important in the mechanism. Work done in the 1930′s confirmed this observation, showing ultraviolet rays convert 7-dehydrocholesterol to vitamin D3 in the skin of hogs [61].7.2. Metabolism and Signaling
4.2. Metabolism and Signaling
Two isoforms of vitamin D3 are known currently. Vitamin D2 is converted from ergocalciferol found in plant material and is known to be a less potent regulator of serum 25-hydroxyvitamin D3 (25(OH)D3) concentration in humans [62][63]. Animal derived vitamin D3 originates from 7-dehydrocholesterol and can be absorbed through the diet or converted in the skin through exposure to ultraviolet rays from sunlight. Further steps in activation of 7-dehydrocholsterol occur through multiple hydroxylation reactions. 25(OH)D3, also called previtamin D3, is formed in the liver through action of several cytochrome P450 hydroxylases, of which the most common is thought to be CYP27A1 [64][65]. Specifically in cattle, CYP27A1 and CYP2J2 have been associated with regulating incidence of milk fever, which results from dysregulation of calcium homeostasis [66][67]. Next, 25(OH)D3 is shuttled through the periphery for local conversion by 1α-hydroxylase (CYP27B1) [68]. Activation of 25(OH)D3 by CYP27B1 occurs at a variety of cellular sites, including the kidney [69][70], maternal uterine tissue [71], bone cells [72][73], macrophages [74], and skin cells [75][76]. A visual summary of vitamin D3 metabolism is presented in Figure 1.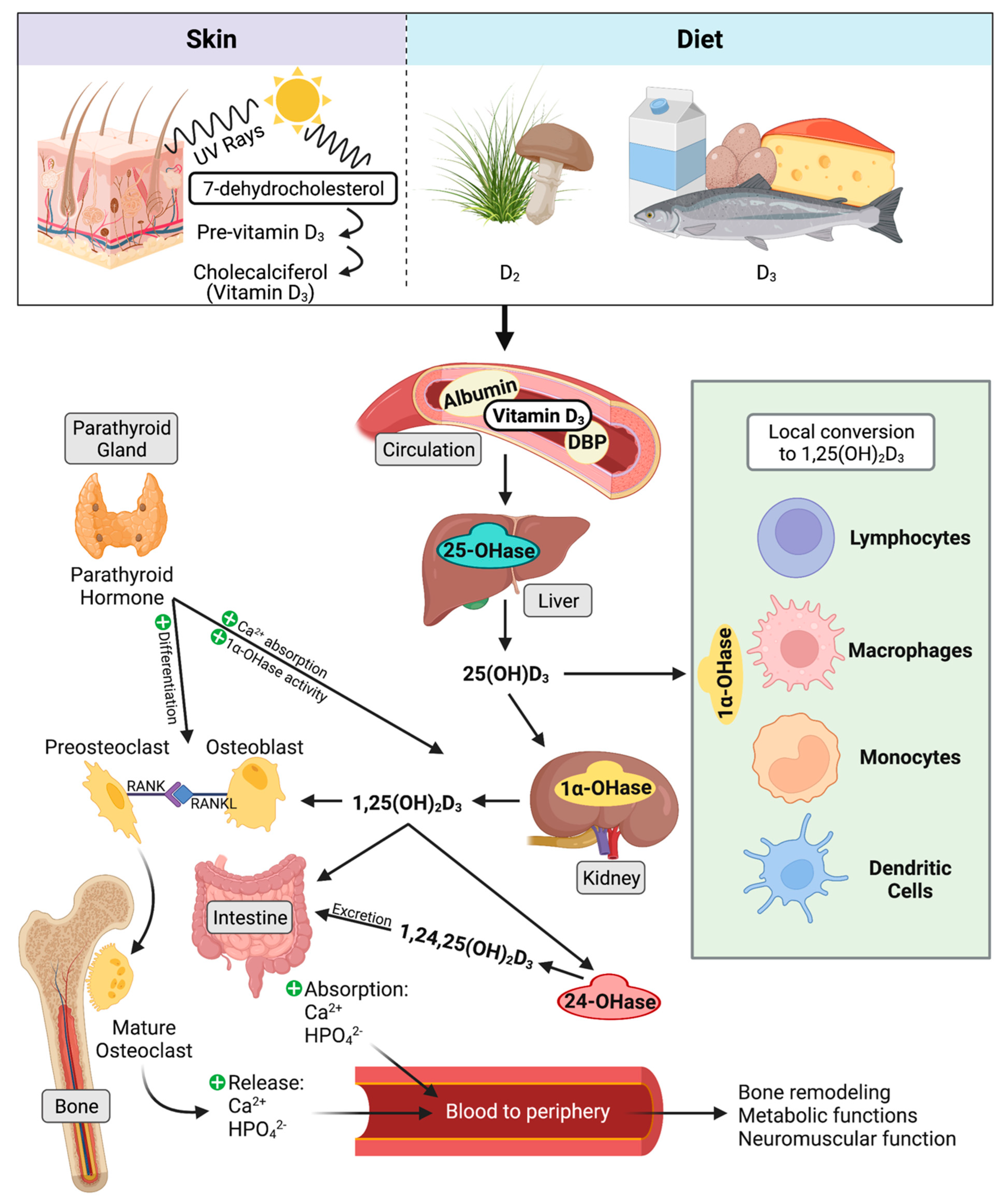
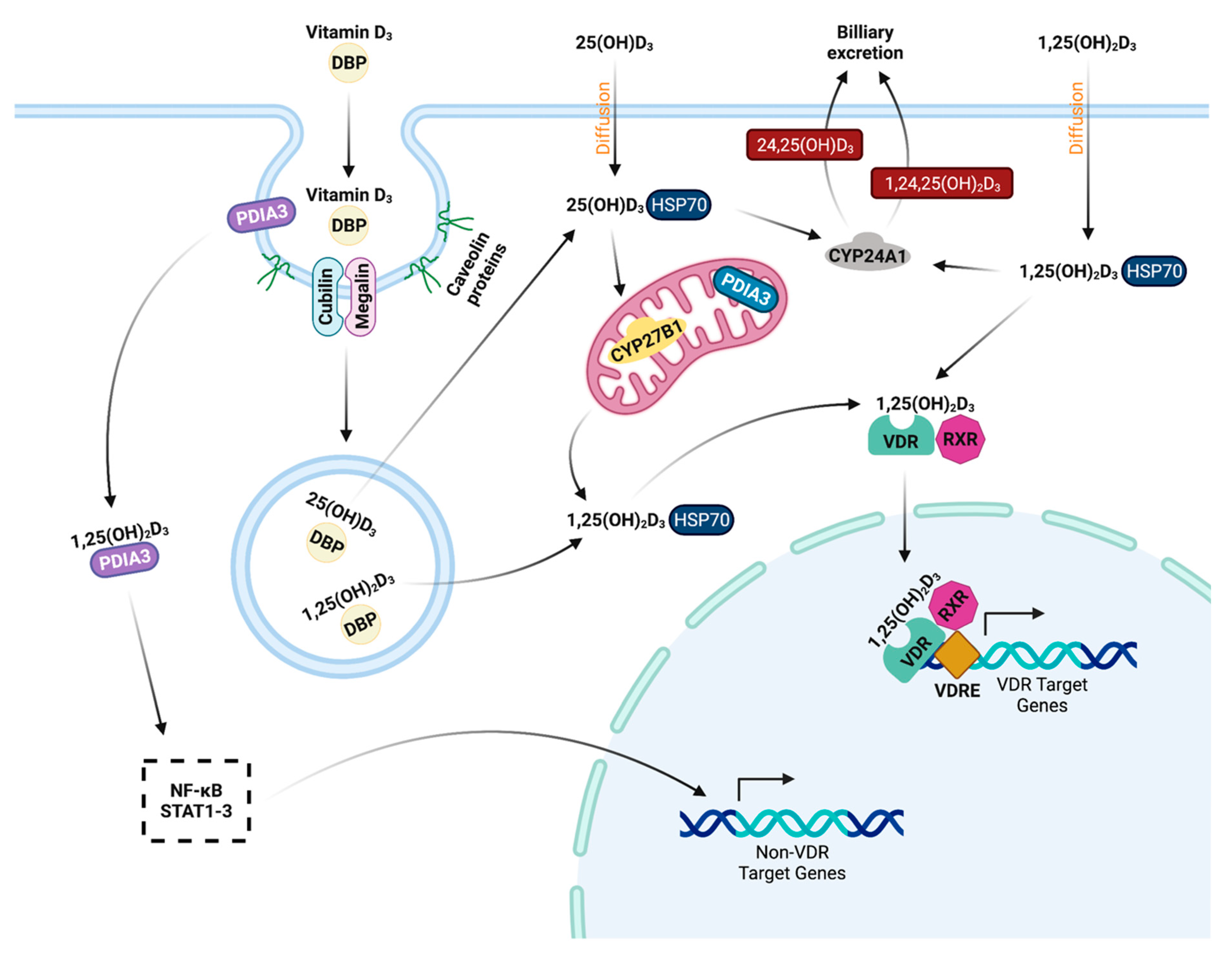
4.3. Classical Function
25(OH)D3 is considered a prohormone, as it shares similar steroid chemical structure as adrenal and sex hormones [101]. When calcium levels are insufficient, bioactive 1,25(OH)2D3 facilitates upregulation of calcium transport mechanisms to increase calcium and phosphate absorption by the intestine and renal tubule cells [67]. 1,25(OH)2D3 facilitates bone growth and remodeling by stimulating differentiation and maturation of osteoblasts and osteoclasts, while remineralization is downregulated by high phosphate and osteopontin levels [102]. Reduced serum calcium concentrations stimulate production of parathyroid hormone from the parathyroid gland. 1,25(OH)2D3 together with parathyroid hormone also facilitates demineralization of bone to release stored calcium by increasing activity of osteoclasts [103]. When sufficient serum calcium levels have been achieved, calcium inhibits production of parathyroid hormone, which then leads to inhibition of 1,25(OH)2D3 synthesis by CYP27B1 in the kidney [104]. While these classical functions of vitamin D3 are generally well understood in several species, much still stands to be elucidated about vitamin D3′s immunomodulatory role, particularly in cattle.4.4. Hydroxylase Expression in Immune Cells
Evidence of 25(OH)D3 conversion to 1,25(OH)2D3 was first shown in macrophages from humans suffering from sarcoidosis [105] and is shown not to be regulated by parathyroid hormone and calcium [106]. Stimulation of 1,25(OH)2D3 production in monocytes and macrophages is a product of cellular activation through pro-inflammatory cytokines such as IFN-γ, TNF-α, and IL-1β [107][108][109]. Activation of peripheral bovine monocytes by lipopolysaccharide (LPS) induces CYP27B1 expression, but the concurrent addition of 1,25(OH)2D3 to cell cultures interrupts expression, bringing it back closer to baseline [110]. Gene expression for both CYP27B1 and CYP24A1 have been shown to be reduced by 1,25(OH)2D3 in bovine PBMCs and MDMs activated by MAP sonicate or live MAP [45][111]. The reduction in vitamin D inactivating hydroxylase in this model may highlight a mechanism that enables monocytes and macrophages to maintain 1,25(OH)2D3 levels during infection. Cattle with clinical stage paratuberculosis experience abrogated expression of CYP27B1 in the ileum, which may be a feature of inadequate access to its substrate 25(OH)D3 as a result of significantly reduced circulating levels in these animals. Interestingly, increased levels of IFN-γ have been associated with upregulation of CYP27B1 activity [112][113]. Additionally, a Streptococcus uberis (S. uberis) bovine mastitis model observed localized expression increased for CYP27B1 in milk CD14+ cells during active mastitis, while CD14- cells saw increased CYP24A1 [114]. Bovine peripheral blood monocytes treated with 1,25(OH)2D3 show increased CYP24A1 transcripts, which are then reduced following activation with LPS [115]. Similar observations have been made in M. bovis-BCG vaccinated calves, where secondary purified protein derivative (PPD) antigen exposure abrogated the 1,25(OH)2D3 induced upregulation of CYP24A1 [116]. Intramammary treatment of healthy cows with 1,25(OH)2D3 upregulates expression of CYP24A1 in total milk somatic cells as early as 4 h post treatment [117], as well as increases CYP24A1 expression in milk macrophages and neutrophils [115]. Furthermore, the S. uberis infected mammary gland overall expressed increased VDR as well [114], however VDR expression does not undergo changes in the bovine ileum from MAP infected cows [113][118]. These studies show vitamin D signaling pathways and metabolism may be differentially mediated depending on the offending pathogen.4.5. Host Vitamin D Status and Cathelicidins
Minimum thresholds of 25(OH)D3 concentrations reflecting deficiency, insufficiency, and therapeutic immune function have not been concretely established for neither humans nor cattle. An estimation for minimum levels required for immune function is speculated to be 30 ng/mL, whereas baseline concentrations for proper calcium signaling mechanisms is 20 ng/mL [119][120][121][122]. As previously mentioned in the case of bovine paratuberculosis, circulating levels of 25(OH)D3 have been shown to be significantly reduced in animals with late stage clinical disease [113]. Work in cattle has shown concentrations of serum 25(OH)D3 in adult dairy cows from across the United States at various stages of lactation and supplemented with 30,000–50,000 IU of vitamin D3 per day vastly fall between 40–100 ng/mL, with an average of 68 ng/mL [123]. Additionally, seasonality has been shown to impact 25(OH)D3 status, with serum concentrations being higher in the summer [124]. This sentudry also reported a high incidence of 25(OH)D3 deficiency in calves following birth, observing concentrations mainly between 3–17 ng/mL and over 80% having lower than 20 ng/mL. These data highlight the importance of vitamin D3 supplementation beginning from birth, especially during a time of greatest susceptibility to MAP infection. Efficiency of vitamin D-induced antimicrobial activity is determined by the vitamin D status of the host, as availability of 25(OH)D3 substrate is ultimately a limiting factor. Evidence for this observation is reported in human monocytes and macrophages activated by M. tb or LPS signaling via TLR2/1 heterodimer or TLR4 binding, where serum with low levels of 25(OH)D3 had reduced ability to induce cathelicidin expression [125][126]. Cathelicidins are small cationic peptides that can upregulate expression of chemokines by macrophages, along with increasing phagocytosis of bacteria [127][128]. One functional cathelicidin gene has been identified in humans and mice, while 7 out of 11 cathelicidin genes in cattle have been shown to produce active protein [129][130]. Three of the bovine genes have purported vitamin D response elements, but studies thus far have shown their expression is not induced by 1,25(OH)2D3 [110]. Other immunologic species differences exist. Cattle are estimated to have over 100 defensin genes while human and mice are estimated to have 39 and 52, respectively [131][132].4.6. Macrophage Phagocytosis and Phenotype
