Drought stress is a major abiotic stress that significantly affects agricultural productivity every year as the plants undergo several morphological, biochemical, and physiological modifications, such as repressed root and shoot growth, reduced photosynthesis and transpiration rate, excessive production of reactive oxygen species (ROS), osmotic adjustments, and modified leaf senescence regulating and stress signaling pathways. Such modifications may permanently damage the plants; therefore, mitigation strategies must be developed. The use of drought-resistant crop cultivars is more expensive and labor-intensive with few advantages. However, exploiting plant growth promoting rhizobacteria (PGPR) is a proven alternative with numerous direct and indirect advantages.
1. Introduction
A drought is an instance of major abiotic stress encountered by the plants throughout their life cycle due to inadequate water availability. This stress lowers crop development and yield by negatively affecting the quality of morphological, physiological, and biochemical attributes
[1][2][10,11]. Hence, it is critical to undertake a real paradigm shift towards agricultural sustainability and to solve water shortage and food security problems. To meet ever-increasing global food requirements, it is necessary to develop effective eco-friendly strategies that can augment the levels of plant endurance during water-deficits and improve crop growth and production
[3][12]. Developing drought-resistant crops, shifting crop cultivation, and establishing resource management practices are some strategies to cope with drought stress
[1][4][10,13]. However, most of them have disadvantages, such as laborious, time-consuming, cross-contamination and loss of beneficial host plant characteristics. These encourage the utilization of plant growth promoting rhizobacteria (PGPR) as cost-effective, a less laborious and promising approach for providing additional drought stress tolerance benefits along with improved plant performance
[5][6][7][8][9,14,15,16]. Therefore, they have often been considered essential tools for facilitating sustainable agriculture. The
presen
ext sectiont review mainly focuses on the current knowledge concerning the various mechanisms of PGPR to counteract the negative impacts of drought stress on host plants and enhance agricultural productivity in dry lands.
2. Plant Growth Promoting Rhizobacteria Mediated Drought Stress Tolerance in Plants
One of the plant’s primary defense responses subjected to drought is the stomatal closure, which reduces the uptake and fixation of carbon dioxide (CO
2), and photosynthesis, thus potentially causing whole plant death in severe drought stress. The plants use efficient adaptive mechanisms to better cope with drought stress, including the changes in shoot and root morphology, reactive oxygen species (ROS) production, synthesis of stress hormones and activating antioxidant defense system
[5][9]. The significant socioeconomic impact of drought stress on agriculture also results in huge monetary losses as it is incurred with long-term effects. The PGPR are the natural habitants of the rhizosphere soil that rely on root exudates to fuel their metabolic activities
[9][10][11][17,18,19]. The great diversity of PGPR is influenced by soil physiological conditions and nutrient availability in the rhizosphere
[12][13][20,21]. These PGPR provide the plants with various growth promoting benefits, primarily through enabling root colonization. These bacteria act as biofertilizers and biostimulants that are most beneficial for agricultural sustainability by promoting the overall plant growth and yield directly or indirectly under severe drought stress
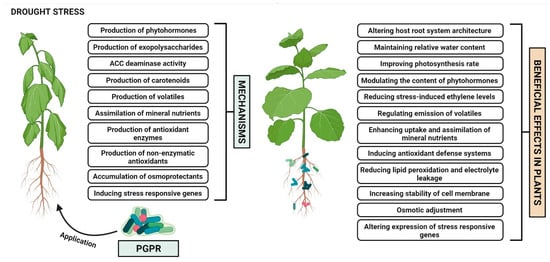
[3][14][15][12,22,23]. The most prominent mechanisms of PGPR employed for mitigating drought stress (
Figure 1) are elaborated below.
Figure 1. Mechanisms of PGPR-mediated induction of plant drought stress tolerance.
2.1. Alteration of Host Root System Architecture
The plant phenotypic response to water stress was mainly associated with the alteration of their root system architecture due to the production of ROS. One of the first steps toward improved drought stress tolerance is the bacterial-mediated alteration in host root system architecture
[16][17][24,25]. The PGPR candidates of
Bacillus sp. and
Enterobacter sp. displayed alteration of root system architecture in
Triticum aestivum and
Zea mays that facilitated the drought avoidance phenotypes viz., enhanced root surface area, root branching, root length, and many root tips when compared to the untreated control plants
[18][26]. Many studies have revealed that the PGPR enhance plant cell membrane stability and root nodulation by modulating the level of phytohormones and activating the antioxidant defense mechanism that is directly correlated with increasing plant drought tolerance
[19][20][27,28]. These research findings implied the possible role of PGPR in mitigating plant stress tolerance through altering their root architecture.
