Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by Andrey Elchaninov.
Macrophages are key participants in the maintenance of tissue homeostasis under normal and pathological conditions, and implement a rich diversity of functions. The largest population of resident tissue macrophages is found in the liver. Hepatic macrophages, termed Kupffer cells, are involved in the regulation of multiple liver functionalities. Kupffer cells (KCs), the resident liver macrophages, constitute a crucially important component of the mononuclear-monocytic system. KCs have a wide variety of responsibilities at both local and systemic level, notably the barrier function preventing various pathogens and their toxic by-products (e.g., endotoxin, also known as bacterial lipopolysaccharide (LPS)) from entering systemic circulation.
- macrophages
- monocytes
- macrophage niche
- Kupffer cells
1. Introduction
Macrophages participate in various physiological, immunological and morphogenetic processes including regeneration. Over 95% of all macrophages found in mammalian body are concentrated in the liver, which harbors the most abundant of the resident tissue macrophage populations [1]. Kupffer cells (KCs), the resident liver macrophages, constitute a crucially important component of the mononuclear-monocytic system. KCs have a wide variety of responsibilities at both local and systemic level, notably the barrier function preventing various pathogens and their toxic by-products (e.g., endotoxin, also known as bacterial lipopolysaccharide (LPS)) from entering systemic circulation [2]. KCs constitute estimated 30–35% of the total non-parenchymal liver cell counts [3] and are typically located within the lumina of sinusoidal capillaries adherent to the endothelium, which provides them with immediate access to immunogenic foreign agents that arrive with portal circulation [4]. KCs also participate in protein and lipid metabolism, as well as the clearance of apoptotic cells from circulation [5]. KC dysfunctions have been associated with a number of liver diseases, e.g., viral hepatitis, cholestasis, alcoholic cirrhosis and fibrosis [6].
Some studies use CD11b as a marker of bone marrow-derived (monocytic) macrophages [43,65][14][37]. Importantly, although CD11b is expressed by both KCs and monocytic macrophages of the liver, monocytic macrophages express it at higher levels than KCs [59,60][31][32].
A remarkable series of recent studies introduce a special macrophage population inhabiting the connective tissue capsule of the liver [62,63][34][35]. It is reasonable to assume that KCs associated with sinusoidal capillaries eliminate pathogens that have already entered the blood, but obviously cannot interfere with the spread of pathogens in the abdominal cavity. The reseauthorchers describe a population of macrophages within the liver capsule that differ from KCs both phenotypically and by origin, which probably participate in the intra-abdominal clearance, showing that up to 3 × 105 macrophages inhabit a single mouse liver capsule [62,63][34][35]. The liver capsule macrophages (LCMs) develop from bone marrow precursors and have very long processes (a morphological distinction). The data indicate that LCMs express F4/80 as well as other macrophage markers such as CD64, CSF-1R, CX3CR1 and CD14. At the same time, they express CD11c at low levels and express neither CD103 nor Tim4. The main function of LCMs is to defend the liver from microbial pathogens [63][35], partly by stimulating the neutrophil infiltration of the liver. Although LCMs express CD207, similar CD207+ macrophages can be observed in the vicinity of central veins in liver lobules [40,63][8][35].
Another recently described liver macrophage population is associated with bile ducts and is positive for the Gpnmb marker, which identifies them as lipid-associated macrophages (LAMs) apparently derived from recruited monocytes [41][9]. LAMs are less sensitive to endotoxin/LPS than KCs, which may be due to their affiliation with the branching portal vein delivering LPS and other pathogen-related products to the liver, and so LAMs may develop tolerance to these agents [41][9]; there is no decisive evidence on this subject as yet.
The enhanced heterogeneity of liver macrophages under pathological conditions reflects both the immigration of monocytic macrophages and the phenotypic alterations in KCs.
Murine model of steatohepatitis harbors two major populations of liver macrophages: KCs expressing Clec4f and monocytic macrophages expressing Lyz2 [66][38]; at that, the population of monocytic macrophages is heterogeneous in itself. The reseauthorchers identify at least three cell subtypes within monocytic macrophage population: (I) expressing high levels of Fn1, Mgst1 and Msrb1; (II) expressing high levels of Chil1; and (III) expressing Il1b [66][38].