Enhanced capacity for phagocytosis has been observed to be induced in monocytes and macrophages following treatment with 1,25(OH)2D3 [111][133][134]. In another study, phagocytosis was upregulated in macrophages from healthy individuals that had a low phagocytic index, but those with a current pulmonary tuberculosis infection did not see a benefit [135]. This study used cultured PBMCs in the presence of 1,25(OH)2D3 for 48 h followed by infection with M. tb for 3 h, so in considering the short half-life of 1,25(OH)2D3 it is possible any 1,25(OH)2D3-induced effects were not captured for this cell culture method. Small et al. showed increased phagocytosis of Staphylococcus aureus and Candida albicans with a concurrent upregulation of complement receptor immunoglobulin (CRIg) in human macrophages treated with 1,25(OH)2D3 [136]. This study utilized complement opsonized microbes; however, CRIg can function as an innate macrophage pattern recognition receptor (PRR) and directly bind some Gram-positive bacteria through recognition of lipoteichoic acids [137]. This posits the question of whether CRIg can also directly recognize any cell wall constituents in mycobacterial species; however, more definitive studies need to be performed to validate the PRR functions of CRIg outside of recognizing opsonized particles. A study using human monocytes differentiated to macrophages by pro-inflammatory IL-15 showed antimicrobial effects against intracellular Mycobacterium leprae (M. leprae) were induced by the presence of 25(OH)D3 during differentiation [138]. Compared to IL-10 differentiated macrophages, IL-15 differentiated macrophages expressed lower amounts of CD163 and higher amounts of CD209, possibly indicating a more M2-like phenotype. This study did not observe any changes in phagocytosis related to 25(OH)D3 treatment, further indicating a M1-like phenotype, as M2 macrophages have been shown to have better phagocytic capabilities [139]. Macrophage phenotype markers CD163 (M1) and CD80 (M2) have recently been shown to have vitamin D3-induced changes in expression in bovine MDMs. When infected in vitro with MAP, MDMs from cows with subclinical and clinical stage paratuberculosis exhibit reduced CD80 expression following treatment with 1,25(OH)2D3 [140]. In control cow MDMs infected in vitro with MAP, both 1,25(OH)2D3 and 25(OH)D3 reduced CD80. CD163 was largely upregulated by 1,25(OH)2D3 treatment regardless of activation with MAP. An exception was observed in subclinical cows, which showed 1,25(OH)2D3 and 25(OH)D3 reduced CD163 expression [140]. To fully understand changes elicited by vitamin D3, further work including a more expansive macrophage phenotype panel is needed.4.7. Cytokines, Nitric Oxide, and β-Defensins
Modulation of immune responses by exogenous vitamin D3 during infectious disease has shown positive outcomes following treatment. PBMCs from M. bovis infected cattle treated in vitro with 1,25(OH)2D3 show antigen-specific recall responses through increased nitric oxide production, although pro-inflammatory IFN-γ production was inhibited [141]. A similar reduction in IFN-γ expression has been observed in stimulated PBMCs treated with 1,25(OH)2D3 from M. bovis vaccinated cattle [142]. Nelson et al. also showed a concurrent reduction in gene expression after vitamin D3 treatment for IFN-γ, IL-17A, and IL-17F, although only significant for IL-17F during 1,25(OH)2D3 treatment [116]. Contrasting previous reports, 1,25(OH)2D3 treatment and activation with MAP has shown to induce a significant increase in IFNG transcripts and a concurrent reduction in IFN-γ secretion in PBMC-MDM co-cultures from cattle with naturally acquired paratuberculosis [111]. Similar observations have been reported in PBMCs, with these disparate effects being highlighted in cows in the clinical stage of Johne’s disease [45]. This may highlight a key defense mechanism employed by MAP; however, in another perspective, by reducing IFN-γ production the host may be protected from unnecessary tissue damage while it employs other antimicrobial mechanisms. Activation of monocytes from healthy cattle by LPS upregulates pro-inflammatory IL-1β (IL1B), but the effect is independent of 1,25(OH)2D3 exposure [110]. In MAP activated PBMC-MDM co-cultures, 1,25(OH)2D3 significantly upregulates production of IL1B transcripts and IL-1β, with an accompanying reduction in IL10 and IL-10 [111]. However, when PBMCs were cultured alone and activated with MAP sonicate, 1,25(OH)2D3 increased IL10 transcripts but reduced IL-10 secretion in cows with paratuberculosis [45]. Similarly activated PBMCs from cows with clinical paratuberculosis observed reduced IL10 and IL-10 following 25(OH)D3 treatment [45]. These data could indicate that vitamin D3-induced effects are more efficiently deployed when PBMCs, especially T cells, have the opportunity for crosstalk with macrophages. In addition to the previously discussed cathelicidin induction, human tuberculosis patients also observe increased nitric oxide production in alveolar macrophages following treatment with vitamin D3, but this mechanism is not thought to be regulated by TLR signaling [143]. Human promyelocytic cell line HL-60 has also reported upregulation of nitric oxide production, NOS2 expression, and a resulting inhibition of intracellular M. tb growth following treatment with 1,25(OH)2D3. 1,25(OH)2D3 also reduces levels of intracellular M. tb in human monocytes and macrophages [144][145]. To further highlight the critical role of vitamin D3 in infectious disease, NOS2 knockout mice infected with M. bovis experience increased capacity to kill the bacteria if they have sufficient circulating 25(OH)D3 [146]. Work in cattle has shown that dietary supplementation with 25(OH)D3 results in a greater proportion of peripheral blood neutrophils with antimicrobial activity, as measured by oxidative burst [147]. 