Taro [Colocasia esculenta (L.) Schott] is the most commonly cultivated species in genus Colocasia and is the fourth most consumed tuber crop globally. It is a member of family Araceae, sub-family Aroideae, and is a tropical monocotyledonous, vegetatively propagated, perennial crop grown primarily for its starchy corm or underground stem. Taro is one of the world’s oldest food crops, with its domestication dating back over 9000 years. It was probably first domesticated in Southeast Asia and thereafter spread across the world, to become one of the most important staple food crops in the Pacific Islands. It is widely distributed across Africa, Oceania, Asia, and the Americas [4,5]. The crop has been largely maintained by smallholder farmers, and the species’ genetic resources have remained largely within local communities. In many societies, taro is considered a sacred plant of strong cultural importance and is used in religious festivals, domestic and agricultural rituals, and as bride price.
- taro
- taro leaf blight
- Phytophthora colocasiae
- taro improvement
- West Africa
1. Taro Production in Africa
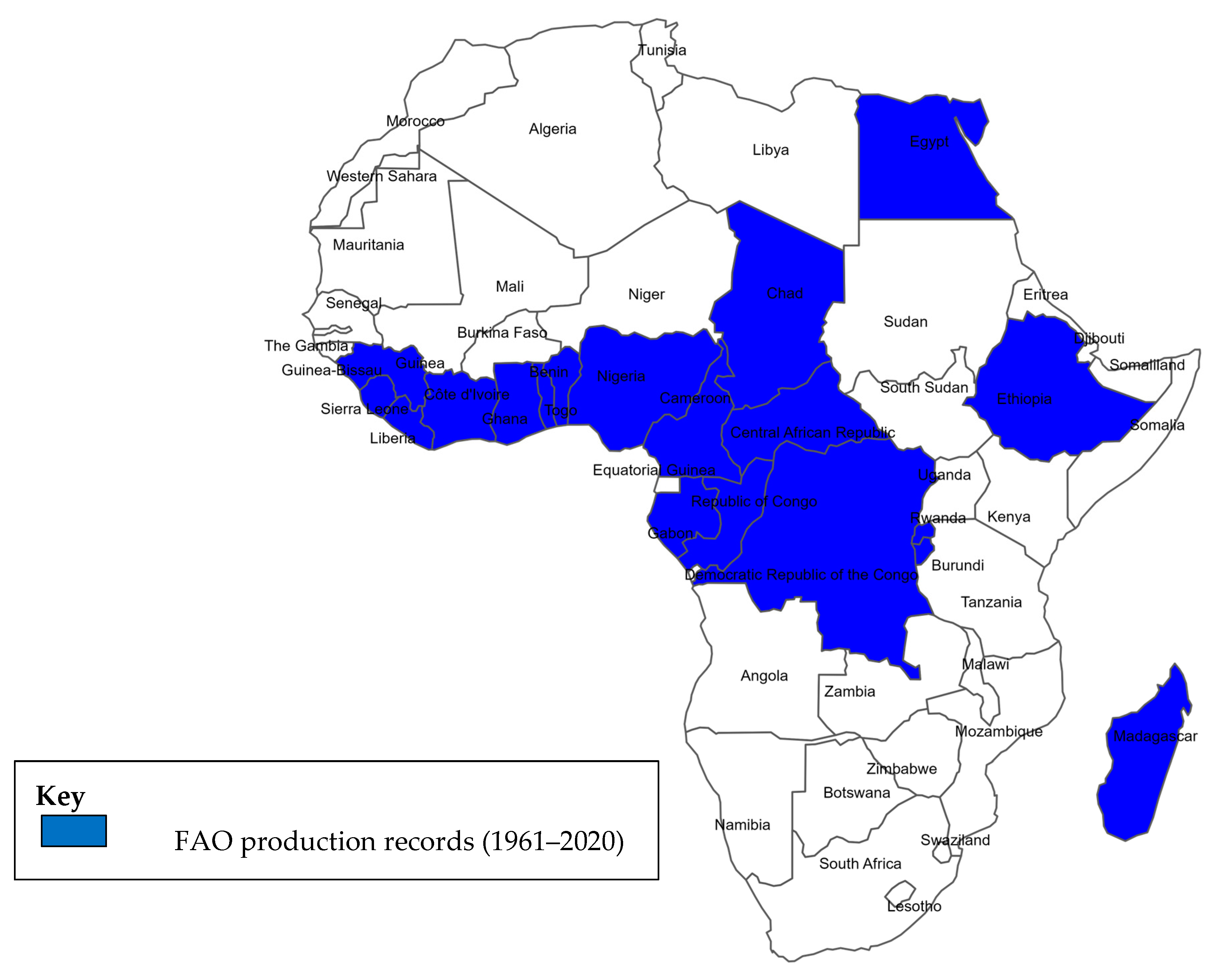
2. Taro Production Constraints in Africa
The major biotic stress for taro is TLB, a disease caused by P. colocasiae; it is an oomycete disease with highly devastating effects [44][14]. Phytophthora colocasiae was first reported in Java by Raciborski [45][15] and has now spread all over the tropics [46,47][16][17]. The disease thrives where day and night temperatures range between 25–28 °C and 20–22 °C, respectively, and can assume epidemic proportions all year round under favorable conditions [48][18]. TLB was not known in WA before 2009, when there were simultaneous outbreaks in Nigeria, Cameroon, Ghana, and other neighboring countries [20,21][19][20]. It is estimated that TLB in WA accounts for an economic loss of about USD 1.4 billion annually [19][21]. Phytophthora colocasiae reproduces asexually during rainy seasons, with the production of sporangia from sporangiophores at the extremity of lesions in infected leaves. Sporangia leave the pedicel during rain fall and germinate to produce motile zoospores that can swim for short distances in water and encyst to form germ tubes that can penetrate the host. This can happen within two hours at a favorable temperature of 20 °C and a minimum humidity of 90%. At an ideal temperature of 24–27 °C, symptoms present 2–4 days after penetration of the germ tube [46,47][16][17]. Phytophthora colocasiae is heterothallic with two mating types, A1 and A2, and can produce oospores via sexual reproduction [49][22]. In the field, P. colocasiae is spread mainly by zoospores and sporangia. The propagules are short-lived in the infected leaves and tissues and are carried by water to a host through rain splashes [47,50][17][23]. Taro corms are, however, rarely harvested from the field and can sustain the pathogen. Usually, planting is carried out within a short time frame after harvesting; any infected tissue debris left in the field is, therefore, a source of inoculum for subsequent infection of new plants [49,50][22][23]. An entire field can become symptomatic of the disease within seven days when conditions are favorable [19,48][18][21]. A symptomatic plant initially displays water-soaked lesions with a diameter of 1.5 cm around the leaf edges; the fluid exudates from the lesions are of bright-yellow to dark-purple colors when dried. As the disease progresses, the lesions enlarge, developing a zonate appearance characterized by brownish to purplish-brown colors. White fuzz also appears on both sides of the leaves, indicating sporangia, which continues to develop until the leaves are completely covered [46,48][16][18]. The white fuzz of sporangia around lesions is a characteristic symptom of P. colocasiae infection [46][16]. The infected leaf tissues collapse after 20 days, unlike healthy leaves, which last 40 days before senescence [50][23]. TLB also affects the corms, causing them to rot, and yield losses as a result of this disease can be as high as 70–100% [46,48,51][16][18][24]. TLB is usually controlled with the use of copper fungicides at a rate of 38 Lha−1, using 2.24 kg of copper oxychloride as the active ingredient. Fungicide applications start from four months after planting (MAPs) and continue until nine MAPs, weekly during rainfall periods and bi-weekly when conditions are dry. Dithane M-45 can also be used, at a rate of 1.68–2.25 kilograms in 189.3–378.5 liters of water per hectare. This application can be weekly or bi-weekly, depending on the severity of the disease, but should not exceed 25 applications and cannot be used once the crop is nine months old [48][18]. Metalaxyl fungicides have also been effectively used to control TLB [49][22]. In Hawaii, planting distance has been used as a control measure, with a decrease in disease incidence achieved by increasing the planting distance from 46 cm to 75 cm. In the Solomon Islands, improved sanitation, via pruning and removing infected leaves on a bi-weekly basis, has also reduced disease incidence. The use of resistant and immune varieties is the most viable control measure for TLB in terms of environmental impact and sustainability [43,52,53,54,55,56,57][25][26][27][28][29][30][31]. However, the use of resistant varieties has been limited by a lack of crop improvement programs and a lack of desirable economic market value traits in resistant genotypes. This is compounded by a lack of understanding of the genetic structure of pathogen populations [56][30]. Compared with other species of Phytophthora, very little attention has been paid to P. colocasiae, either globally or at a regional level. Some research has been carried out on screening for disease-resistance genotypes and their adaptability in WA and beyond. For example, Ackah et al. [58][32] evaluated taro genotypes from Ghana for resistance to TLB and found all the genotypes to be susceptible to varying degrees. Similarly, Amadi et al. [58][32] characterized some local and exotic collections of taro for yield, local adaptation, and TLB resistance in Nigeria and found some promising genotypes, although no single genotype combined all the desired traits. Additional major taro diseases are caused by viruses and other microorganisms that are specific to the Pacific [41][33]. These reduce corm size and quality, with yield losses of up to 20%. For example, the co-infection of taro with taro bacilliform virus (TaBV) and Colocasia bobone disease virus (CBDV) is thought to be lethal to the crop. TaBV, along with taro bacilliform CH virus (TaBCHV) diseases [59][34] and dasheen mosaic virus disease [10][35], has been reported in Africa. Several other taro viruses have also been found in the Pacific, which currently restricts international movement of germplasm; thus, many countries do not have access to agronomically elite genotypes and selected traditional cultivars. Taro diseases reported in the Pacific include taro soft rot, caused by several species of Pythium, sclerotium rot, caused by Sclerotium rolfsii, and cladosporium leaf spot, caused by Cladosporium colocasiae [41][33]. Taro soft root rot and cladosporium leaf spot have been reported in Africa [19,60][21][36].3. Opportunities for Improved Taro Production in WA
Among root crops, taro currently has the lowest average yield (5.4 tons/ha globally) [92][37]. WA countries dominate as the major producers of taro globally, producing 4.9 million tons of the estimated 12.0 million tons of taro produced in the world [79][38]. In the same year, Nigeria, the world’s largest producer of taro, harvested approximately 3.2 million tons from 0.8 million hectares, followed by Ghana with 1.68 million tons [81][39]. Currently, there is no taro germplasm repository in WA responsible for preserving, characterizing, and distributing taro germplasm. The International Institute of Tropical Agriculture (IITA) in collaboration with the National Root Crop Research Institute of Nigeria (NRCRI) had collected and preserved taro landraces. However, this collection was lost during the outbreak of TLB in West and Central Africa, including Nigeria [20][19], Ghana [21][20], and Cameroon [93][40]. Breeding resistant cultivars offers the best long-term control of TLB disease in most production systems; thus, urgent and collaborative efforts among research groups and donors are needed to combat the TLB epidemic and prevent taro from going extinct in the region. In recent years, scientists in WA and beyond have been gathering information, developing strategies, and evaluating stress factors to help to improve the taro crop [8,18,19,58,64,94,95,96,97,98][12][21][32][41][42][43][44][45][46][47]. Two of the improvements achieved to date are outlined as follows:- i.
-
Standardized collection protocols: Dansi [95][44] has outlined a collection procedure for root and tuber crops, including taro. The methodology is based on synthesized information from the publications of international bodies such as Biodiversity International, World Conservation Union (IUCN), FAO, and United Nations Environment Programme (UNEP). This helps to ensure that collection programs are executed using international standard procedures;
- ii.
-
Characterization of local and exotic germplasm for agro-morphological traits, disease resistance, and nutritional qualities, to be used as the basis for taro improvement: Several authors have used morphological characterization to evaluate taro cultivars, including agro-morphological traits, diseases, and flowering ability [58,75,90,97][32][46][48][49]. This has helped the classification of taro into different morphotypes, which can then be used in taro improvement programs.