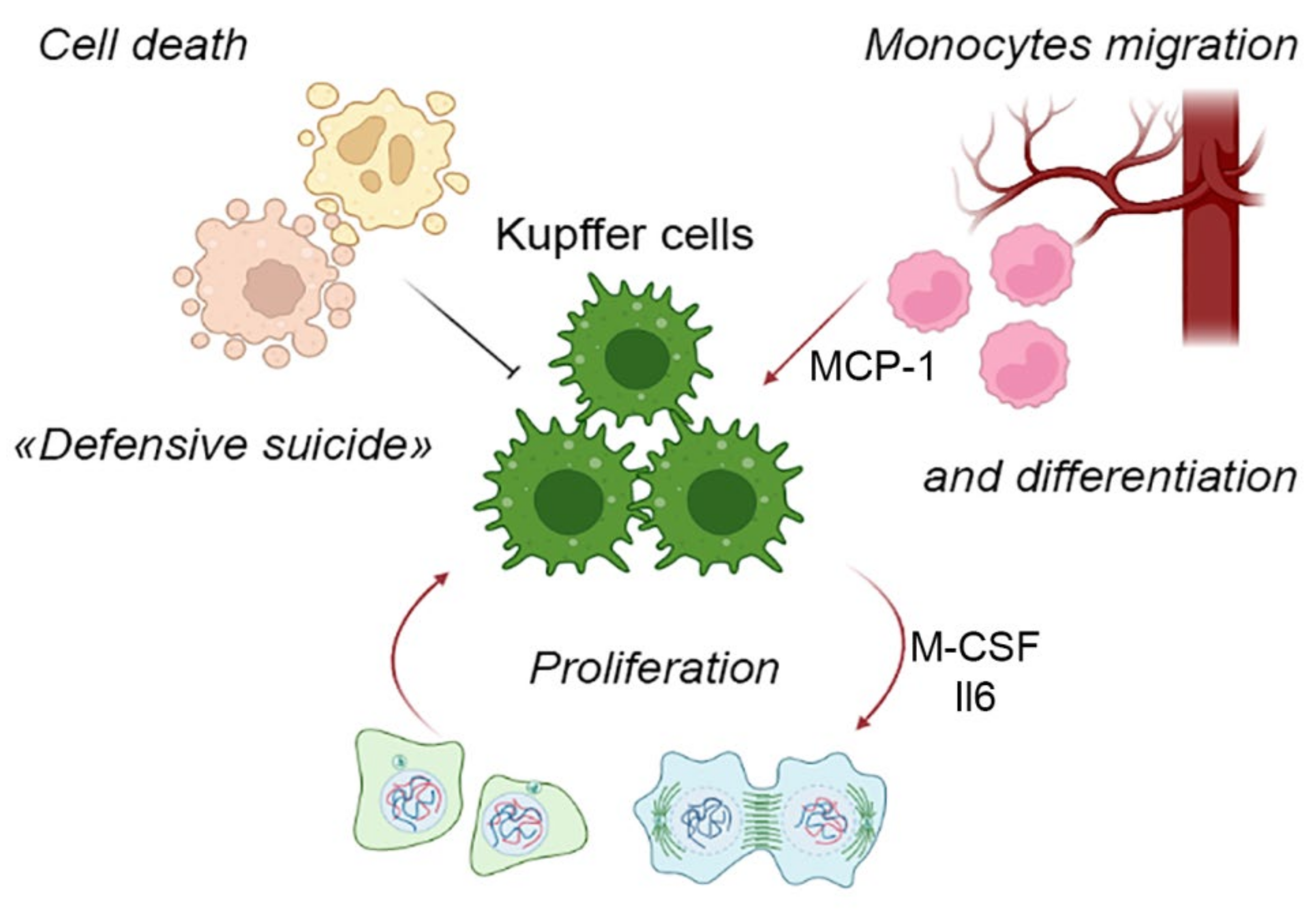 The remarkable proliferative capacity of liver macrophages, most unexpected by the researchers, turned out to be virtually the only route of the liver macrophage population maintenance under physiological conditions and even in certain pathologies [68][40].
It should be noted that in some studies, even after non-genotoxic depletion of local macrophages in lungs and red bone marrow, their populations are successfully replenished by means of resident cell proliferation with minimal participation of bone marrow-derived monocytic precursors. However, with the temporary block of the local macrophage proliferation (e.g., by applying a lethal dose of ionizing radiation), the population is restored by recruitment of circulating monocytes [70,71][42][43]. Macrophage proliferation is presumably stimulated by IL-4 acting independently of the cell origin (monocytic or resident) [68,72][40][44].
The proliferative capacity specifically of KCs has been demonstrated experimentally in various models, including resections of different volume and acute liver injury [44,73][15][45]. For instance, after 70% liver mass resection in mice, up to 50% of macrophages within the liver remnant enter proliferation [74][46]. Importantly, for Kupffer cells, the main proliferation-driving cytokine is not IL-4, but IL-6 [75][47], which indicates some organ-specificity of mitotic cycle regulation mechanisms in macrophages. For lung macrophages and some other macrophage populations, proliferation has been shown to depend on M-CSF and GM-CSF as well [70][42].
The proliferative capacity of KCs has clinical significance. For instance, in non-alcoholic steatohepatitis, the mitotic capacity of KCs is blocked, and their population is replaced with monocytic macrophages that gradually acquire KC-like phenotypes. However, these ‘new’ macrophages have pro-inflammatory phenotypes and metabolize triglycerides less efficiently, which eventually leads to aggravation of the condition [76][48].
Another component process affecting macrophage numbers in the liver is migration of blood monocytes to the liver and their subsequent differentiation into macrophages. This component is most pronounced under conditions of toxic liver injury modeled in laboratory animals with the use of carbon tetrachloride or acetaminophen (paracetamol). The numbers of immigrating monocytes in the liver peaked 24 h after acetaminophen-induced damage; the cells differentiated into macrophages and were subsequently eliminated from the liver [44][15].
The immigration of monocytes/macrophages, albeit at a much lower scale, can be observed during regeneration of the liver after 70% resection in mice. An increase in Ly6C+ and CD11b+ cell numbers within the liver as early as 24 h after resection was demonstrated. Notably, the counts of Ly6C+ cells continue to increase until day 7 post-resection, amid a decline in CD11b+ cell counts [74][46]. The extent of monocyte/macrophage migration to the remnant liver depends on both the resection volume and model animal species. For example, resection of more than 80% liver volume in rats promotes negligible migration of monocytic macrophages to the remnant liver [73][45].
It has long been held that macrophages immigrating to the liver are totally derived from blood monocytes. This long-standing opinion was challenged by recent findings obtained in various in vivo settings, including hepatotoxicity models and 70% liver resections in mice.
Apart from the blood monocytes, an alternative source of macrophages immigrating to the liver can be provided by peritoneal macrophages [64][36]. This phenomenon was discovered in a model of localized sterile heat injury of the liver: macrophages migrating to the area of damage expressed classical markers but also CD102 and GATA6 indicating their belonging to the peritoneal macrophage population. Experimental findings by another group of reseauthorchers indicate that this population does not penetrate deep into the liver and is not involved in the inflammatory and repair processes [77][49]. It should be noted that macrophages with F4/80+Ly6C+CD11b+ phenotype, corresponding to peritoneal macrophages, also appear in the liver after resections, but their origin remains unexplored [64,74,78][36][46][50].