1,25(OH)2D3 has recently been shown to induce nitric oxide production along with upregulating NOS2 expression in macrophages from healthy cattle that have underwent in vitro infection with M. bovis [133]. In contrast, MAP activated PBMCs and PBMC-MDM co-cultures from dairy cattle with naturally acquired paratuberculosis experience reduced NOS2 expression following 1,25(OH)2D3 treatment [45][111]. Curiously, 1,25(OH)2D3 significantly upregulated nitrite production, measured as an indicator of iNOS activity, in the activated PBMC-MDM co-cultures from cows with subclinical and clinical paratuberculosis [111]. At this 24 h timepoint, live MAP may target 1,25(OH)2D3-induced signaling events to disrupt availability of transcripts for protein translation. In bovine PBMCs activated with MAP sonicate, CCL5 expression has been shown to be reduced by both 1,25(OH)2D3 or 25(OH)D3 [45]. Other work in peripheral bovine monocytes from healthy dairy cattle has shown upregulated gene expression at 24 h for inducible nitric oxide synthase (iNOS/NOS2) and RANTES/CCL5 in bovine monocytes coordinated by 1,25(OH)2D3, an effect that is greatly enhanced by concurrent LPS activation [110]. This sentudry also showed CCL5, NOS2, and nitrite production increase in a dose-dependent manner for 1,25(OH)2D3 [110]. In a model utilizing PBMCs from calves vaccinated with M. bovis-BCG, secondary exposure to M. bovis PPD and in vitro treatment with 1,25(OH)2D3 or 25(OH)D3 resulted in RANTES/CCL5 gene expression upregulation [116]. A recent report has similarly shown induction of NOS2 by 1,25(OH)2D3 following LPS activation and increased nitrite production [148]. Peripheral blood monocytes from healthy cows treated in vitro with 1,25(OH)2D3 increase expression of various β-defensins, including DEFB3, DEFB6, DEFB7, DEFB10, along with NOS2, with the effect enhanced further in LPS stimulated cells [115]. Opposite observations were seen in DEFB5, whose expression was reduced, and the effect enhanced by LPS. This experiment used an 18 h timepoint, and a follow up experiment using a 4 h timepoint shows peripheral blood monocytes upregulated all bovine β-defensin genes investigated following treatment with 1,25(OH)2D3, indicating these cellular responses are more robust early in antigen exposure. Recently, studies have shown PBMCs from cattle at different stages of paratuberculosis infection do not have significantly different levels of DEFB4 and DEFB7 expression, and there were no notable vitamin D3 induced effects [45]. Infection status effects on expression of these β-defensin genes were also not present in co-cultures of PBMCs and MDMs; however, in this model 1,25(OH)2D3 was shown to significantly reduce DEFB7 in control and subclinical cows. In milk neutrophils, LPS activation alone at 18 h increased transcripts of DEFB3, DEFB4, DEFB7, and DEFB10. The authors further show that 1,25(OH)2D3 treatment of the mammary gland facilitates upregulation of DEFB7 at 8 h in milk macrophages [115]. Follow up studies by Merriman et al. show direct treatment of mammary gland with 1,25(OH)2D3 in healthy cows upregulated expression of NOS2 and DEFB7 in total milk somatic cells at 4 h following treatment [117]. When cows with subclinical mastitis had their mammary gland treated with 1,25(OH)2D3, upregulation was enhanced further and increased expression was observed for DEFB4 and DEFB7 along with NOS2, with a significant treatment effect being seen by 24 h [117]. 1,25(OH)2D3 induces localized expression of NOS2 in milk CD14+ cells and increased CCL5 in CD14- cells during experimentally induced mastitis caused by S. uberis [114]. In a similar study, treatment of mammary gland with 25(OH)D3 in cattle with LPS induced mastitis showed total milk somatic cells having upregulated expression of NOS2, RANTES/CCL5, DEFB3, DEFB4, DEFB7, DEFB10, IL1B, and IL8, with most effects being observed between 4–8 h following treatment [149]. Further isolating the source of these responses, the authors showed milk macrophages expressing significantly greater amounts of NOS2 transcripts when compared to untreated controls, but the significant upregulation of other responses were sourced from neutrophils [149]. Another S. uberis induced model of mastitis has shown a resulting decrease in bacterial load in the mammary gland, along with clinical symptoms, after directly treating the gland with 25(OH)D3 following each milking [150]. Collectively, these studies may indicate that the type of pathogen and its preferred tissue may heavily influence antimicrobial responses facilitated by vitamin D3.4.8. Macrophage Endosomal Trafficking
The desire to understand mechanisms of intracellular MAP survival have driven recent studies to investigate the relationship between vitamin D3 and endosomal trafficking markers at different stages of Johne’s disease. 1,25(OH)2D3 and 25(OH)D3 have no significant effects on early endosomal marker Rab5 expression in bovine MDMs; however, control cow MDMs infected with live MAP and treated with either form of vitamin D3 experienced significantly reduced Rab5 expression [140]. A notable observation was made in MDMs from cows with subclinical and clinical paratuberculosis, which had significantly reduced Rab5 expression upon in vitro MAP infection when compared to MDMs from control cows. Late endosomal marker Rab7 expression was consistently reduced by 1,25(OH)2D3 treatment in MDMs from control and subclinical cows, regardless of in vitro MAP infection [140]. The lack of downregulatory effect in clinical cows posits the question if 1,25(OH)2D3 treatment has therapeutic potential for these animals, especially considering previous reports of animals in this severe stage of disease having reduced circulating 25(OH)D3. Expression of endosomal markers in ileal macrophages from cows in different stages of paratuberculosis have also been recently investigated. Rab5 expression in clinical cow macrophages was shown to be significantly reduced compared to other groups [118]. Total macrophage Rab7 expression was not different among groups, but when colocalized with intracellular MAP subclinical cows showed no detectable association [118]. This may indicate MAP employs virulence mechanisms during early infection to inhibit recruitment of Rab7 to the intracellular compartment it is contained in, which would provide opportunity for MAP’s replication and facilitation of the chronic subclinical phase. Further studies are needed to fully elucidate MAP’s ability to interrupt the phagosomal maturation pathway and the role vitamin D3 plays.References
- Buergelt, C.D.; Hall, C.; McEntee, K.; Duncan, J.R. Pathological Evaluation of Paratuberculosis in Naturally Infected Cattle. Vet. Pathol. 1978, 15, 196–207.