4. Breeding Efforts in Taro
Taro breeding efforts should focus on traits that are important for both producers and consumers, such as yield, pest and disease resistance, nutritional quality, shelf life, plant architecture, maturity, and culinary characteristics [99][50]. Breeding programs involving different stakeholders can be used to gather more information and adopt new technologies. For example, there is a need to assess the acridity levels of different cultivars so that consumers can develop suitable cooking methods for increased edibility. Taro breeding was initiated in the late 1970s, and varieties were released in Fiji (1978), Samoa (1982; 1996), Solomon Islands (1978; 1992), Papua New Guinea (1993), and India (1995) [100,101][51][52]. The first successful controlled hybridization of taro in Nigeria was reported in 2015 [18][12]. Breeding schemes such as bi-parental crossing and recurrent selection were used at an early stage of plant development for traits such as taro corm flesh and corm fiber colors, which were correlated with the color of different petiole zones [102][53]. There have been several efforts in Papua New Guinea towards resistance breeding, but one of the difficulties is getting rid of deleterious traits from wild types. Breeding programs have been initiated in Hawaii for agronomic traits, and pests and disease resistance, to develop improved varieties for the restaurant and landscape trades [103][54]. The major objectives of taro breeding so far have been to improve plant architecture (such as optimal number of suckers, absence of stolons, optimal number of leaves, etc.), corm yield, resistance/tolerance to diseases such as TLB, and quality traits (such as dry-matter content, low levels of phenolic compounds causing irritation, etc.) [104][55]. Breeding activities with parents from a diverse genetic base could result in improved targeted traits. However, taro is usually propagated through vegetative means, seldom through flowers; the flowers are protogynous, making conventional breeding methods difficult [32][56]. Taro breeding has been initiated in many countries within the South Pacific under two major programs, TaroGen and TANSAO [105][57]. These programs have focused on hybridization to develop new cultivars with higher yields, better taste, and improved resistance to TLB [106,107][58][59]. Hybridization in taro is promising, but it is labor intensive and lengthy in terms of the induction of flowering (although gibberellic acid has been used to induce flowering in taro [18,47,75][12][17][48]), pollination, and seed harvesting. It takes 10 years or more from pollination for a new improved cultivar to be developed [108][60]. There is a scarcity of reports on taro breeding in WA, and the majority of the work is limited to the agro-morphological characterization or evaluation of local landraces. Amadi et al. [18][12] did achieve 109 crosses using 15 exotic taro cultivars and 4 local Nigerian cultivars, of which only 20 crosses (18.3%) were successful, with 9 crosses reaching maturity and producing seeds. The limited success with taro breeding can be linked to the weak institutional capacities of most national institutes engaged in breeding, coupled with a lack of genetic resources and funding for establishing a sustainable taro breeding program in WA. The work of existing national, regional, and international networks should also be consolidated to optimize breeding methodologies. The application of biotechnological techniques for the disease-free clonal propagation of taro plants is another viable option. Tuia [109][61] developed an efficient taro multiplication protocol and reported that it was possible to eliminate viruses using meristem cultures. There are also reports of somatic embryogenesis in taro [110[62][63][64],111,112], but the regeneration rate is low. Similarly, Fukino et al. [113][65] reported transformation in taro (C. esculenta var. antiquorum) calluses by particle bombardment, but only two putative transgenic plants were obtained. Transformation in Colocasia esculenta var. esculenta via microprojectile bombardment [114][66] and Agrobacterium tumefaciens [115,116][67][68] was also reported. These efforts show that using biotechnology to generate taro plantlets is a possibility, but there is a need to validate the vigor of plants in terms of growth rate, pest and disease resistance, and corm characteristics, in addition to extensive field trials to record the frequency of somaclonal variations [112][64].
References
- Grimaldi, I.M.; van Andel, T.R. Food and medicine by what name? The ethnobotanical and linguistic diversity of Taro in Africa. Econ. Bot. 2018, 72, 217–228.
- Grimaldi, I.M.; Leke, W.; Borokini, I.; Wanjama, D.; Van Andel, T.R. From landraces to modern cultivars: Field observations on taro Colocasia esculenta (L.) Schott in sub-Saharan Africa. Genet. Resour. Crop. Evol. 2018, 65, 1809–1828.
- Matthews, P.J. Written Records of Taro in the Eastern Mediterranean. In Ethnobotany: At the junction of the continents and the disciplines, Proceedings of the IVth International Congress of Ethnobotany, ICEB, Istanbul, Turkey, 21–26 August 2006; Ertug, Z.F., Ed.; Istanbul Yayinlari: İstanbul, Turkey, 2006; pp. 419–426.
- FAOSTAT. FAO Statistical Database. 2017. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 8 December 2021).
- FAOSTAT. FAO Statistical Database. 2022. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 8 April 2022).
- Grimaldi, I.M. Taro across the Oceans, journeys of one of our oldest crops. In News from the past, Progress in African Archaeobotany, Proceedings of the Seventh International Workshop on African Archaeobotany, Vienna, Austria, 2–5 July 2012; Advances in Archaeobotany 3; Barkhuis: Groningen, The Netherlands, 2016; pp. 67–81.