One of the main uncertainties concerning liver macrophage populations under pathological conditions is their further destiny. Several studies show that after the completion of repair processes in the liver monocytic macrophages become eliminated, whereas the resident macrophage numbers are restored by means of resident macrophage proliferation [44[15][22],50], and such scenario has been demonstrated for inflammatory damage in other organs as well [79][51]. However, with depletion of KCs from the liver, the niche is filled by immigrating monocytic macrophages which successfully engage in the long-term maintenance by proliferation [61[33][52],80], although in other organs recolonization proceeds differently for reasons as yet unexplained [70][42].
Moreover, the degree of identity of the colonizing monocytic derivatives to KC is rather controversial. Some studies demonstrate full correspondence between the two lineages, both molecular (gene expression profiles) and functional (proliferation and phagocytosis capacities) [61][33]. Other studies show that the differences persist: for instance, KC express Tim4 and Marco at higher levels and engulf acetylated low density lipoprotein with higher intensity, while showing lower rates of phagocytosis towards bacterial pathogens, as compared to the substitute bone marrow-derived macrophages [81][53]. The controversy possibly results from different observation lengths used in the studies and can be interpreted as follows: the longer bone marrow-derived macrophages stay in the liver (depleted of KC) the closer they come to resemble KC phenotypically and functionally [39][7]. This issue will be given additional consideration in an upcoming section on the macrophage niche of the liver.
Cell death is yet another component process affecting liver macrophage ‘demography’ and the mechanisms of its regulation are poorly understood. Relatively few studies have been focused on this issue, although many studies feature the massive death of macrophages within the framework of primary tissue response to damage, and the liver is no exception. The death of resident macrophages qualified as necroptosis or necrosis is typical for bacterial and viral infections, as well as malaria [82,83,84][54][55][56]. This phenomenon, described for the first time in alveolar macrophages and termed ‘defensive suicide’, essentially triggers the defensive inflammatory reaction and is by no means a passive casualty of the invading microbial pathogen [83,85][55][57]. Increased rates of cell death in liver macrophages have been described in various murine models (hepatotoxic injury, 70% liver resection), with the scale of cell death among F4/80+ liver macrophages reaching 16% [74][46]. The scale, functional significance and molecular mechanisms of macrophage cell death during tissue repair, particularly in the liver, remain understudied. Clearly, the death of resident macrophages may represent a part of tissue response, acting as a trigger for repair processes.
The remarkable proliferative capacity of liver macrophages, most unexpected by the researchers, turned out to be virtually the only route of the liver macrophage population maintenance under physiological conditions and even in certain pathologies [68][40].
It should be noted that in some studies, even after non-genotoxic depletion of local macrophages in lungs and red bone marrow, their populations are successfully replenished by means of resident cell proliferation with minimal participation of bone marrow-derived monocytic precursors. However, with the temporary block of the local macrophage proliferation (e.g., by applying a lethal dose of ionizing radiation), the population is restored by recruitment of circulating monocytes [70,71][42][43]. Macrophage proliferation is presumably stimulated by IL-4 acting independently of the cell origin (monocytic or resident) [68,72][40][44].
The proliferative capacity specifically of KCs has been demonstrated experimentally in various models, including resections of different volume and acute liver injury [44,73][15][45]. For instance, after 70% liver mass resection in mice, up to 50% of macrophages within the liver remnant enter proliferation [74][46]. Importantly, for Kupffer cells, the main proliferation-driving cytokine is not IL-4, but IL-6 [75][47], which indicates some organ-specificity of mitotic cycle regulation mechanisms in macrophages. For lung macrophages and some other macrophage populations, proliferation has been shown to depend on M-CSF and GM-CSF as well [70][42].
The proliferative capacity of KCs has clinical significance. For instance, in non-alcoholic steatohepatitis, the mitotic capacity of KCs is blocked, and their population is replaced with monocytic macrophages that gradually acquire KC-like phenotypes. However, these ‘new’ macrophages have pro-inflammatory phenotypes and metabolize triglycerides less efficiently, which eventually leads to aggravation of the condition [76][48].
Another component process affecting macrophage numbers in the liver is migration of blood monocytes to the liver and their subsequent differentiation into macrophages. This component is most pronounced under conditions of toxic liver injury modeled in laboratory animals with the use of carbon tetrachloride or acetaminophen (paracetamol). The numbers of immigrating monocytes in the liver peaked 24 h after acetaminophen-induced damage; the cells differentiated into macrophages and were subsequently eliminated from the liver [44][15].
The immigration of monocytes/macrophages, albeit at a much lower scale, can be observed during regeneration of the liver after 70% resection in mice. An increase in Ly6C+ and CD11b+ cell numbers within the liver as early as 24 h after resection was demonstrated. Notably, the counts of Ly6C+ cells continue to increase until day 7 post-resection, amid a decline in CD11b+ cell counts [74][46]. The extent of monocyte/macrophage migration to the remnant liver depends on both the resection volume and model animal species. For example, resection of more than 80% liver volume in rats promotes negligible migration of monocytic macrophages to the remnant liver [73][45].
It has long been held that macrophages immigrating to the liver are totally derived from blood monocytes. This long-standing opinion was challenged by recent findings obtained in various in vivo settings, including hepatotoxicity models and 70% liver resections in mice.
Apart from the blood monocytes, an alternative source of macrophages immigrating to the liver can be provided by peritoneal macrophages [64][36]. This phenomenon was discovered in a model of localized sterile heat injury of the liver: macrophages migrating to the area of damage expressed classical markers but also CD102 and GATA6 indicating their belonging to the peritoneal macrophage population. Experimental findings by another group of reseauthorchers indicate that this population does not penetrate deep into the liver and is not involved in the inflammatory and repair processes [77][49]. It should be noted that macrophages with F4/80+Ly6C+CD11b+ phenotype, corresponding to peritoneal macrophages, also appear in the liver after resections, but their origin remains unexplored [64,74,78][36][46][50].