2.2. Maintenance of Relative Water Content
The relative water content is a useful indicator of leaf water status in the plant as it is involved in plant metabolism. The drought causes a fall in relative water content, often followed by a reduction in transpiration rate and leaf water potential
[21][29]. The turgor pressure also decreases in the plant cells when water levels fall, which consequently causes cell damage, wilting, and a decline in plant development
[22][30]. The drought usually causes a decrease in hydraulic conductivity, which alters the relative water content of the plant. Drought-induced ROS and osmotic stress are thwarted by the plants’ high relative water content, which may increase crop productivity
[23][24][25][1,4,31]. As a result, relative water content measurements are crucial for determining the severity of drought stress. The PGPR regulates water potential by modifying the hydraulic conductivity, stomatal opening and transpiration rate that improve the plant survival rate in drought stress conditions
[26][32]. The literature indicated that the PGPR treatment effectively caused the root system development that improves the water uptake, enabling inoculated plants to tolerate drought stress
[16][27][24,33]. The exact mechanisms underlying the increased relative water content with PGPR application during drought stress have yet to be explored. However, it is believed that PGPR can help the plant increase its relative water content due to bacterial production of phytohormones and osmoprotectants and alleviate the drought stress
[28][34]. It has been noted that inoculation of
T. aestivum plants with
Azospirillum sp. significantly increased the relative water content attributed to the bacterial IAA production. Apart from increasing the relative water content, the bacterial treatment also enhanced root growth and the formation of lateral roots, which help uptake water and mineral nutrients under drought stress
[11][19]. Similarly,
Arabidopsis plants inoculated with ABA producing
A. brasilense improved the relative leaf water content, thereby ameliorating growth response under drought stress
[14][22], which confirmed the ability of phytohormone producing PGPR strains to enhance plant growth under drought stress conditions. Such studies emphasize the necessity of comprehending the mechanisms behind demonstrated PGPR-mediated drought tolerance via maintaining the high relative water content.
2.3. Improvement of Photosynthesis Rate
The plant’s primary response to drought stress is the promotion of stomatal closure to reduce water loss by transpiration
[29][35]. The negative effect of early partial stomatal closure is the reduction in CO
2 assimilation, which leads to a decrease in net photosynthetic capacity, but the favorable effect is a drop in transpiration rate, which helps to preserve water in plants
[30][36]. During drought stress, the rate of assimilatory surface growth slows first, followed by photosynthesis inhibition in plants. Drought-induced reduction in plant photosynthetic rate can be attributable to stomatal and non-stomatal limitations. Stomatal photosynthetic limitations are inadequate CO
2 assimilation rate in plant leaves and sub-stomatal cavities caused by early partial stomatal closure. Non-stomatal photosynthetic limitations occur when there is a decline in chloroplast activity, leaf nitrogen, synthesis of adenosine triphosphate (ATP), ribulose1,5-bisphosphate carboxylase/oxygenase (RuBisCO) activity, ribulose 1,5-bisphosphate (RuBP) synthesis, and impairment of photosystem I and II reactions. However, the effects of non-stomatal photosynthetic limitations are prominent during severe drought stress and the imposed limitations are difficult to alleviate and require prolonged recovery time.
The drought stress restricts CO
2 uptake, which results in a decline in photosynthetic rate and excess ROS accumulation in various organelles found in plants, especially chloroplasts, peroxisomes, and mitochondria
[31][37]. Excessive ROS accumulation disrupted the structure of thylakoid membranes, enzyme activity, and photosynthetic pigments. Under drought stress, PGPR increases the stomatal conductance, photosynthetic rate, and maximum potential quantum efficiency of photosystem II (Fv/Fm), and decreases the transpiration rate in plants by producing the phytohormones, which affect the essential structural and functional characteristics of their photosynthetic apparatus
[27][32][33][33,38,39]. Liu et al.
[34][40] demonstrated the 11.6% enhancement in stomatal conductance, 13% elevation of photosynthetic rate, and the reversal process of chlorophyll degradation when the drought-stressed
Sambucus williamsii plants were inoculated with cytokinin producing
Acinetobacter calcoaceticus strain. Thus, it may be deduced that PGPR help to improve the photosynthetic rates under drought stress due to their ability to produce phytohormones.
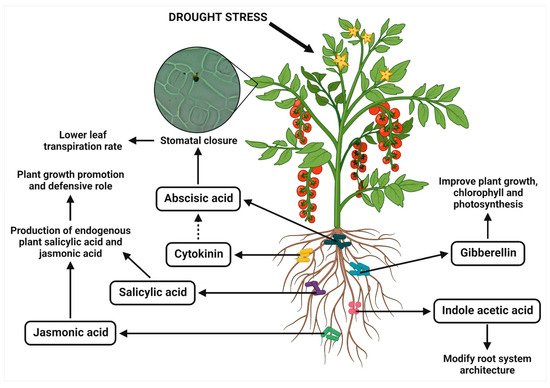
2.4. Production of Phytohormones
The phytohormones, such as auxin, cytokinin, gibberellin, abscisic acid (ABA), ethylene, salicylic acid, and jasmonic acid, are produced regularly in the plant system for their overall growth and development in response to their environmental stresses. The plant growth is promoted by auxin, cytokinin, and gibberellin, but ABA and ethylene inhibit growth
[35][41]. The drought stress reduces the level of these phytohormones, which has several negative impacts on plants
[21][29]. However, the application of PGPR can facilitate regulating the level of phytohormones by synthesizing and secreting them to improve the root system architecture of plants and assist them to thrive during drought (
Figure 2)
[36][37][42,43]. The reduction in ethylene production, altering the normal balance of cytokinins and ABA, and modification of the movement and concentration of IAA in host plants are associated with the endurance to drought stress upon applying PGPR. These changes may facilitate the further investigation of PGPR-mediated drought tolerance in plants. Moreover, the bacterially produced growth regulating phytohormones contributed to host plant metabolism and respiration
[38][39][44,45]. The different PGPR strains for modulating the plant hormonal levels in mitigating drought stress are listed in
Table 1.
Figure 2. PGPR-mediated modulation of phytohormones for enhancing drought tolerance in plants.
Table 1. PGPR-mediated production of phytohormones in enhancing plant drought stress tolerance.