Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 3 by Jessie Wu and Version 2 by Jessie Wu.
Arsenic is a chemical element that is toxic, and long-term exposure to it causes cancers such as lung, skin, liver, and bladder cancers. Over 150 million people around the world are affected by arsenic exposure.
- arsenic
- carcinogenesis
- DNA methylation
1. Arsenic and Mechanisms of Arsenic-Induced Carcinogenesis
Arsenic (As), a chemical element, is classified as a toxic metalloid and is associated with various human cancers [1], and its toxicity depends on the molecular form and oxidation state. International Agency for Research on Cancer (IARC) and US Environmental Protection Agency (USEPA) designated arsenic as Group 1 and Group A human carcinogens, respectively [2][3][2,3]. Furthermore, it is graded as first on the substance priority list in the Agency for Toxic Substances and Disease Registry (ASTDR), USA (https://www.atsdr.cdc.gov/spl/index.html, accessed on 9 February 2022) [4]. Chronic exposure to dietary arsenic is linked to skin, bladder, liver, and lung cancer [4][5][6][7][4,5,6,7]. Drinking water contaminated with arsenic has been linked with increased mortality of both noncancerous diseases and cancers in Bangladesh [8]. Both chronic and acute exposure to arsenic is harmful to different tissues and organs in the body, such as alteration in skin pigmentation and hyperkeratosis, peripheral neuropathy, development, cognitive impairments, and cardiovascular diseases.
The contamination of arsenic increases with the finding of newer places [9]. The familiar sources of arsenic exposure include drinking water, food, and inhalation in an industrial work setting. Over 150 million people on the earth are exposed to carcinogenic (10 μg/L) levels of arsenic [9][10][9,10], and the majority of these people are affected by drinking water from aquifers contaminated with arsenic. Countries with arsenic concentrations exceeding this carcinogenic level (10 μg/L) in the drinking water include Bangladesh, India, China, Argentina, Mexico, Canada, the USA, and Chile [11]. Arsenic exposure to foods usually occurs by growing crops in the soil contaminated with arsenic and/or irrigating water contaminated with arsenic [12]. Furthermore, NIOSH estimates that approximately 1.5 million workers have been affected by arsenic or arsenic compounds [13].
Arsenic has several states; the most common valence states of arsenic are inorganic AsIII (arsenite) and AsV (arsenate). Inorganic arsenic is very toxic to humans, whereas organic arsenic has low toxicity. AsIII is the highest toxic form because it is more soluble in water than arsenic compounds. It contains a lone electron pair that can engage in chemical bonds [14][15][14,15]. Depending on the types of food, arsenic can be found in both inorganic (when combined with oxygen, chlorine, and sulfur, among other elements) and organic forms (when linked with carbon and hydrogen). Inorganic arsenic is typically found in the inorganic form in drinking water, soil, and some terrestrial foods such as rice, as either AsIII or AsV. Inorganic pentavalent arsenic AsV is absorbed by the body through drinking water and uses membrane transporters such as aquaporin and inorganic phosphate transporters (PiT) to enter the cells [16][17][16,17]. In the cell, arsenic AsV is converted to the more toxic form arsenite in a glutathione-dependent reaction (GSH), with subsequent methylation to mono-methylated (MMA) and di-methylated arsenicals (DMA), respectively [18][19][18,19]. Methylated arsenicals, especially MMAIII, are considered more toxic than inorganic AsIII both in vivo (in hamsters) [20] and in vitro (human cell lines) [21].