One of the main uncertainties concerning liver macrophage populations under pathological conditions is their further destiny. Several studies show that after the completion of repair processes in the liver monocytic macrophages become eliminated, whereas the resident macrophage numbers are restored by means of resident macrophage proliferation [44[15][22],50], and such scenario has been demonstrated for inflammatory damage in other organs as well [79][51]. However, with depletion of KCs from the liver, the niche is filled by immigrating monocytic macrophages which successfully engage in the long-term maintenance by proliferation [61[33][52],80], although in other organs recolonization proceeds differently for reasons as yet unexplained [70][42].
Moreover, the degree of identity of the colonizing monocytic derivatives to KC is rather controversial. Some studies demonstrate full correspondence between the two lineages, both molecular (gene expression profiles) and functional (proliferation and phagocytosis capacities) [61][33]. Other studies show that the differences persist: for instance, KC express Tim4 and Marco at higher levels and engulf acetylated low density lipoprotein with higher intensity, while showing lower rates of phagocytosis towards bacterial pathogens, as compared to the substitute bone marrow-derived macrophages [81][53]. The controversy possibly results from different observation lengths used in the studies and can be interpreted as follows: the longer bone marrow-derived macrophages stay in the liver (depleted of KC) the closer they come to resemble KC phenotypically and functionally [39][7]. This issue will be given additional consideration in an upcoming section on the macrophage niche of the liver.
Cell death is yet another component process affecting liver macrophage ‘demography’ and the mechanisms of its regulation are poorly understood. Relatively few studies have been focused on this issue, although many studies feature the massive death of macrophages within the framework of primary tissue response to damage, and the liver is no exception. The death of resident macrophages qualified as necroptosis or necrosis is typical for bacterial and viral infections, as well as malaria [82,83,84][54][55][56]. This phenomenon, described for the first time in alveolar macrophages and termed ‘defensive suicide’, essentially triggers the defensive inflammatory reaction and is by no means a passive casualty of the invading microbial pathogen [83,85][55][57]. Increased rates of cell death in liver macrophages have been described in various murine models (hepatotoxic injury, 70% liver resection), with the scale of cell death among F4/80+ liver macrophages reaching 16% [74][46]. The scale, functional significance and molecular mechanisms of macrophage cell death during tissue repair, particularly in the liver, remain understudied. Clearly, the death of resident macrophages may represent a part of tissue response, acting as a trigger for repair processes.
 Another consideration to be added to the fundamental macrophage niche concept is a new parameter denoted as “time of residence” within the organ. Introduction of this parameter was necessitated by apparent controversy of experimental findings—success or failure to distinguish between true resident macrophages and those differentiated from the arriving blood monocytes depending on particular experimental setting [61,81][33][53]. Given that in many organs resident macrophage populations are gradually replaced by bone marrow-derived cells, such mixed populations are significantly heterogeneous in terms of time since colonization, which can be also defined as the niche occupation length. Expression levels of TIMD4 (TIM4) protein have been shown to correlate with the length of macrophage lineage affiliation with its current place of residence (niche). For instance, Scott et al. (2016) observed rapid (in the course of several days) acquisition of KC-like phenotypes by monocytes arriving in the liver; at that, Timd4 expression in these new KC-like monocytic macrophages stayed negligible and its induction was delayed for over a month [61][33]. These data are consistent with the results of studies on liver macrophage dynamics in non-alcoholic steatohepatitis murine model [41][9].
To take stock, the process of full differentiation of the newly arrived monocytes into resident liver macrophages consists of two stages. At the first of them, a vacant macrophage niche generates ‘stay here’ signals to facilitate quick adaptation; the second stage, assimilation, is lengthier: the macrophage becomes fully integrated in the niche, receiving support in the form of ‘learn this’ signals [80,111][52][83].
Despite the opinion that monocytes can be differentiated in any type of tissue macrophages and exactly mimic any resident macrophage lineage except CNS microglia [105][77], monocyte-derived mouse liver macrophages still differed by their expression profiles from KCs as late as 6 weeks post-colonization [81][53], and a similar delay in leukocyte differentiation apparently occurs in the human liver [112][84].
To what extent does TIM4 expression reflect the length of stay and the depth of monocytic macrophage assimilation in the macrophage niche of KCs? In ouresearchers' opinion, this is a complex issue. OurResearchers own data obtained in a model of 70% liver resection in mice indicate that Tim4 expression in hepatic tissues can rapidly increase in the remnant liver tissue. This increase can be attributed to the influence of boosted endotoxin/LPS blood levels, and this suggestion has been supported by in vitro experiments [113][85]. Notably, KCs isolated from the intact liver by magnetic sorting for F4/80 had Tim4 expression levels similar to macrophages differentiated from peripheral blood monocytes isolated by magnetic sorting for CD115 and cultured with M-CSF. Under LPS exposure, Tim4 expression was significantly upregulated in both types of cultures. As it was also demonstrated, after liver resection in mice Ly6C+ monocytes migrate to it in high numbers. Considering the reduced Tim4 expression in the ‘newcomers’ [61][33], it would be reasonable to expect decreased expression of TIM4 marker in total liver macrophages after resection. However, this proved not to be the case, apparently due to the stimulating effect of LPS on the new, differentiating monocytic macrophages [113][85]. It can be concluded that, in this setting, Tim4 expression levels depend more on LPS exposure than on the length of residence in the macrophage niche of KCs [113][85].