- Onwueme, I. Taro cultivation in Asia and the Pacific. RAP Publ. 1999, 16, 1–60. Available online: https://www.sciencedirect.com/science/article/pii/S240584402102733X-bbib18 (accessed on 9 September 2021).
- Matthews, P.J.; Ghanem, M.E. Perception gaps that may explain the status of taro (Colocasia esculenta) as an “orphan crop”. Plants People Planet 2021, 3, 99–112.
- Dagne, Y.; Mulualem, T.; Kifle, A. Development of high yielding Taro (Colocasia esculenta L.) Variety for mid altitude growing areas of Southern Ethiopia. J. Plant Sci. 2014, 2, 50–54.
- Maretta, D.; Sobir, S.; Helianti, I.; Purwono, P.; Santosa, E. Genetic diversity in Eddoe Taro (Colocasia esculenta var. antiquorum) from Indonesia based on morphological and nutritional characteristics. Biodiversitas. 2020, 21, 3525–3533. Available online: https://smujo.id/biodiv/article/view/5930 (accessed on 10 February 2021).
- Rao, R.V.; Peter, M.J.; Pablo, E.B.; Hunter, D. The Global Diversity of Taro: Ethnobotany and Conservation; Bioversity International: Rome, Italy, 2010.
- Amadi, C.O.; Onyeka, J.; Chukwu, G.O.; Okoye, B.C. Hybridization and Seed Germination of Taro (Colocasia esculenta) in Nigeria. J. Crop. Improv. 2015, 29, 106–116.
- Terry, E.R.; Oduro, K.A.; Caveness, F.E. Cocoyam Farming Systems in Nigeria. In Tropical root crops: Research Strategies for the 1980s, Proceedings of the First Triennial Root Crops Symposium of the International Society for Tropical Root Crops—Africa Branch, Ibadan, Nigeria, 8–12 September 1980; International Development Research Centre: Ottawa, Canada, 1981.
- Vetukuri, R.R.; Kushwaha, S.K.; Sen, D.; Whisson, S.C.; Lamour, K.H.; Grenville-Briggs, L.J. Genome Sequence Resource for the Oomycete Taro Pathogen Phytophthora colocasiae. Mol. Plant Microbe Interact. 2018, 31, 903–905.
- Raciborski, M. Parasitic algae and fungi. Java. Batavia Bull. N. Y. State Mus. 1900, 19, 189.
- Mbong, G.; Fokunang, C.; Manju, E.; Njukeng, A.; Tembe-Fokunang, E.; Hanna, R. Mycelia Growth and Sporulation of Phytophthora colocasiae Isolates under Selected Conditions. Am. J. Exp. Agric. 2015, 8, 193–201.
- Padmaja, G.; Uma Devi, G.; Kanaka Mahalakshmi, B.; Sridevi, D. Characterization of Isolates of Phytophthora colocasiae Collected from Andhra Pradesh and Telangana Causing Leaf Blight of Taro. Int. J. Curr. Microbiol Appl. Sci. 2017, 6, 1901–1912.
- Ooka, J.J. Taro Diseases. Research-Extension-Series. Hawai Inst. Trop. Agricul. Hum. Res. 1990, 114, 51–59.
- Bandyopadhyay, R.; Sharma, K.; Onyeka, T.J.; Aregbesola, A.; Kumar, P.L. First Report of Taro (Colocasia esculenta) Leaf Blight Caused by Phytophthora colocasiae in Nigeria. Plant Dis. 2011, 95, 618.
- Omane, E.; Oduro, K.A.; Cornelius, E.W.; Akrofi, A.Y.; Asare, E.K.; Adongo, B.A. Improvement in the Management of Leaf Blight Disease of Taro (Colocasia esculenta) with Copper and Metalaxyl Fungicides. J. Exp. Agric. Int. 2020, 42, 127–136.
- Onyeka, J. Status of Cocoyam (Colocasia esculenta and Xanthosoma spp.) in West and Central Africa: Production, Household Importance and the Threat from Leaf Blight. Lima (Peru). CGIAR Res. Program Roots Tubers Banan. (RTB) 2014, 39, 1–39.
- Misra, R.S.; Mishra, A.K.; Sharma, K.; Jeeva, M.L.; Hegde, V. Characterisation of Phytophthora colocasiae isolates associated with leaf blight of taro in India. Arch. Phytopathol. Pflanzenschutz 2011, 44, 581–591.