The mechanisms by which arsenic induces carcinogenesis are still a point of debate. However, it has been known that arsenical compounds contribute to carcinogenesis by disrupting the signaling cascade, changing gene expression, elevating levels of oxidative stress and inflammation, increasing genotoxic and DNA damage, decreasing DNA repair, inducing cell cycle arrest and apoptosis [19][22][23][24][19,22,23,24], acting as co-carcinogenesis with other environmental toxicants [25], and alterations of epigenetic regulation. There is also doubt about whether arsenic is genotoxic or not because arsenic does not cause point mutations in standard mutagenicity assays; hence it is considered to be nongenotoxic [26][27][26,27]. Although arsenic is viewed as a carcinogen, its non-mutagenic characteristics violet its function in causing genetic alteration. However, a less studied mechanism, but one that is crucial for understanding arsenic-induced carcinogenesis, is the dysregulation of epigenetic modifications. Studies investigating epigenetic regulation changes upon arsenic exposure during the last decade are increasing. The researchers have attempted to explore the role of DNA methylation, histone modification, miRNAs and lncRNAs alteration, mRNA modification, and alternative splicing in arsenic toxicity and carcinogenesis. The present review will comprehensively discuss the epigenetic regulations involved in gene expression, and their dysregulation is pivotal in arsenic-induced transformation, tumor growth, and angiogenesis. The general scheme of the mechanism of arsenic-induced carcinogenesis is shown as follows (Figure 1).
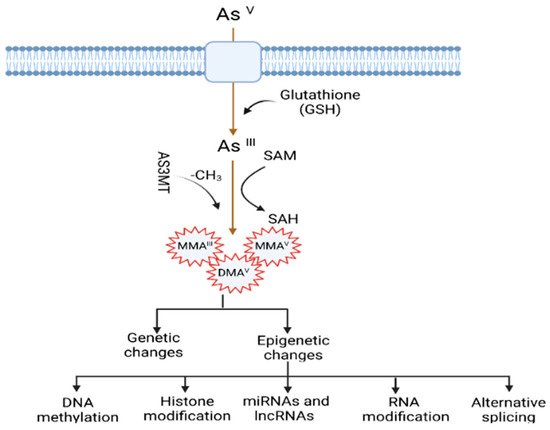
Figure 1. Mechanisms of arsenic-induced carcinogenesis. Arsenic exposure induces carcinogenesis via its biotransformation process, which causes effects on both genetic and epigenetic levels. The biotransformation of arsenic happens via a series of reactions such as reduction, oxidation, and methylation. Pentavalent arsenic (AsV) is reduced to trivalent (AsIII) and then methylated into organic arsenic species with higher carcinogenic potential. Here, S-adenosylmethionine (SAM) acts as a methyl donor, and Glutathione (GSH) and other thiols serve as reducing agents. Epigenetic alterations induced by arsenic exposure include abnormal changes in DNA methylation, histone modification, miRNAs and lncRNAs expression, RNA modification, and alternative splicing.
2. Arsenic-Induced Changes in DNA Methylation
DNA methylation is the inclusion of the methyl group (-CH3) in the 5-carbon on the cytosine residues (5 mC) in CpG (Cytosine-Phosphate-Guanine) and non-CpG (CpA, CpT, and CpC) dinucleotides. The methyl group comes from a methyl donor, generally from S-adenyl methionine (SAM), and this process is mediated by DNA methyl transferases (DNMTs) [28]. CpG dinucleotides are concentrated in CpG islands (short CpG-rich DNA stretches) and regions of repetitive sequences such as centromeric repeats, retrotransposon elements, rDNA, etc. [29][30][31][29,30,31]. In cancers, the changes of methylation status mainly occur within CpG islands, which occupy ~70% of all mammalian promotors. In addition, these islands play an important role in the regulation of transcription, and their general changes have been found during malignant transformation [32][33][32,33]. The functional effect of the dysregulation of DNA methylation is context- and spatial-dependent, dynamic, tissue-specific, and trans-generationally heritable [34][35][36][34,35,36]. Generally, gene silencing involves promotor methylation, and constitutive gene expression is associated with gene body methylation [32]. However, the methylation of the gene body may also be found to inactivate repetitive DNA elements within the gene body [35][37][35,37] and show dramatic alteration intron-exon boundaries [38]. These complex methylation patterns underline the necessity of DNA methylation profiling to answer biological questions.