Similar considerations and findings are applicable to another marker of resident liver macrophages, MARCO [81][53]. The use of this marker for distinguishing between macrophages arising from different sources has been based on the assumption that its expression is relatively constant and does not respond to endotoxin/LPS [81][53]. However, the latter point is questionable, considering the long-known role of this receptor in antimicrobial immunity [114,115][86][87]. True enough, KCs and bone marrow-derived macrophages express Marco at different levels; however, a sharp increase in Marco expression at both mRNA and protein levels in the liver remnant was observed after 70% hepatectomy, and this effect was reproduced by in vitro exposure of liver macrophage cultures to LPS [113][85]. Such dynamics of MARCO expression are consistent with its being a marker of pro-inflammatory state in murine macrophages [103][75].
The endotoxin/LPS sensitivity of the candidate macrophage markers is essential, as the vast portion of LPS in mammalian body is metabolized by the liver, not to mention the role of any tissue macrophages in antibacterial defense [116][88]. These considerations further implicate the tissue inflammatory status as a decisive factor which determines the phenotypical and functional properties of macrophages. The impact of inflammatory status on macrophage functionalities is especially prominent in the liver, given the barrier function of the organ. It has been already mentioned that various infections can promote a wave of cell death in resident macrophages, considered a ‘defensive suicide’ [85][57]. It is important to note that infectious lesions, which become sites of massive death of resident macrophages, are rapidly colonized by macrophages differentiating from the migratory blood monocytes. Notably, during influenza A infections, the resident alveolar macrophages become replaced by monocyte-derived macrophages, which appear to be more efficient in fighting Streptococcus pneumoniae infections due to higher production levels of IL6, CCL3, CCL4 and G-CSF [117,118][89][90]. Similarly, after a herpesvirus infection, alveolar macrophages effectively prevent the development of asthma by virtue of replacement of the ‘old’ resident alveolar macrophages by ‘new’ regulatory monocyte-derived macrophages that block the ability of dendritic cells to trigger Th2 responses [119][91].
A team of reseauthorchers observing an extraordinary robust immune response to S. pneumoniae following adenoviral infection attributed it to a previously unidentified ‘macrophage memory’ phenomenon, thereby suggesting the existence of memory cells in macrophage lineages [117,118][89][90]. In essence, the adenoviral infection causes activation of the resident alveolar macrophages, thus stimulating formation of a special self-perpetuating alveolar macrophage population. Upon activation, the naïve alveolar memory macrophages memorize the microenvironmental cues under the influence of Th cells and IFN-γ released by them [118][90]. The identified memory macrophage population is considered self-perpetuating and independent of blood monocytes. These cells are believed to retrieve the memorized information on previous inflammatory reactions to microbial pathogens in case of new infections; their action involves recruitment of neutrophils to the inflammatory foci [118][90].
The monocyte-derived macrophages which colonize the liver in the aftermath of KC depletion provide more efficient clearance of Neisseria meningitidis or Listeria monocytogenes by phagocytosis compared with KCs [81][53]; they also exert a more pronounced pro-inflammatory effect [76][48]. OuResearchers own in vitro experiments have demonstrated that, at early time points of stimulation, monocytic macrophages engulf latex particles at higher rates compared with KCs [87,120][59][92]. Such data can be interpreted in terms of hepatic tissue macrophage niche and its special features.
Over the entire history of research on the mononuclear phagocyte system in mammals macrophages were considered as cells with pronounced phenotypical plasticity confirmed in numerous studies [103][75]. Comparative evaluation of resident macrophages vs blood monocyte-derived macrophages shows higher sensitivity of the latter to activating factors and their higher phenotypic plasticity. Presumably, the prolonged exposure of resident macrophages to the conditions of organ-specific tissue niche leads to a reduction in plasticity through epigenetic block of inflammation-related genes. Such suppression is beneficial, as it suits the needs of the organ homeostasis. This view is based on research involving alveolar macrophages [79][51] and its validity for other resident macrophage populations, including those of the liver, has not been verified so far.
The liver has long been considered an immunotolerogenic organ [4]. Maintenance of this capacity is largely a responsibility and merit of KCs. Under normal conditions, KCs produce PD-L1, which participates in suppression of cellular immunity reactions, and small amounts of TNFa and IL-12. Under stimulation, KCs produce both pro-and anti-inflammatory cytokines [60[32][93],121], but their responses to many pathogen-associated molecular patterns (PAMPs), notably those of endotoxin/LPS are remarkably low. One of the probable reasons is the continuous exposure of KCs to LPS, the concentration of which in the portal blood flow varies within 0.1–1 ng/mL [116][88]. Such exposure presumably endows KCs with LPS tolerance, or at least reduced LPS sensitivity, compared with blood monocytes and monocyte-derived macrophages. OuResearchers own data agree with this assumption. For example, Tlr4 expression in peripheral blood monocytes is significantly higher compared with KCs [122[94][95],123], which is consistent with the evidence on more facile and LPS-sensitive induction of synthesis of certain interleukins in monocytic macrophages [87][59]. At the same time, studying the expression of LPS tolerance-associated genes in KCs and monocytic macrophages, no classical signatures of LPS tolerance were found. Still, KCs revealed lower expression of MAPK signaling-related genes Erk2 and p38 [124][96], known to participate in pro-inflammatory cytokine synthesis and release by macrophages. Reduced production of cytokines by macrophages has been associated with tolerance [125,126][97][98].
It can be assumed that similar mechanisms can reduce KC tolerance to PAMPs other than endotoxin/LPS: for instance, higher expression of Tlr2, Tlr7 and Tlr8 genes in monocytes compared with KCs was observed [122][94]. It should be also noted that the benefit of re-colonization with monocytic macrophages with regard to prevention of asthma and resistance to bacterial pathogens observed in the lungs cannot be straightforwardly extrapolated to the liver. A similar replacement scenario applied to KCs in the liver may have a deeply damaging effect and result in a chronic inflammatory process in hepatic tissues and at systemic level [76,90][48][62].