- Jackson, G.V.H.; Gollifer, D.E. Disease and Pest Problems of Taro (Colocasia esculenta L. Schott) in the British Solomon Islands. PANS Pest. Artic. News Summ. 1975, 21, 45–53.
- Indrani Devi, Y.; Sahoo, M.R.; Mandal, J.; Dasgupta, M.; Prakash, N. Correlations between antioxidative enzyme activities and resistance to Phytophthora leaf blight in taro. J. Crop. Improv. 2020, 35, 250–263.
- Singh, D.; Jackson, G.; Hunter, D.; Fullerton, R.; Lebot, V.; Taylor, M.; Iosefa, T.; Okpul, T.; Tyson, J. Taro Leaf Blight—A Threat to Food Security. Agriculture 2012, 2, 182–203.
- Ganança, J.F.T.; Freitas, J.G.F.; Nóbrega, H.G.M.; Rodrigues, V.; Antunes, G.; Rodrigues, M.; Carvalho, M.Â.A.P.; Lebot, V. Screening of Elite and Local Taro (Colocasia esculenta) Cultivars for Drought Tolerance. Procedia Environ. Sci. 2015, 29, 41–42.
- Anjitha Nair, U.M.; Devi, A.A.; Veena, S.S.; Krishnan, B.S.P.; Arya, R.S. Genetic Diversity Analysis of Leaf Blight Resistant and Susceptible Taro Genotypes Using ISSR Markers. J. Root Crops 2018, 44, 44–52.
- Ganança, J.F.T.; Freitas, J.G.R.; Nóbrega, H.G.M.; Rodrigues, V.; Antunes, G.; Gouveia, C.S.S.; Rodrigues, M.; de Carvalho, M.A.P.; Lebot, V. Screening for Drought Tolerance in Thirty Three Taro Cultivars. Not. Bot. Horti. Agrobot. Cluj Napoca 2018, 46, 65–74.
- Bellinger, M.R.; Paudel, R.; Starnes, S.; Kambic, L.; Kantar, M.B.; Wolfgruber, T.; Lamour, K.; Geib, S.; Sim, S.; Miyasaka, S.C.; et al. Taro Genome Assembly and Linkage Map Reveal QTLs for Resistance to Taro Leaf Blight. G3 2020, 10, 13.
- Martin, F.N.; English, J.T. Population genetics of soil-borne fungal plant pathogens. Phytopathology 1997, 87, 446–447.
- Ackah, F.K.; van der Puije, G.C.; Moses, E. First evaluation of taro (Colocasia esculenta) genotypes against leaf blight (Phytophthora colocasiae) in Ghana. HRS 2014, 3, 390–391.
- Amadi, C.O.; Onyeka, J.; Okoye, B.C.; Ezeji, L.; Ezigbo, E.C. Evaluation of Exotic Genotypes of Taro (Colocasia esculenta) in Nigeria. Niger. Agric. J. 2015, 46, 36–42.
- Reddy, P.P. Taro, Colocasia esculenta. In Plant Protection in Tropical Root and Tuber Crops; Springer: New Delhi, India, 2015.
- Kidanemariam, D.B.; Sukal, A.C.; Abraham, A.D.; Stomeo, F.; Dale, J.L.; James, A.P.; Harding, R.M. Identification and molecular characterization of Taro bacilliform virus and Taro bacilliform CH virus from East Africa. Plant Pathol. 2018, 67, 1977–1986.
- Kidanemariam, D.B.; Sukal, A.C.; Abraham, A.D.; Njuguna, J.N.; Stomeo, F.; Dale, J.L.; James, A.P.; Harding, R.M. Incidence of RNA viruses infecting taro and tannia in East Africa and molecular characterisation of dasheen mosaic virus isolates. Ann. Appl. Biol. 2021, 180, 211–223.
- Awuah, R.T. Leafspot of taro (Colocasia esculenta (L.) Schott) in Ghana and suppression of symptom development with Thiophanate methyl. Afr. Crop. Sci. J. 1995, 3, 519–523.
- FAOSTAT. FAO Statistical Database. 2015. Available online: https://www.fao.org/faostat/en/#compare (accessed on 10 February 2022).
- Alexandra, S.; Jamora, N.; Smale, M.; Ghanem, M.E. The tale of taro leaf blight: A global effort to safeguard the genetic diversity of taro in the Pacific. Food Secur. 2020, 12, 1005–1016.