- Ott, S.L.; Wells, S.J.; Wagner, B.A. Herd-Level Economic Losses Associated with Johne’s Disease on US Dairy Operations. Prev. Vet. Med. 1999, 40, 179–192.
- Rasmussen, P.; Barkema, H.W.; Mason, S.; Beaulieu, E.; Hall, D.C. Economic Losses Due to Johne’s Disease (Paratuberculosis) in Dairy Cattle. J. Dairy Sci. 2021, 104, 3123–3143.
- Batt, S.M.; Minnikin, D.E.; Besra, G.S. The Thick Waxy Coat of Mycobacteria, a Protective Layer against Antibiotics and the Host’s Immune System. Biochem. J. 2020, 477, 1983–2006.
- Gao, L.-Y.; Laval, F.; Lawson, E.H.; Groger, R.K.; Woodruff, A.; Morisaki, J.H.; Cox, J.S.; Daffe, M.; Brown, E.J. Requirement for kasB in Mycobacterium Mycolic Acid Biosynthesis, Cell Wall Impermeability and Intracellular Survival: Implications for Therapy. Mol. Microbiol. 2003, 49, 1547–1563.
- Salgado, M.; Collins, M.T.; Salazar, F.; Kruze, J.; Bölske, G.; Söderlund, R.; Juste, R.; Sevilla, I.A.; Biet, F.; Troncoso, F.; et al. Fate of Mycobacterium avium Subsp. paratuberculosis after Application of Contaminated Dairy Cattle Manure to Agricultural Soils. Appl. Environ. Microbiol. 2011, 77, 2122–2129.
- Whittington, R.J.; Marshall, D.J.; Nicholls, P.J.; Marsh, I.B.; Reddacliff, L.A. Survival and Dormancy of Mycobacterium avium Subsp. paratuberculosis in the Environment. Appl. Environ. Microbiol. 2004, 70, 2989–3004.
- Whittington, R.J.; Marsh, I.B.; Reddacliff, L.A. Survival of Mycobacterium avium Subsp. paratuberculosis in Dam Water and Sediment. Appl. Environ. Microbiol. 2005, 71, 5304–5308.
- Chiodini, R.J.; Van Kruiningen, H.J. Eastern White-Tailed Deer as a Reservoir of Ruminant Paratuberculosis. J. Am. Vet. Med. Assoc. 1983, 182, 168–169.
- Reyes-García, R.; Pérez-de-la-Lastra, J.M.; Vicente, J.; Ruiz-Fons, F.; Garrido, J.M.; Gortázar, C. Large-Scale ELISA Testing of Spanish Red Deer for Paratuberculosis. Vet. Immunol. Immunopathol. 2008, 124, 75–81.
- Mortier, R.A.R.; Barkema, H.W.; Bystrom, J.M.; Illanes, O.; Orsel, K.; Wolf, R.; Atkins, G.; De Buck, J. Evaluation of Age-Dependent Susceptibility in Calves Infected with Two Doses of Mycobacterium avium Subspecies paratuberculosis Using Pathology and Tissue Culture. Vet. Res. 2013, 44, 94.
- Windsor, P.A.; Whittington, R.J. Evidence for Age Susceptibility of Cattle to Johne’s Disease. Vet. J. 2010, 184, 37–44.
- Sweeney, R.W.; Whitlock, R.H.; Rosenberger, A.E. Mycobacterium paratuberculosis Cultured from Milk and Supramammary Lymph Nodes of Infected Asymptomatic Cows. J. Clin. Microbiol. 1992, 30, 166–171.
- Sweeney, R.W.; Whitlock, R.H.; Rosenberger, A.E. Mycobacterium paratuberculosis Isolated from Fetuses of Infected Cows Not Manifesting Signs of the Disease. Am. J. Vet. Res. 1992, 53, 477–480.
- Whittington, R.J.; Windsor, P.A. In Utero Infection of Cattle with Mycobacterium avium Subsp. paratuberculosis: A Critical Review and Meta-Analysis. Vet. J. 2009, 179, 60–69.
- Mitchell, R.M.; Schukken, Y.; Koets, A.; Weber, M.; Bakker, D.; Stabel, J.R.; Whitlock, R.H.; Louzoun, Y. Differences in Intermittent and Continuous Fecal Shedding Patterns between Natural and Experimental Mycobacterium avium Subspecies paratuberculosis Infections in Cattle. Vet. Res. 2015, 46, 66.
- Schukken, Y.H.; Whitlock, R.H.; Wolfgang, D.; Grohn, Y.; Beaver, A.; VanKessel, J.; Zurakowski, M.; Mitchell, R. Longitudinal Data Collection of Mycobacterium avium Subspecies paratuberculosis Infections in Dairy Herds: The Value of Precise Field Data. Vet. Res. 2015, 46, 65.
- Whitlock, R.H.; Buergelt, C. Preclinical and Clinical Manifestations of Paratuberculosis (Including Pathology). Vet. Clin. N. Am. Food Anim. Pract. 1996, 12, 345–356.
- Sweeney, R.W. Pathogenesis of Paratuberculosis. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 537–546.
- Martinez, F.O.; Gordon, S. The M1 and M2 Paradigm of Macrophage Activation: Time for Reassessment. F1000Prime Rep. 2014, 6, 13.
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686.
- Ambarus, C.A.; Krausz, S.; van Eijk, M.; Hamann, J.; Radstake, T.R.D.J.; Reedquist, K.A.; Tak, P.P.; Baeten, D.L.P. Systematic Validation of Specific Phenotypic Markers for in Vitro Polarized Human Macrophages. J. Immunol. Methods 2012, 375, 196–206.
- Grewal, I.S.; Flavell, R.A. A Central Role of CD40 Ligand in the Regulation of CD4+ T-Cell Responses. Immunol. Today 1996, 17, 410–414.
- Grewal, I.S.; Flavell, R.A. CD40 and CD154 in Cell-Mediated Immunity. Annu. Rev. Immunol. 1998, 16, 111–135.
- Kato, T.; Hakamada, R.; Yamane, H.; Nariuchi, H. Induction of IL-12 P40 Messenger RNA Expression and IL-12 Production of Macrophages via CD40-CD40 Ligand Interaction. J. Immunol. 1996, 156, 3932–3938.
- Sommer, S.; Pudrith, C.B.; Colvin, C.J.; Coussens, P.M. Mycobacterium avium Subspecies paratuberculosis Suppresses Expression of IL-12p40 and iNOS Genes Induced by Signalling through CD40 in Bovine Monocyte-Derived Macrophages. Vet. Immunol. Immunopathol. 2009, 128, 44–52.
- Stabel, J.R. Production of γ-Interferon by Peripheral Blood Mononuclear Cells: An Important Diagnostic Tool for Detection of Subclinical Paratuberculosis. J. Vet. Diagn. Investig. 1996, 8, 345–350.
- Stabel, J.R. Cytokine Secretion by Peripheral Blood Mononuclear Cells from Cows Infected with Mycobacterium paratuberculosis. Am. J. Vet. Res. 2000, 61, 754–760.
- Koo, H.C.; Park, Y.H.; Hamilton, M.J.; Barrington, G.M.; Davies, C.J.; Kim, J.B.; Dahl, J.L.; Waters, W.R.; Davis, W.C. Analysis of the Immune Response to Mycobacterium avium Subsp. paratuberculosis in Experimentally Infected Calves. Infect. Immun. 2004, 72, 6870–6883.
- Bonecini-Almeida, M.G.; Chitale, S.; Boutsikakis, I.; Geng, J.; Doo, H.; He, S.; Ho, J.L. Induction of In Vitro Human Macrophage Anti-Mycobacterium tuberculosis Activity: Requirement for IFN-γ and Primed Lymphocytes. J. Immunol. 1998, 160, 4490–4499.
- Hostetter, J.M.; Steadham, E.M.; Haynes, J.S.; Bailey, T.B.; Cheville, N.F. Cytokine Effects on Maturation of the Phagosomes Containing Mycobacteria avium Subspecies paratuberculosis in J774 Cells. FEMS Immunol. Med. Microbiol. 2002, 34, 127–134.
- Lee, B.O.; Haynes, L.; Eaton, S.M.; Swain, S.L.; Randall, T.D. The Biological Outcome of CD40 Signaling Is Dependent on the Duration of CD40 Ligand Expression: Reciprocal Regulation by Interleukin (IL)-4 and IL-12. J. Exp. Med. 2002, 196, 693–704.