The introduction of the tissue inflammatory status as one of the parameters ‘in charge’ of the counts and properties of macrophages in particular organ may also ease the apparent controversy concerning the cell fate of monocytes colonizing the liver after toxic injury. As has been mentioned, these monocytes colonize the injured liver to become totally eliminated later on, despite a sharp decline of the resident macrophage populations and the overwhelming abundance of vacant niches. Presumably, the inflammatory status, and notably the time window of elevated TNFa and IL-1 levels determine the permission for monocytes to occupy the vacant niches previously occupied by KCs [80,82][52][54]. Still, even under this assumption, it is difficult to explain subsequent disappearance of these new monocytic lineages after resolution of the inflammatory process, especially given that at early stages of recovery the immigrating monocytes outweigh the preserved KCs numerically. For the reasons as yet unknown, in the toxically injured liver, surviving KCs clearly outcompete the immigrating monocytes/macrophages in settling the vacant niches. Incidentally, despite the just-experienced toxic shock, resident macrophages enter proliferation much earlier. The comparative dynamics imply that to make the monocytic impact visible, 80% of the resident liver macrophages should be depleted, which is hardly possible to achieve with available experimental techniques [80,105][52][77].
Notably, in the rat model of subtotal live resection, expression of Tnfa and Il1 increased significantly by the end of regeneration only, whereas the content of TNFa protein in the remnant was reduced from the beginning and stayed low since. This observation can be related to the lack of immigration of CX3CR1+ macrophages to the remnant liver in this model [73,127][45][99].
With these important amendments to the fundamental macrophage niche concept, the macrophage populations of individual organs are perceived as complex systems engaged in specific functionalities depending on their localization and ensuring the communication among different compartments inside each organ [105][77]. Moreover, macrophages have been implicated in the inter-organ crosstalk as well [39][7]. For example, myocardial overload leads to activation of the sympathetic innervation of the kidneys, resulting in enhanced secretion of S100A8/A9 peptides by the collecting duct epithelium. These peptides stimulate kidney macrophages to release TNFa, which promotes secretion of GM-CSF by endothelial cells in the interstitium. In return, the increased blood levels of GM-CSF promote accumulation of Ly6Clo macrophages in the myocardium; these macrophages produce amphiregulin, which causes hypertrophy of cardiomyocytes [128][100]. Similar data on the relationship between macrophages of the heart, lungs and kidneys were obtained in a model of myocardial infarction [129][101].
This new paradigm provides an unexpected explanation to the experimentally observed increased expression of certain interleukins and growth factors in lungs and kidneys after subtotal hepatectomy in rats [127,130][99][102]. The effect is accompanied by an increase in CD68+ macrophage counts in the lungs [131][103]. The coherence between monocyte/macrophage populations of the spleen and the liver is more comprehensible given the anatomical connection between the two organs via portal circulation [132,133][104][105]. At the same time, the spleen has been implicated as a monocyte supply for other organs as well. For instance, monocytes deposited in the spleen have been shown to migrate to inflammation foci in myocardial infarction and cerebral ischemic stroke [38,134,135][106][107][108].
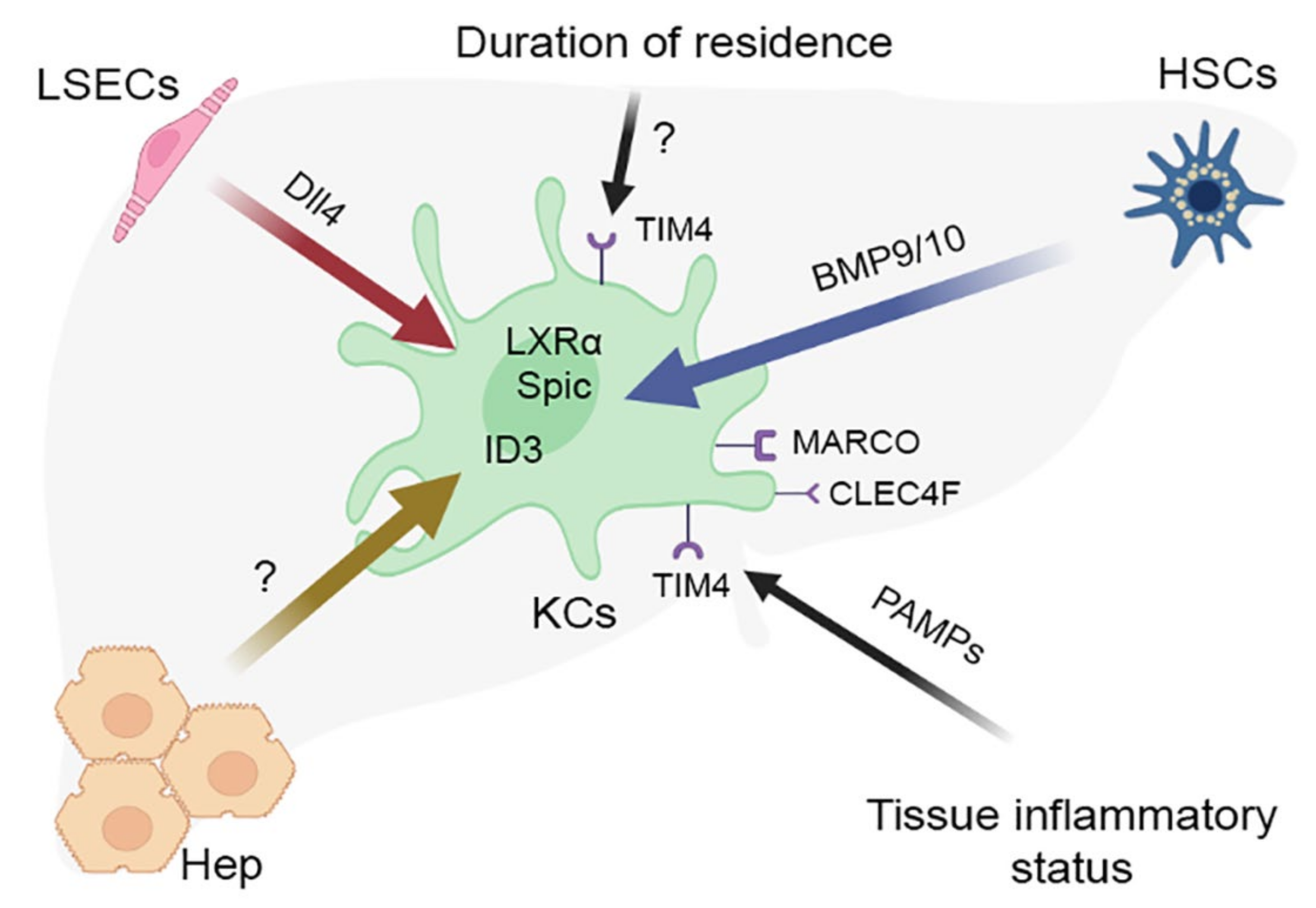