Although it is evident that the dysregulation of DNA methylation has been demonstrated in different cancers, our knowledge of the impact of inorganic arsenic (iAs) on DNA methylation is still growing. Methyl transferase (MTs) catalyze the methyl group transfer in the 5-carbon on the cytosine residues (5 mC) in CpG dinucleotides and use SAM, a coenzyme, as a methyl group donor. Long-term exposure to arsenic causes depletion of the SAM by MTs such as AS3MT [arsenic (III) methyl transferase]. Furthermore, arsenic can also control DNMTs and decrease their activities. For example, studies have found that arsenic exposure causes a reduction of mRNA levels and activity of DNMTs [39][40][41][39,40,41].
iAs exposure has been shown to change global DNA methylation in vitro, in animal studies as well as in population studies (Table 1). For instance, a chronic low-dose of iAs exposure induces DNA hypo-methylation in cells [42]. In addition, fish, mice, and rats exposed to iAs exhibit hepatic global DNA hypomethylation [42][43][44][42,43,44]. However, limited studies are available for the human population compared to in vitro and animal studies. A recent study assessed the association between arsenic exposure and global DNA methylation (∼850,000 CpGs) through drinking water among 396 Bangladeshi people who joined the Health Effects of Arsenic Longitudinal Study (HEALS). The study identified 34 CpGs associated with arsenic concentration in the urinary tract and found a positive relationship between higher arsenic concentration and DNA hypomethylation in those CpGs. Among the arsenic-associated CpGs, most of the genes were annotated to the reactive oxygen species (ROS) pathway, tumor necrosis factor-α (TNF-α) signaling, and inflammatory response via nuclear factor kappa B (NF-κB). These are essential hallmarks of cancer and aging [45]. The results are consistent with earlier studies indicating that epigenetic alterations potentially regulate arsenic toxicity [45]. Pilsner et al. showed that iAs exposure led to global hypomethylation of leukocytes in human skin. The authors observed that people with hypo methylation in the peripheral blood lymphocytes (PBL) DNA were prone to skin lesions two years later when they adjusted for age, urinary As, and other factors [46]. A whole-genome microarray-based study showed that the status of DNA methylation changed over time in people who were affected by arsenic-induced skin lesions compared to control in Bangladesh. The study found the top 20 differentially methylated CpG sites. Among these top CpG sites, the methylation percentages increased in 13 CpGs, and decreased in 7 CpGs between baseline and follow-up [47]. Bandyopadhyay et al. evaluated the association of cytogenetic damage by measuring lymphocyte micronucleus (MN) frequency and long interspersed nuclear element-1 (LINE-1) methylation status among children who were exposed to arsenic in the areas of West Bengal. They observed that a high reduction of LINE-1 methylation was associated with MN frequency in exposed children compared to unexposed children, suggesting that LINE-1 methylation is a potential epigenetic marker for arsenic toxicity in individuals [48].
Besides the changes in global DNA methylation status, iAs exposure also causes changes in DNA methylation in specific regions of targeted genes in different cancers [49]. For example, the association between arsenic exposure and hypomethylated or hypermethylated promotors of some genes was found in human skin cancer [50] and bladder cancer [51][52][51,52]. The carcinogenesis can occur due to the silence of tumor suppressor genes via hypermethylation [40]. Some studies have found that iAs exposure leads to increased methylation of the promotor for tumor suppressor genes such as p15, p16, p53, and death-associated protein kinase (DAPK) in vitro and in vivo [40][50][51][40,50,51], DNA repair-related genes such as ERCC2, RPA1 in human hepatocytes [53], MLH1 in whole human blood [54], and genes associated with the Wnt pathway like MYC and WNT2B [53]. However, another study involving a human population chronically exposed to arsenic demonstrated hypomethylation at the promoter of the DNA repair gene ERCC2 [55]. Smeester et al. comprehensively studied the status of DNA methylation within CpG islands for more than 14,000 genes among arsenic-exposed individuals with skin lesions and without skin lesions [56]. They identified 183 genes with differentially methylated CpG islands, of which 182 were hyper-methylated in individuals with signs of arsenicosis. Gene enrichment analysis showed that most genes involved cancer-linked pathways via genes such as p53. They also identified an arsenic-methylated tumor suppressorome, a