- Food and Agriculture Organization of the United Nations (FAO). The State of Food Security and Nutrition in the World. Building Climate Resilience for Food Security and Nutrition; FAO: Rome, Italy, 2018.
- Mbi, T.K.; Ntsomboh-Ntsefong, G.; Tonfack, L.B.; Youmbi, E. Field management of Taro (Colocasia esculenta (L.) Schott) leaf blight via fungicidal spray of foliage. JCAS 2021, 16, 197–208.
- Chaïr, H.; Traore, R.E.; Duval, M.F.; Rivallan, R.; Mukherjee, A.; Aboagye, L.M.; Van Rensburg, W.J.; Andrianavalona, V.; Pinheiro de Carvalho, M.A.A.; Saborio, F.; et al. Genetic Diversification and Dispersal of Taro (Colocasia esculenta (L.) Schott. PLoS ONE 2016, 11, e0157712.
- Rashmi, D.R.; Raghu, N.; Gopenath, T.S.; Palanisamy, P.; Bakthavatchalam, P.; Karthikeyan, M.; Gnanasekaran, A.; Ranjith, M.S.; Chandrashekrappa, G.K.; Basalingappa, K.M. Taro (Colocasia esculenta): An overview. J. Med. Plants Stud. 2018, 6, 156–161.
- Dansi, A.; Adjatin, A.; Adoukonou-Sagbadja, H.; Faladé, V.; Adomou, A.C.; Yedomonhan, H.; Akpagana, K.; de Foucault, B. Traditional leafy vegetables in Benin: Folk nomenclature, species under threat and domestication. Acta Bot. Gall. 2009, 156, 183–199.
- Dansi, A. Chapter 21: Collecting Vegetatively Propagated Crops (Especially Roots and Tubers); Guarino, L., Ramanatha, R.V., Goldberg, E., Eds.; Bioversity International: Rome, Italy, 2011; p. 8.
- Akplogan, R.M.; Cacaï, G.H.T.; Ahanhanzo, C.; Houédjissin, S.S.; Traoré, E.R. Endogenous Perception of the Diversity of Taro (Colocasia esculenta) Cultivars Produced in Benin. J. Plant. Sci. 2018, 6, 144–148.
- Bammite, D.; Matthews, P.J.; Dagnon, D.Y.; Agbogan, A.; Odah, K.; Dansi, A.; Tozo, K. Agro morphological characterization of taro (Colocasia esculenta) and yautia (Xanthosoma mafaffa) in Togo, West Africa. Afr. J. Agric. Res. 2018, 13, 934–945.
- Boampong, R.; Boateng, S.K.; Adu Amoah, R.; Adu Gyamfi, B.; Aboagye, L.M.; Ansah, E.O. Growth and Yield of Taro (Colocasia esculenta (L) Schott.) as Affected by Planting Distance. Int. J. Agron. 2020, 2020, 1–8.
- Boampong, R.; Aboagye, L.M.; Nyadanu, D.; Esilfie, M. Agro-morphological characterization of some taro (Colocasia esculenta (L.) Schott.) germplasms in Ghana. J. Plant Breed. Crop Sci. 2018, 10, 191–202.
- Orji, K.O.; Ogbonna, P.E. Morphological correlation analysis on some agronomic traits of taro (Colocasia esculenta) in the plains of Nsukka, Nigeria. J. Glo. Biosci. 2015, 4, 1120–1126.
- Mgbedike, N.; Celestine, U.; Josephine, O.; Cletus, A. Strategies for Improving Cocoyam Production among Farmers in Anambra State, Nigeria. 2017. Available online: https://www.semanticscholar.org/paper/Strategies-for-Improving-Cocoyam-Production-among-%2C-Mgbedike-Celestine/78d939e5d33a183b2a31326c9070b856ee0ad289 (accessed on 9 September 2021).
- Jackson, G.V.H.; Pelomo, P.M. Breeding for Resistance to Diseases of Taro, Colocasia esculenta, in Solomon Islands. In International Symposium on Taro and Cocoyam in Baybay Philippines; International Foundation for Science: Stockholm, Sweden, 1980; Volume 5, pp. 287–289.
- Brown, V.M.; Asemota, H. PCR-Based Characterization of Dasheen (Colocasia sp.) and Cocoyam (Xanthosoma sp.). J. Biotech. Res. 2009, 1, 28–40.