- Rogers, P.R.; Croft, M. Peptide Dose, Affinity, and Time of Differentiation Can Contribute to the Th1/Th2 Cytokine Balance. J. Immunol. 1999, 163, 1205–1213.
- Buechler, C.; Ritter, M.; Orsó, E.; Langmann, T.; Klucken, J.; Schmitz, G. Regulation of Scavenger Receptor CD163 Expression in Human Monocytes and Macrophages by Pro- and Antiinflammatory Stimuli. J. Leukoc. Biol. 2000, 67, 97–103.
- Khare, S.; Drake, K.L.; Lawhon, S.D.; Nunes, J.E.S.; Figueiredo, J.F.; Rossetti, C.A.; Gull, T.; Everts, R.E.; Lewin, H.A.; Adams, L.G. Systems Analysis of Early Host Gene Expression Provides Clues for Transient Mycobacterium avium Ssp avium vs. Persistent Mycobacterium avium Ssp paratuberculosis Intestinal Infections. PLoS ONE 2016, 11, e0161946.
- Hussain, T.; Shah, S.Z.A.; Zhao, D.; Sreevatsan, S.; Zhou, X. The Role of IL-10 in Mycobacterium avium Subsp. paratuberculosis. Cell Commun. Signal. 2016, 14, 29.
- Khalifeh, M.S.; Stabel, J.R. Effects of Gamma Interferon, Interleukin-10, and Transforming Growth Factor β on the Survival of Mycobacterium avium Subsp. paratuberculosis in Monocyte-Derived Macrophages from Naturally Infected Cattle. Infect. Immun. 2004, 72, 1974–1982.
- Jenvey, C.J.; Shircliff, A.L.; Bannantine, J.P.; Stabel, J.R. Phenotypes of Macrophages Present in the Intestine Are Impacted by Stage of Disease in Cattle Naturally Infected with Mycobacterium avium Subsp. paratuberculosis. PLoS ONE 2019, 14, e0217649.
- Roussey, J.A.; Steibel, J.P.; Coussens, P.M. Regulatory T Cell Activity and Signs of T Cell Unresponsiveness in Bovine Paratuberculosis. Front. Vet. Sci. 2014, 1, 20.
- Haddad, J.J.; Saadé, N.E.; Safieh-Garabedian, B. Interleukin-10 and the Regulation of Mitogen-Activated Protein Kinases: Are These Signalling Modules Targets for the Anti-Inflammatory Action of This Cytokine? Cell. Signal. 2003, 15, 255–267.
- Weiss, D.J.; Souza, C.D. Review Paper: Modulation of Mononuclear Phagocyte Function by Mycobacterium avium Subsp. paratuberculosis. Vet. Pathol. 2008, 45, 829–841.
- Weiss, D.J.; Souza, C.D.; Evanson, O.A.; Sanders, M.; Rutherford, M. Bovine Monocyte TLR2 Receptors Differentially Regulate the Intracellular Fate of Mycobacterium avium Subsp. paratuberculosis and Mycobacterium avium Subsp. avium. J. Leukoc. Biol. 2008, 83, 48–55.
- de Almeida, D.E.; Colvin, C.J.; Coussens, P.M. Antigen-Specific Regulatory T Cells in Bovine Paratuberculosis. Vet. Immunol. Immunopathol. 2008, 125, 234–245.
- Stabel, J.R.; Bannantine, J.P. Divergent Antigen-Specific Cellular Immune Responses during Asymptomatic Subclinical and Clinical States of Disease in Cows Naturally Infected with Mycobacterium avium Subsp. paratuberculosis. Infect. Immun. 2019, 88, e00650-19.
- Wherry, T.L.T.; Mooyottu, S.; Stabel, J.R. Effects of 1,25-Dihydroxyvitamin D3 and 25-Hydroxyvitamin D3 on PBMCs from Dairy Cattle Naturally Infected with Mycobacterium avium Subsp. paratuberculosis. Front. Vet. Sci. 2022, 9, 830144.
- Alvarez, A.J.; Endsley, J.J.; Werling, D.; Mark Estes, D. WC1 + γδ T Cells Indirectly Regulate Chemokine Production During Mycobacterium bovis Infection in SCID-Bo Mice. Transbound. Emerg. Dis. 2009, 56, 275–284.
- Guzman, E.; Hope, J.; Taylor, G.; Smith, A.L.; Cubillos-Zapata, C.; Charleston, B. Bovine γδ T Cells Are a Major Regulatory T Cell Subset. J. Immunol. Baltim. Md 1950 2014, 193, 208–222.
- Price, S.J.; Hope, J.C. Enhanced Secretion of Interferon-γ by Bovine γδ T Cells Induced by Coculture with Mycobacterium bovis-Infected Dendritic Cells: Evidence for Reciprocal Activating Signals. Immunology 2009, 126, 201–208.
- Albarrak, S.M.; Waters, W.R.; Stabel, J.R.; Hostetter, J.M. WC1+ γδ T Cells from Cattle Naturally Infected with Mycobacterium avium Subsp. paratuberculosis Respond Differentially to Stimulation with PPD-J. Vet. Immunol. Immunopathol. 2017, 190, 57–64.
- Plattner, B.L.; Doyle, R.T.; Hostetter, J.M. Gamma-Delta T Cell Subsets Are Differentially Associated with Granuloma Development and Organization in a Bovine Model of Mycobacterial Disease. Int. J. Exp. Pathol. 2009, 90, 587–597.
- Koets, A.P.; Eda, S.; Sreevatsan, S. The within Host Dynamics of Mycobacterium avium Ssp. paratuberculosis Infection in Cattle: Where Time and Place Matter. Vet. Res. 2015, 46, 61.
- Lei, L.; Plattner, B.L.; Hostetter, J.M. Live Mycobacterium avium Subsp. paratuberculosis and a Killed-Bacterium Vaccine Induce Distinct Subcutaneous Granulomas, with Unique Cellular and Cytokine Profiles. Clin. Vaccine Immunol. 2008, 15, 783–793.
- Da Silva, D.A.A.; da Silva, M.V.; Barros, C.C.O.; Alexandre, P.B.D.; Timóteo, R.P.; Catarino, J.S.; Sales-Campos, H.; Machado, J.R.; Rodrigues, D.B.R.; Oliveira, C.J.; et al. TNF-α Blockade Impairs in Vitro Tuberculous Granuloma Formation and down Modulate Th1, Th17 and Treg Cytokines. PLoS ONE 2018, 13, e0194430.
- Steinbach, S.; Vordermeier, H.M.; Jones, G.J. CD4+ and γδ T Cells Are the Main Producers of IL-22 and IL-17A in Lymphocytes from Mycobacterium bovis-Infected Cattle. Sci. Rep. 2016, 6, 29990.
- Khader, S.A.; Guglani, L.; Rangel-Moreno, J.; Gopal, R.; Junecko, B.A.F.; Fountain, J.J.; Martino, C.; Pearl, J.E.; Tighe, M.; Lin, Y.; et al. IL-23 Is Required for Long-Term Control of Mycobacterium tuberculosis and B Cell Follicle Formation in the Infected Lung. J. Immunol. 2011, 187, 5402–5407.
- DeKuiper, J.L.; Cooperider, H.E.; Lubben, N.; Ancel, C.M.; Coussens, P.M. Mycobacterium avium Subspecies paratuberculosis Drives an Innate Th17-like T Cell Response Regardless of the Presence of Antigen-Presenting Cells. Front. Vet. Sci. 2020, 7, 108.
- Funk, C. On the Chemical Nature of the Substance Which Cures Polyneuritis in Birds Induced by a Diet of Polished Rice. J. Physiol. 1911, 43, 395–400.
- Holst, A.; Frölich, T. Experimental Studies Relating to “Ship-Beri-Beri” and Scurvy. J. Hyg. 1907, 7, 634–671.
- Hopkins, F.G. Feeding Experiments Illustrating the Importance of Accessory Factors in Normal Dietaries. J. Physiol. 1912, 44, 425–460.
- McCollum, E.V.; Simmonds, N.; Becker, J.E.; Shipley, P.G. Studies on Experimental Rickets: XXI. an Experimental Demonstration of the Existence of a Vitamin Which Promotes Calcium Deposition. J. Biol. Chem. 1922, 53, 293–312.
- Windaus, A.; Bock, F. Über Das Provitamin Aus Dem Sterin Der Schweineschwarte. Z. Physiol. Chem. 1937, 245, 168–170.
- Armas, L.A.G.; Hollis, B.W.; Heaney, R.P. Vitamin D2 Is Much Less Effective than Vitamin D3 in Humans. J. Clin. Endocrinol. Metab. 2004, 89, 5387–5391.