complex of 17 known or putative tumor suppressors silenced in human cancers, which includes hypermethylated genes such as chromosome 11 open reading frame 70 (C11orf70), centromere protein E (CENPE), forkhead box F1 (FOXF1), homeobox B5 (HOXB5), homeobox B9 (HOXB9), hsa-mir-126, SWI/SNF related, matrix associated, actin dependent regulator of chromatin subfamily d member 2 (SMARCD2), T-box brain 1 (TBR1), etc. Chanda et al. showed the hypermethylation of GMDS gene fragments in the peripheral blood leukocyte DNA of individuals exposed to arsenic and with skin cancer. They indicated it as a biomarker for arsenic-induced cancer [57]. The AS3MT gene plays an essential role in the metabolism of arsenic and its toxicological mechanism. Gribble et al. found decreased methylation in the promotor region of AS3MT in an arsenic-exposed area in Arizona [58]. However, no further studies have been performed to investigate the association between skin lesion status and AS3MT promoter methylation to date. On the other hand, carcinogenesis can also occur due to the activation of oncogene genes via hypomethylation. For instance, mice treated with iAs showed hypomethylation of the promoter region of oncogene Hras1 and increased mRNA levels of Hras1 [59], which was consistent with another study showing hypomethylation and increased mRNA levels of Hras1 and c-myc in vitro [60][61][60,61]. Arsenic exposure also led to Esr1 gene overexpression via hypomethylation of its promoter region, which is closely related to arsenic-induced hepatocarcinogenesis [44]. However, a recent study by Janasik et al. found hypermethylation of genes promoter of Nuclear factor-erythroid factor 2-related factor 2 (NRF2) and Kelch-like ECH-associated protein 1 (KEAP1) among occupationally arsenic-exposed copper mill workers from Poland [62].
DNA methylation inhibition occurs in a site-specific manner by proteins known as the ten-eleven translocation (TET) enzymes [63][64][63,64]. These TET enzymes oxidize 5 mc to 5 hyrdoxymethylcytosine (5 hmc). Disruption of this group of proteins has been shown in different types of cancer. Wang et al. showed that As inhibited the TET-mediated DNA demethylation and subsequently induced the hypermethylation in the promotor region to suppress the antioxidant genes 8-oxoguanine DNA glycosylase (OGG1) and glutathione S-transferase Pi 1 (GSTP1), thus increasing oxidative stress in human bronchial epithelial (HBE) cells in vitro [65]. In another recent study, Domingo-Relloso et al. conducted an epigenome-wide association study (EWAS) to compare the association of different As exposure levels and human blood 5 mc and 5 hmc markers in two diverse populations from the Aragon Workers Health Study (AWHS, Spain) and the Folic Acid and Creatinine Trial (FACT, Bangladesh) [66]. The effect of As on site-specific 5 mC and 5 hmC was measured using the Illumina methylation EPIC array on more than 850,000 CpG sites. They indicated different epigenetic effects for low As exposure in the AWHS population and high As exposure in the FACT population. The differentially methylated (DMP) and hydroxymethylated (DHP) positions were primarily found in distinct genomic sites. For example, they found three DMPs annotated to CLEC12A, a gene that plays a role in inflammation and immune response, which was consistent with previous studies [67]. In addition, they also found one DHP annotated to NPLOC4, a gene that has protein processing function in the endoplasmic reticulum (ER) in the FACT population exposed to a high dose of As. This is invariable to a study that reported a role of As in ER stress-associated protein misfolding and apoptosis [68], for which mechanisms are known to be associated with cardiometabolic diseases and cancer.
In addition, arsenic exposure also causes transgenerational genotoxicity and the alteration of global DNA methylation patterns in the animal model. Parental chronic arsenic exposure led to genotoxic damage (F0–F3), different methylation patterns, changes in physical and reproductive parameters, abnormal morphology in the ovaries (F0 and F1) and testicles (F1–F3), and a decline in the quality of sperm (F0–F3, except F2), suggesting that an individual’s early life disruptions can negatively impact later generations’ health [36]. An association was found between low or high-dose exposure to arsenic during gestation with umbilical cord blood DNA methylation. There was increased DNA methylation in CpG sites of LINE-1 and, to a lesser extent, within the promotor region of p16 [69]. Studies also showed the sex-dependent association between arsenic exposure and cord blood DNA methylation status, and the impact was even more prominent in the boys than in the girls [70].