- Ivancic, A.; Garcia, J.Q.; Lebot, V. Development of visual tools for selecting qualitative corm characteristics of taro (Colocasia esculenta (L.) Schott). Aust. J. Agric. Res. 2003, 54, 581–587.
- Cho, J.J. Breeding Hawaiian taros for the future. In Proceedings of the Third International Taro Symposium, Nadi, Fiji Islands, 21–23 May 2003.
- Quero-García, J.; Letourmy, P.; Ivancic, A.; Feldmann, P.; Courtois, B.; Noyer, J.L.; Lebot, V. Hybrid performance in taro (Colocasia esculenta) in relation to genetic dissimilarity of parents. Theor. Appl. Genet. 2009, 119, 213–221.
- Amadi, C.O.; Mbanaso, E.N.A.; Chukwu, G.O. A Review Cocoyam Breeding in Nigeria: Achievements, Challenges and Prospects. Niger. Agric. J. 2012, 43, 72–80.
- Singh, D.; Hunter, D.; Iosefa, T.; Okpul, T.; Fonoti, P.; Delp, C. Improving taro production in the South Pacific through breeding and selection. In The Global Diversity of Taro: Ethnobotany and Conservation; Ramanatha, R.V., Matthews, P.J., Eyzaguirre, P.B., Hunter, D., Eds.; Bioversity International: Rome, Italy, 2010; p. 168.
- TaroGen. AusAID/SPC Taro Genetic Resources: Conservation and Utilisation Project; Annual Report 1998/1990; Secretariat of the Pacific Community: Noumea, New Caledonia, 1999.
- Singh, D.; Hunter, D.; Okpul, T.; Iosefa, T. Introduction to techniques and methods of taro breeding. In Proceedings of the AusAID/SPC Taro Genetic Resources: Conservation and Utilization, Taro Pathology and Breeding Workshop Taro Pathology and Breeding Workshop, Alafua Campus, Samoa, 5–7 November 2001; pp. 38–41.
- Wilson, J.E. Taro Breeding. AgroFacts Crop, No. 3/89; IRETA Publications: Pittsburgh, PA, USA, 1990.
- Tuia, V.S. In Vitro Multiplication of Taro (Colocasia esculenta var. esculenta L. Schott). Ph.D. Thesis, University of the South Pacific, Suva, Fiji, 1997.
- Thinh, N.T. Cryopreservation of Germplasm of Vegetatively Propagated Tropical Monocots by Vitrification. Ph.D. Thesis, Kobe University, Kobe, Hyogo, Japan, 1997.
- Verma, V.M.; Cho, J.J.; Aikne, J.; David, J. High frequency plant production of taro (Colocasia esculenta (L.) Schott) by tissue culture. In Proceedings of the 4th International Crop Science Congress, Brisbane, Australia, 26 September 2004.
- Deo, P.C.; Harding, R.M.; Taylor, M.; Tyagi, A.P.; Becker, D.K. Somatic embryogenesis, organogenesis and plant regeneration in taro (Colocasia esculenta var. esculenta). PCTOC 2009, 99, 61–71.
- Fukino, I.T.O.N.; Hanada, K.; Ajisaka, H.; Sakai, J.; Hirochika, H.; Hirai, M.; Hagio, T.; Enomoto, S. Transformation of Taro (Colocasia esculenta Schott) using particle bombardment. Jpn. Agric. Res. Q. 2000, 34, 159–165.
- He, X.; Miyasaka, S.; Fitch, M.M.; Zhu, Y.J.; Moore, P.H. Transformation of taro (Colocasia esculenta) with a rice chitinase gene (Abstract). Vitr. Cell Dev. Biol. Plant 2004, 40, 2104.
- He, X.; Miyasaka, S.C.; Fitch, M.M.; Moore, P.H.; Zhu, Y.J. Agrobacterium tumefaciens-mediated transformation of taro (Colocasia esculenta (L.) Schott) with a rice chitinase gene for improved tolerance to a fungal pathogen Sclerotium rolfsii. Plant Cell Rep. 2008, 27, 903–909.
- Deo, P.C. Somatic Embryogenesis and Transformation in Taro (Colocasia esculenta var. esculenta). Ph.D. Thesis, The University of the South Pacific, Suva, Fiji, 2008.