- Tripkovic, L.; Lambert, H.; Hart, K.; Smith, C.P.; Bucca, G.; Penson, S.; Chope, G.; Hyppönen, E.; Berry, J.; Vieth, R.; et al. Comparison of Vitamin D2 and Vitamin D3 Supplementation in Raising Serum 25-Hydroxyvitamin D Status: A Systematic Review and Meta-Analysis. Am. J. Clin. Nutr. 2012, 95, 1357–1364.
- Ponchon, G.; Kennan, A.L.; DeLuca, H.F. “Activation” of Vitamin D by the Liver. J. Clin. Investig. 1969, 48, 2032–2037.
- Prosser, D.E.; Jones, G. Enzymes Involved in the Activation and Inactivation of Vitamin D. Trends Biochem. Sci. 2004, 29, 664–673.
- Pacheco, H.A.; da Silva, S.; Sigdel, A.; Mak, C.K.; Galvão, K.N.; Texeira, R.A.; Dias, L.T.; Peñagaricano, F. Gene Mapping and Gene-Set Analysis for Milk Fever Incidence in Holstein Dairy Cattle. Front. Genet. 2018, 9, 465.
- Reinhardt, T.A.; Horst, R.L.; Goff, J.P. Calcium, Phosphorus, and Magnesium Homeostasis in Ruminants. Vet. Clin. N. Am. Food Anim. Pract. 1988, 4, 331–350.
- Takeyama, K.; Kitanaka, S.; Sato, T.; Kobori, M.; Yanagisawa, J.; Kato, S. 25-Hydroxyvitamin D3 1α-Hydroxylase and Vitamin D Synthesis. Science 1997, 277, 1827–1830.
- Fraser, D.R.; Kodicek, E. Unique Biosynthesis by Kidney of a Biologically Active Vitamin D Metabolite. Nature 1970, 228, 764–766.
- Paulson, S.K.; DeLuca, H.F. Subcellular Location and Properties of Rat Renal 25-Hydroxyvitamin D3-1α-Hydroxylase. J. Biol. Chem. 1985, 260, 11488–11492.
- Weisman, Y.; Harell, A.; Edelstein, S.; David, M.; Spirer, Z.; Golander, A. 1α, 25-Dihydroxyvitamin D3, and 24,25-Dihydroxyvitamin D3 in Vitro Synthesis by Human Decidua and Placenta. Nature 1979, 281, 317–319.
- Howard, G.A.; Turner, R.T.; Sherrard, D.J.; Baylink, D.J. Human Bone Cells in Culture Metabolize 25-Hydroxyvitamin D3 to 1,25-Dihydroxyvitamin D3 and 24,25-Dihydroxyvitamin D3. J. Biol. Chem. 1981, 256, 7738–7740.
- Turner, R.T.; Puzas, J.E.; Forte, M.D.; Lester, G.E.; Gray, T.K.; Howard, G.A.; Baylink, D.J. In Vitro Synthesis of 1α,25-Dihydroxycholecalciferol and 24,25-Dihydroxycholecalciferol by Isolated Calvarial Cells. Proc. Natl. Acad. Sci. USA 1980, 77, 5720–5724.
- Reichel, H.; Koeffler, H.P.; Norman, A.W. Synthesis in Vitro of 1,25-Dihydroxyvitamin D3 and 24,25-Dihydroxyvitamin D3 by Interferon-γ-Stimulated Normal Human Bone Marrow and Alveolar Macrophages. J. Biol. Chem. 1987, 262, 10931–10937.
- Bikle, D.D.; Nemanic, M.K.; Whitney, J.O.; Elias, P.W. Neonatal Human Foreskin Keratinocytes Produce 1,25-Dihydroxyvitamin D3. Biochemistry 1986, 25, 1545–1548.
- Bikle, D.D.; Nemanic, M.K.; Gee, E.; Elias, P. 1,25-Dihydroxyvitamin D3 Production by Human Keratinocytes. Kinetics and Regulation. J. Clin. Investig. 1986, 78, 557–566.
- Hollis, B.W.; Wagner, C.L.; Drezner, M.K.; Binkley, N.C. Circulating Vitamin D3 and 25-Hydroxyvitamin D in Humans: An Important Tool to Define Adequate Nutritional Vitamin D Status. J. Steroid Biochem. Mol. Biol. 2007, 103, 631–634.
- Jones, G. Pharmacokinetics of Vitamin D Toxicity. Am. J. Clin. Nutr. 2008, 88, 582S–586S.
- Horst, R.L.; Littledike, E.T.; Riley, J.L.; Napoli, J.L. Quantitation of Vitamin D and Its Metabolites and Their Plasma Concentrations in Five Species of Animals. Anal. Biochem. 1981, 116, 189–203.
- Antoniucci, D.M.; Black, D.M.; Sellmeyer, D.E. Serum 25-Hydroxyvitamin D Is Unaffected by Multiple Freeze-Thaw Cycles. Clin. Chem. 2005, 51, 258–261.
- Bikle, D.D.; Gee, E.; Halloran, B.; Kowalski, M.A.; Ryzen, E.; Haddad, J.G. Assessment of the Free Fraction of 25-Hydroxyvitamin D in Serum and Its Regulation by Albumin and the Vitamin D-Binding Protein. J. Clin. Endocrinol. Metab. 1986, 63, 954–959.
- Hollis, B.W. Comparison of Equilibrium and Disequilibrium Assay Conditions for Ergocalciferol, Cholecalciferol and Their Major Metabolites. J. Steroid Biochem. 1984, 21, 81–86.
- Hoy, D.A.; Ramberg, C.F., Jr.; Horst, R.L. Evidence That Discrimination against Ergocalciferol by the Chick Is the Result of Enhanced Metabolic Clearance Rates for Its Mono- and Dihydroxylated Metabolites. J. Nutr. 1988, 118, 633–638.
- Chun, R.F.; Lauridsen, A.L.; Suon, L.; Zella, L.A.; Pike, J.W.; Modlin, R.L.; Martineau, A.R.; Wilkinson, R.J.; Adams, J.; Hewison, M. Vitamin D-Binding Protein Directs Monocyte Responses to 25-Hydroxy- and 1,25-Dihydroxyvitamin D. J. Clin. Endocrinol. Metab. 2010, 95, 3368–3376.
- Vanham, G.; Van Baelen, H.; Tan, B.K.; Bouillon, R. The Effect of Vitamin D Analogs and of Vitamin D-Binding Protein on Lymphocyte Proliferation. J. Steroid Biochem. 1988, 29, 381–386.
- Vargas, S.; Bouillon, R.; Van Baelen, H.; Raisz, L.G. Effects of Vitamin D-Binding Protein on Bone Resorption Stimulated by 1,25 Dihydroxyvitamin D3. Calcif. Tissue Int. 1990, 47, 164–168.
- Adams, J.S.; Chen, H.; Chun, R.; Ren, S.; Wu, S.; Gacad, M.; Nguyen, L.; Ride, J.; Liu, P.; Modlin, R.; et al. Substrate and Enzyme Trafficking as a Means of Regulating 1,25-Dihydroxyvitamin D Synthesis and Action: The Human Innate Immune Response. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2007, 22 (Suppl. S2), V20–V24.
- Sirajudeen, S.; Shah, I.; Al Menhali, A. A Narrative Role of Vitamin D and Its Receptor: With Current Evidence on the Gastric Tissues. Int. J. Mol. Sci. 2019, 20, 3832.
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.-C.; Jurutka, P.W. Molecular Mechanisms of Vitamin D Action. Calcif. Tissue Int. 2013, 92, 77–98.
- Zmijewski, M.A.; Carlberg, C. Vitamin D Receptor(s): In the Nucleus but Also at Membranes? Exp. Dermatol. 2020, 29, 876–884.
- Chen, J.; Doroudi, M.; Cheung, J.; Grozier, A.L.; Schwartz, Z.; Boyan, B.D. Plasma Membrane Pdia3 and VDR Interact to Elicit Rapid Responses to 1α,25(OH)2D3. Cell. Signal. 2013, 25, 2362–2373.
- Hu, W.; Zhang, L.; Li, M.X.; Shen, J.; Liu, X.D.; Xiao, Z.G.; Wu, D.L.; Ho, I.H.T.; Wu, J.C.Y.; Cheung, C.K.Y.; et al. Vitamin D3 Activates the Autolysosomal Degradation Function against Helicobacter Pylori through the PDIA3 Receptor in Gastric Epithelial Cells. Autophagy 2019, 15, 707–725.
- Wu, W.; Beilhartz, G.; Roy, Y.; Richard, C.L.; Curtin, M.; Brown, L.; Cadieux, D.; Coppolino, M.; Farach-Carson, M.C.; Nemere, I.; et al. Nuclear Translocation of the 1,25D3-MARRS (Membrane Associated Rapid Response to Steroids) Receptor Protein and NFκB in Differentiating NB4 Leukemia Cells. Exp. Cell Res. 2010, 316, 1101–1108.