Thus, the accumulated body of evidence suggests a general scheme of acquisition of unique properties by a tissue macrophage population. The first step involves implementation of the core macrophage differentiation program represented by PU.1, MYB, C-MAF, MAFB and ZEB2 transcription factors [136][109]. This universal basis becomes subsequently adjusted and refined by the influence of particular tissue niche, which shapes a transcription program characteristic of particular type of resident macrophages [39][7]. For KCs, the tissue niche comprises Ito cells, sinusoidal endothelial cells and hepatocytes, as well as various ECM components of hepatic parenchyma and paracrine factors; the joint influence of these components shapes a specific transcription program involving the LXR-α/ID3/SPIC expression signature in macrophages [80,111][52][83]. Interestingly, the transcriptional program of Kupffer cells turned out to be similar in mammals (humans, mice, pigs, hamsters and macaques), chicken and zebrafish [40][8], whereas induction of most genes unique to macrophages in these animal species required the interaction of activin receptor-like kinase (ALK1) on Kupffer cells with BMP9/10 secreted by Ito cells, and was also more or less TGFb dependent [40,111][8][83].
The concept of a macrophage niche is consistent with data on the epigenetic regulation of the resident macrophages phenotype, including those of the liver [137,138][110][111]. It has already been mentioned that the expression of the Clec4f gene, which encodes a lectin required for the presentation of alpha-galactosylceramide to natural killer T, is specific for the liver [99][71]. Assessment of histone modification status showed the presence of unique poised and active enhancers in the region of the Clec4f gene, as well as open chromatin regions in the region of the transcription factor LXRa, specific for Kupffer cells [138][111]. Histone acetylation sites were also found in the region of LXR gene, as well as in RBPJ gene, which is consistent with studies that established the dependence of the formation of a specific transcriptional program of Kupffer cells on the NOTCH-ligand DLL4 secreted by liver sinusoid endotheliocytes [111][83]. The effect of LPS on histone acetylation in the area of DNA regulatory regions during the differentiation of monocytes into macrophages has been also shown, which, given the constant contact with LPS, is especially important for Kupffer cells [139][112]. However, more detailed studies in this respect of Kupffer cells have not been conducted.
Another consideration to be added to the fundamental macrophage niche concept is a new parameter denoted as “time of residence” within the organ. Introduction of this parameter was necessitated by apparent controversy of experimental findings—success or failure to distinguish between true resident macrophages and those differentiated from the arriving blood monocytes depending on particular experimental setting [61,81][33][53]. Given that in many organs resident macrophage populations are gradually replaced by bone marrow-derived cells, such mixed populations are significantly heterogeneous in terms of time since colonization, which can be also defined as the niche occupation length. Expression levels of TIMD4 (TIM4) protein have been shown to correlate with the length of macrophage lineage affiliation with its current place of residence (niche). For instance, Scott et al. (2016) observed rapid (in the course of several days) acquisition of KC-like phenotypes by monocytes arriving in the liver; at that, Timd4 expression in these new KC-like monocytic macrophages stayed negligible and its induction was delayed for over a month [61][33]. These data are consistent with the results of studies on liver macrophage dynamics in non-alcoholic steatohepatitis murine model [41][9].
To take stock, the process of full differentiation of the newly arrived monocytes into resident liver macrophages consists of two stages. At the first of them, a vacant macrophage niche generates ‘stay here’ signals to facilitate quick adaptation; the second stage, assimilation, is lengthier: the macrophage becomes fully integrated in the niche, receiving support in the form of ‘learn this’ signals [80,111][52][83].
Despite the opinion that monocytes can be differentiated in any type of tissue macrophages and exactly mimic any resident macrophage lineage except CNS microglia [105][77], monocyte-derived mouse liver macrophages still differed by their expression profiles from KCs as late as 6 weeks post-colonization [81][53], and a similar delay in leukocyte differentiation apparently occurs in the human liver [112][84].