Arsenic doesn’t induce point mutations but causes deletion mutations and chromosomal instability [40]. One possible mechanism by which arsenical compounds contribute to carcinogenesis is the disruption of normal epigenetic marks at specific loci, which may cause changes in gene expression and carcinogenesis [71][72][71,72]. Although arsenic exposure was found to alter methylation levels in global DNA and promoters of some genes, current research is hard to understand due to the complexity and insubstantial information provided in the current studies. Further investigations are necessary to systematically explore DNA methylation on a genome-wide level in cell lines exposed to arsenic and target tissues from well-characterized arsenic-exposed populations or tumor tissues from arsenic-associated cancers. Such studies will assist in elucidating the possible biological effects of arsenic exposure on DNA methylation and carcinogenesis. Arsenic-induced alterations of DNA methylation status and carcinogenesis are summarized in Table 1.
Table 1.
Arsenic-induced alterations of DNA methylation status and carcinogenesis.
| Tissue/Cells | Source of Arsenic | DNA Methylation | References | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Global | Gene-Specific | ||||||||
| Hyper | Hypo | ||||||||
| Prostate epithelial cell line RWPE-1 | As | III | Hypo | [73] | [73 | [74] | ,74] | ||
| HaCaT keratinocytes | As | III | Hypo | [39] | |||||
| TRL 1215 rat liver epithelial cells | As | III | Hypo | [42] | |||||
| Goldfish | As | III | Hypo | [75] | |||||
| Fisher 344 rat | As | III | Hypo | [43] | |||||
| 129/SvJ mice | As | III | Hypo | [44] | |||||
| Blood samples | Drinking water | Hypo | [45] | ||||||
| Blood samples from skin lesion patients and control | 13 Hyper and 7 hypo methylation of CpG islands | [47] | |||||||
| Human | Hyper | [76] | |||||||
| Hypo (in skin lesion patients) | [46] | ||||||||
| Peripheral blood lymphocyte DNA from skin lesions and non-skin lesions | Drinking water (urine samples) | 182 genes out of 183 hypermethylated; Identified a silenced tumor suppressorome consists of 17 genes |
[56] | ||||||
| MMA | III | ZHCAN12 | and | C1QTNF6 | |||||
| Uroepithelial SV-HUC-1 cells | As | III | DAPK | [77] | |||||
| Hamster embryo cells | As | III | c-myc | and | Ha-ras | [61] | |||
| TRL 1215 rat liver epithelial cells | As | III | c-myc | [51] | |||||
| C57BL/6J mice | As | III | c-Ha-ras | [59] | |||||
| A/J mice | As | V | p16 | , | RASSF1 | [78] | |||
| C3H mice | As | III | ER | α | [79] | ||||
| Blood samples from the people of West Bengal, India | Drinking water | p53 | and | p21 | in skin cancer patients | [50] | |||
| Tissues from arsenic-induced skin lesions (cases) and with no skin lesions (controls) | Drinking water | DAPK | and | p16 | [80] | ||||
| Blood samples from copper mill workers and Non-occupationally exposed healthy controls in Poland | Copper mill (urine) | NRF2 | and | KEAP1 | [62] | ||||
| Blood samples from arsenic-exposed individuals (with and without skin lesions) | Drinking water (water, urine) | MLH1 | and | MSH2 | [81] | ||||
| Samples from bladder tumor | Drinking water (toenail) | RASSF1A | and | PRSS3 | [52] | ||||
| Cord blood lymphocytes | Drinking water (cord blood, nails, and hair) | p53 | [82] | ||||||
| Blood samples from the West Bengal population and HEK293 cell lines | Drinking water(water, urine), sodium arsenite, As | III | Increased | ERCC2 | expression | [55] | |||
| Blood samples from arsenic-exposed individuals (with and without skin lesions) | Drinking water (water, urine) | Increased | Tfam | and | PGC1α | expression | [83] | ||