- Siu-Caldera, M.L.; Zou, L.; Ehrlich, M.G.; Schwartz, E.R.; Ishizuka, S.; Reddy, G.S. Human Osteoblasts in Culture Metabolize Both 1α,25-Dihydroxyvitamin D3 and Its Precursor 25-Hydroxyvitamin D3 into Their Respective Lactones. Endocrinology 1995, 136, 4195–4203.
- Clements, M.R.; Chalmers, T.M.; Fraser, D.R. Enterohepatic Circulation of Vitamin D: A Reappraisal of the Hypothesis. Lancet 1984, 323, 1376–1379.
- Hahn, T.J.; Birge, S.J.; Scharp, C.R.; Avioli, L.V. Phenobarbital-Induced Alterations in Vitamin D Metabolism. J. Clin. Investig. 1972, 51, 741–748.
- Horst, R.L.; Goff, J.P.; Reinhardt, T.A. Calcium and Vitamin D Metabolism in the Dairy Cow. J. Dairy Sci. 1994, 77, 1936–1951.
- Horst, R.L.; Reinhardt, T.A.; Reddy, G.S. CHAPTER 2—Vitamin D Metabolism. In Vitamin D (Second Edition); Feldman, D., Ed.; Academic Press: Burlington, VT, USA, 2005; pp. 15–36. ISBN 978-0-12-252687-9.
- Morgan, J.W.; Reddy, G.S.; Uskokovic, M.R.; May, B.K.; Omdahl, J.L.; Maizel, A.L.; Sharma, S. Functional Block for 1α,25-Dihydroxyvitamin D3-Mediated Gene Regulation in Human B Lymphocytes. J. Biol. Chem. 1994, 269, 13437–13443.
- St-Arnaud, R.; Arabian, A.; Travers, R.; Barletta, F.; Raval-Pandya, M.; Chapin, K.; Depovere, J.; Mathieu, C.; Christakos, S.; Demay, M.B.; et al. Deficient Mineralization of Intramembranous Bone in Vitamin D-24-Hydroxylase-Ablated Mice Is Due to Elevated 1,25-Dihydroxyvitamin D and Not to the Absence of 24,25-Dihydroxyvitamin D. Endocrinology 2000, 141, 2658–2666.
- Cutolo, M.; Plebani, M.; Shoenfeld, Y.; Adorini, L.; Tincani, A. Chapter Fourteen—Vitamin D Endocrine System and the Immune Response in Rheumatic Diseases. In Vitamins & Hormones; Litwack, G., Ed.; Vitamins and the Immune System; Academic Press: Cambridge, MA, USA, 2011; Volume 86, pp. 327–351.
- Beck, G.R.; Zerler, B.; Moran, E. Phosphate Is a Specific Signal for Induction of Osteopontin Gene Expression. Proc. Natl. Acad. Sci. USA 2000, 97, 8352–8357.
- Gil, Á.; Plaza-Diaz, J.; Mesa, M.D. Vitamin D: Classic and Novel Actions. Ann. Nutr. Metab. 2018, 72, 87–95.
- Conigrave, A.D. The Calcium-Sensing Receptor and the Parathyroid: Past, Present, Future. Front. Physiol. 2016, 7, 563.
- Adams, J.S.; Sharma, O.P.; Gacad, M.A.; Singer, F.R. Metabolism of 25-Hydroxyvitamin D3 by Cultured Pulmonary Alveolar Macrophages in Sarcoidosis. J. Clin. Investig. 1983, 72, 1856–1860.
- Insogna, K.L.; Dreyer, B.E.; Mitnick, M.; Ellison, A.F.; Broadus, A.E. Enhanced Production Rate of 1,25-Dihydroxyvitamin D in Sarcoidosis. J. Clin. Endocrinol. Metab. 1988, 66, 72–75.
- Gyetko, M.R.; Hsu, C.H.; Wilkinson, C.C.; Patel, S.; Young, E. Monocyte 1α-Hydroxylase Regulation: Induction by Inflammatory Cytokines and Suppression by Dexamethasone and Uremia Toxin. J. Leukoc. Biol. 1993, 54, 17–22.
- Noyola-Martínez, N.; Díaz, L.; Zaga-Clavellina, V.; Avila, E.; Halhali, A.; Larrea, F.; Barrera, D. Regulation of CYP27B1 and CYP24A1 Gene Expression by Recombinant Pro-Inflammatory Cytokines in Cultured Human Trophoblasts. J. Steroid Biochem. Mol. Biol. 2014, 144, 106–109.
- Stoffels, K.; Overbergh, L.; Giulietti, A.; Verlinden, L.; Bouillon, R.; Mathieu, C. Immune Regulation of 25-Hydroxyvitamin-D3-1α-Hydroxylase in Human Monocytes. J. Bone Miner. Res. 2006, 21, 37–47.
- Nelson, C.D.; Reinhardt, T.A.; Thacker, T.C.; Beitz, D.C.; Lippolis, J.D. Modulation of the Bovine Innate Immune Response by Production of 1α,25-Dihydroxyvitamin D3 in Bovine Monocytes. J. Dairy Sci. 2010, 93, 1041–1049.
- Wherry, T.L.T.; Dassanayake, R.P.; Casas, E.; Mooyottu, S.; Bannantine, J.P.; Stabel, J.R. Exogenous Vitamin D3 Modulates Response of Bovine Macrophages to Mycobacterium avium Subsp. paratuberculosis Infection and Is Dependent Upon Stage of Johne’s Disease. Front. Cell. Infect. Microbiol. 2022, 11, 1446.
- Fabri, M.; Stenger, S.; Shin, D.-M.; Yuk, J.-M.; Liu, P.T.; Realegeno, S.; Lee, H.-M.; Krutzik, S.R.; Schenk, M.; Sieling, P.A.; et al. Vitamin D Is Required for IFN-γ–Mediated Antimicrobial Activity of Human Macrophages. Sci. Transl. Med. 2011, 3, 104ra102.
- Stabel, J.R.; Reinhardt, T.A.; Hempel, R.J. Short Communication: Vitamin D Status and Responses in Dairy Cows Naturally Infected with Mycobacterium avium Ssp. paratuberculosis. J. Dairy Sci. 2019, 102, 1594–1600.
- Nelson, C.D.; Reinhardt, T.A.; Beitz, D.C.; Lippolis, J.D. In Vivo Activation of the Intracrine Vitamin D Pathway in Innate Immune Cells and Mammary Tissue during a Bacterial Infection. PLoS ONE 2010, 5, e15469.
- Merriman, K.E.; Kweh, M.F.; Powell, J.L.; Lippolis, J.D.; Nelson, C.D. Multiple β-Defensin Genes Are Upregulated by the Vitamin D Pathway in Cattle. J. Steroid Biochem. Mol. Biol. 2015, 154, 120–129.
- Nelson, C.D.; Nonnecke, B.J.; Reinhardt, T.A.; Waters, W.R.; Beitz, D.C.; Lippolis, J.D. Regulation of Mycobacterium-Specific Mononuclear Cell Responses by 25-Hydroxyvitamin D3. PLoS ONE 2011, 6, e21674.
- Merriman, K.E.; Poindexter, M.B.; Kweh, M.F.; Santos, J.E.P.; Nelson, C.D. Intramammary 1,25-Dihydroxyvitamin D3 Treatment Increases Expression of Host-Defense Genes in Mammary Immune Cells of Lactating Dairy Cattle. J. Steroid Biochem. Mol. Biol. 2017, 173, 33–41.
- Wherry, T.L.T.; Heggen, M.; Shircliff, A.L.; Mooyottu, S.; Stabel, J.R. Stage of Infection with Mycobacterium avium Subsp. paratuberculosis Impacts Expression of Rab5, Rab7, and CYP27B1 in Macrophages Within the Ileum of Naturally Infected Cows. Vet. Microbiol. 2022. manuscript in review.
- Adams, J.S.; Hewison, M. Update in Vitamin D. J. Clin. Endocrinol. Metab. 2010, 95, 471–478.
- Hollis, B.W. Circulating 25-Hydroxyvitamin D Levels Indicative of Vitamin D Sufficiency: Implications for Establishing a New Effective Dietary Intake Recommendation for Vitamin D. J. Nutr. 2005, 135, 317–322.
- Nelson, C.D.; Reinhardt, T.A.; Lippolis, J.D.; Sacco, R.E.; Nonnecke, B.J. Vitamin D Signaling in the Bovine Immune System: A Model for Understanding Human Vitamin D Requirements. Nutrients 2012, 4, 181–196.
- Vieth, R. Why the Minimum Desirable Serum 25-Hydroxyvitamin D Level Should Be 75 Nmol/L (30 Ng/Ml). Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 681–691.
- Nelson, C.D.; Lippolis, J.D.; Reinhardt, T.A.; Sacco, R.E.; Powell, J.L.; Drewnoski, M.E.; O’Neil, M.; Beitz, D.C.; Weiss, W.P. Vitamin D Status of Dairy Cattle: Outcomes of Current Practices in the Dairy Industry. J. Dairy Sci. 2016, 99, 10150–10160.
- Nelson, C.D.; Powell, J.L.; Price, D.M.; Hersom, M.J.; Yelich, J.V.; Drewnoski, M.E.; Bird, S.L.; Bridges, G.A. Assessment of Serum 25-Hydroxyvitamin D Concentrations of Beef Cows and Calves across Seasons and Geographical Locations. J. Anim. Sci. 2016, 94, 3958–3965.
- Adams, J.S.; Ren, S.; Liu, P.T.; Chun, R.F.; Lagishetty, V.; Gombart, A.F.; Borregaard, N.; Modlin, R.L.; Hewison, M. Vitamin D-Directed Rheostatic Regulation of Monocyte Antibacterial Responses. J. Immunol. Baltim. Md 1950 2009, 182, 4289–4295.
- Liu, P.T.; Stenger, S.; Tang, D.H.; Modlin, R.L. Cutting Edge: Vitamin D-Mediated Human Antimicrobial Activity against Mycobacterium tuberculosis Is Dependent on the Induction of Cathelicidin. J. Immunol. 2007, 179, 2060–2063.
- Scott, M.G.; Davidson, D.J.; Gold, M.R.; Bowdish, D.; Hancock, R.E.W. The Human Antimicrobial Peptide LL-37 Is a Multifunctional Modulator of Innate Immune Responses. J. Immunol. 2002, 169, 3883–3891.
- Wan, M.; van der Does, A.M.; Tang, X.; Lindbom, L.; Agerberth, B.; Haeggström, J.Z. Antimicrobial Peptide LL-37 Promotes Bacterial Phagocytosis by Human Macrophages. J. Leukoc. Biol. 2014, 95, 971–981.
- Scocchi, M.; Wang, S.; Zanetti, M. Structural Organization of the Bovine Cathelicidin Gene Family and Identification of a Novel Member. FEBS Lett. 1997, 417, 311–315.
- Whelehan, C.J.; Barry-Reidy, A.; Meade, K.G.; Eckersall, P.; Chapwanya, A.; Narciandi, F.; Lloyd, A.T.; O’Farrelly, C. Characterisation and Expression Profile of the Bovine Cathelicidin Gene Repertoire in Mammary Tissue. BMC Genom. 2014, 15, 128.
- Elsik, C.G.; Tellam, R.L.; Worley, K.C. The Genome Sequence of Taurine Cattle: A Window to Ruminant Biology and Evolution. Science 2009, 324, 522–528.
- Patil, A.A.; Cai, Y.; Sang, Y.; Blecha, F.; Zhang, G. Cross-Species Analysis of the Mammalian β-Defensin Gene Family: Presence of Syntenic Gene Clusters and Preferential Expression in the Male Reproductive Tract. Physiol. Genom. 2005, 23, 5–17.
- García-Barragán, Á.; Gutiérrez-Pabello, J.A.; Alfonseca-Silva, E. Calcitriol Increases Nitric Oxide Production and Modulates Microbicidal Capacity against Mycobacterium bovis in Bovine Macrophages. Comp. Immunol. Microbiol. Infect. Dis. 2018, 59, 17–23.
- Xu, H.; Soruri, A.; Gieseler, R.K.H.; Peters, J.H. 1,25-Dihydroxyvitamin D3 Exerts Opposing Effects to IL-4 on MHC Class-II Antigen Expression, Accessory Activity, and Phagocytosis of Human Monocytes. Scand. J. Immunol. 1993, 38, 535–540.
- Chandra, G.; Selvaraj, P.; Jawahar, M.S.; Banurekha, V.V.; Narayanan, P.R. Effect of Vitamin D3 on Phagocytic Potential of Macrophages with Live Mycobacterium tuberculosis and Lymphoproliferative Response in Pulmonary Tuberculosis. J. Clin. Immunol. 2004, 24, 249–257.
- Small, A.G.; Harvey, S.; Kaur, J.; Putty, T.; Quach, A.; Munawara, U.; Perveen, K.; McPhee, A.; Hii, C.S.; Ferrante, A. Vitamin D Upregulates the Macrophage Complement Receptor Immunoglobulin in Innate Immunity to Microbial Pathogens. Commun. Biol. 2021, 4, 401.
- Zeng, Z.; Surewaard, B.G.J.; Wong, C.H.Y.; Geoghegan, J.A.; Jenne, C.N.; Kubes, P. CRIg Functions as a Macrophage Pattern Recognition Receptor to Directly Bind and Capture Blood-Borne Gram-Positive Bacteria. Cell Host Microbe 2016, 20, 99–106.
- Kim, E.W.; Teles, R.M.B.; Haile, S.; Liu, P.T.; Modlin, R.L. Vitamin D Status Contributes to the Antimicrobial Activity of Macrophages against Mycobacterium leprae. PLoS Negl. Trop. Dis. 2018, 12, e0006608.
- Leidi, M.; Gotti, E.; Bologna, L.; Miranda, E.; Rimoldi, M.; Sica, A.; Roncalli, M.; Palumbo, G.A.; Introna, M.; Golay, J. M2 Macrophages Phagocytose Rituximab-Opsonized Leukemic Targets More Efficiently than M1 Cells In Vitro. J. Immunol. 2009, 182, 4415–4422.
- Wherry, T.L.T.; Dassanayake, R.P.; Bannantine, J.P.; Mooyottu, S.; Stabel, J.R. Vitamin D3 Alters Macrophage Phenotype and Endosomal Trafficking Markers in Dairy Cattle Naturally Infected with Mycobacterium avium Subsp. paratuberculosis. Front. Cell. Infect. Microbiol. 2022. manuscript in review.
- Waters, W.R.; Nonnecke, B.J.; Rahner, T.E.; Palmer, M.V.; Whipple, D.L.; Horst, R.L. Modulation of Mycobacterium bovis-Specific Responses of Bovine Peripheral Blood Mononuclear Cells by 1,25-Dihydroxyvitamin D3. Clin. Diagn. Lab. Immunol. 2001, 8, 1204–1212.
- Nonnecke, B.J.; Waters, W.R.; Foote, M.R.; Horst, R.L.; Fowler, M.A.; Miller, B.L. In Vitro Effects of 1,25-Dihydroxyvitamin D3 on Interferon-γ and Tumor Necrosis Factor-α Secretion by Blood Leukocytes from Young and Adult Cattle Vaccinated with Mycobacterium bovis BCG. Int. J. Vitam. Nutr. Res. 2003, 73, 235–244.
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-like Receptor Triggering of a Vitamin D-Mediated Human Antimicrobial Response. Science 2006, 311, 1770–1773.
- Crowle, A.J.; Ross, E.J.; May, M.H. Inhibition by 1,25(OH)2-Vitamin D3 of the Multiplication of Virulent Tubercle Bacilli in Cultured Human Macrophages. Infect. Immun. 1987, 55, 2945–2950.
- Rook, G.A.; Steele, J.; Fraher, L.; Barker, S.; Karmali, R.; O’Riordan, J.; Stanford, J. Vitamin D3, Gamma Interferon, and Control of Proliferation of Mycobacterium tuberculosis by Human Monocytes. Immunology 1986, 57, 159–163.
- Waters, W.R.; Palmer, M.V.; Nonnecke, B.J.; Whipple, D.L.; Horst, R.L. Mycobacterium bovis Infection of Vitamin D-Deficient NOS2−/− Mice. Microb. Pathog. 2004, 36, 11–17.
- Martinez, N.; Rodney, R.M.; Block, E.; Hernandez, L.L.; Nelson, C.D.; Lean, I.J.; Santos, J.E.P. Effects of Prepartum Dietary Cation-Anion Difference and Source of Vitamin D in Dairy Cows: Health and Reproductive Responses. J. Dairy Sci. 2018, 101, 2563–2578.
- Kweh, M.F.; Merriman, K.E.; Wells, T.L.; Nelson, C.D. Vitamin D Signaling Increases Nitric Oxide and Antioxidant Defenses of Bovine Monocytes. JDS Commun. 2021, 2, 73–79.
- Merriman, K.E.; Powell, J.L.; Santos, J.E.P.; Nelson, C.D. Intramammary 25-Hydroxyvitamin D3 Treatment Modulates Innate Immune Responses to Endotoxin-Induced Mastitis. J. Dairy Sci. 2018, 101, 7593–7607.
- Lippolis, J.D.; Reinhardt, T.A.; Sacco, R.A.; Nonnecke, B.J.; Nelson, C.D. Treatment of an Intramammary Bacterial Infection with 25-Hydroxyvitamin D3. PLoS ONE 2011, 6, e25479.