To what extent does TIM4 expression reflect the length of stay and the depth of monocytic macrophage assimilation in the macrophage niche of KCs? In ouresearchers' opinion, this is a complex issue. OurResearchers own data obtained in a model of 70% liver resection in mice indicate that Tim4 expression in hepatic tissues can rapidly increase in the remnant liver tissue. This increase can be attributed to the influence of boosted endotoxin/LPS blood levels, and this suggestion has been supported by in vitro experiments [113][85]. Notably, KCs isolated from the intact liver by magnetic sorting for F4/80 had Tim4 expression levels similar to macrophages differentiated from peripheral blood monocytes isolated by magnetic sorting for CD115 and cultured with M-CSF. Under LPS exposure, Tim4 expression was significantly upregulated in both types of cultures. As it was also demonstrated, after liver resection in mice Ly6C+ monocytes migrate to it in high numbers. Considering the reduced Tim4 expression in the ‘newcomers’ [61][33], it would be reasonable to expect decreased expression of TIM4 marker in total liver macrophages after resection. However, this proved not to be the case, apparently due to the stimulating effect of LPS on the new, differentiating monocytic macrophages [113][85]. It can be concluded that, in this setting, Tim4 expression levels depend more on LPS exposure than on the length of residence in the macrophage niche of KCs [113][85].
Similar considerations and findings are applicable to another marker of resident liver macrophages, MARCO [81][53]. The use of this marker for distinguishing between macrophages arising from different sources has been based on the assumption that its expression is relatively constant and does not respond to endotoxin/LPS [81][53]. However, the latter point is questionable, considering the long-known role of this receptor in antimicrobial immunity [114,115][86][87]. True enough, KCs and bone marrow-derived macrophages express Marco at different levels; however, a sharp increase in Marco expression at both mRNA and protein levels in the liver remnant was observed after 70% hepatectomy, and this effect was reproduced by in vitro exposure of liver macrophage cultures to LPS [113][85]. Such dynamics of MARCO expression are consistent with its being a marker of pro-inflammatory state in murine macrophages [103][75].
The endotoxin/LPS sensitivity of the candidate macrophage markers is essential, as the vast portion of LPS in mammalian body is metabolized by the liver, not to mention the role of any tissue macrophages in antibacterial defense [116][88]. These considerations further implicate the tissue inflammatory status as a decisive factor which determines the phenotypical and functional properties of macrophages. The impact of inflammatory status on macrophage functionalities is especially prominent in the liver, given the barrier function of the organ. It has been already mentioned that various infections can promote a wave of cell death in resident macrophages, considered a ‘defensive suicide’ [85][57]. It is important to note that infectious lesions, which become sites of massive death of resident macrophages, are rapidly colonized by macrophages differentiating from the migratory blood monocytes. Notably, during influenza A infections, the resident alveolar macrophages become replaced by monocyte-derived macrophages, which appear to be more efficient in fighting Streptococcus pneumoniae infections due to higher production levels of IL6, CCL3, CCL4 and G-CSF [117,118][89][90]. Similarly, after a herpesvirus infection, alveolar macrophages effectively prevent the development of asthma by virtue of replacement of the ‘old’ resident alveolar macrophages by ‘new’ regulatory monocyte-derived macrophages that block the ability of dendritic cells to trigger Th2 responses [119][91].
A team of reseauthorchers observing an extraordinary robust immune response to S. pneumoniae following adenoviral infection attributed it to a previously unidentified ‘macrophage memory’ phenomenon, thereby suggesting the existence of memory cells in macrophage lineages [117,118][89][90]. In essence, the adenoviral infection causes activation of the resident alveolar macrophages, thus stimulating formation of a special self-perpetuating alveolar macrophage population. Upon activation, the naïve alveolar memory macrophages memorize the microenvironmental cues under the influence of Th cells and IFN-γ released by them [118][90]. The identified memory macrophage population is considered self-perpetuating and independent of blood monocytes. These cells are believed to retrieve the memorized information on previous inflammatory reactions to microbial pathogens in case of new infections; their action involves recruitment of neutrophils to the inflammatory foci [118][90].
The monocyte-derived macrophages which colonize the liver in the aftermath of KC depletion provide more efficient clearance of Neisseria meningitidis or Listeria monocytogenes by phagocytosis compared with KCs [81][53]; they also exert a more pronounced pro-inflammatory effect [76][48]. OuResearchers own in vitro experiments have demonstrated that, at early time points of stimulation, monocytic macrophages engulf latex particles at higher rates compared with KCs [87,120][59][92]. Such data can be interpreted in terms of hepatic tissue macrophage niche and its special features.
Over the entire history of research on the mononuclear phagocyte system in mammals macrophages were considered as cells with pronounced phenotypical plasticity confirmed in numerous studies [103][75]. Comparative evaluation of resident macrophages vs blood monocyte-derived macrophages shows higher sensitivity of the latter to activating factors and their higher phenotypic plasticity. Presumably, the prolonged exposure of resident macrophages to the conditions of organ-specific tissue niche leads to a reduction in plasticity through epigenetic block of inflammation-related genes. Such suppression is beneficial, as it suits the needs of the organ homeostasis. This view is based on research involving alveolar macrophages [79][51] and its validity for other resident macrophage populations, including those of the liver, has not been verified so far.
The liver has long been considered an immunotolerogenic organ [4]. Maintenance of this capacity is largely a responsibility and merit of KCs. Under normal conditions, KCs produce PD-L1, which participates in suppression of cellular immunity reactions, and small amounts of TNFa and IL-12. Under stimulation, KCs produce both pro-and anti-inflammatory cytokines [60[32][93],121], but their responses to many pathogen-associated molecular patterns (PAMPs), notably those of endotoxin/LPS are remarkably low. One of the probable reasons is the continuous exposure of KCs to LPS, the concentration of which in the portal blood flow varies within 0.1–1 ng/mL [116][88]. Such exposure presumably endows KCs with LPS tolerance, or at least reduced LPS sensitivity, compared with blood monocytes and monocyte-derived macrophages. OuResearchers own data agree with this assumption. For example, Tlr4 expression in peripheral blood monocytes is significantly higher compared with KCs [122[94][95],123], which is consistent with the evidence on more facile and LPS-sensitive induction of synthesis of certain interleukins in monocytic macrophages [87][59]. At the same time, studying the expression of LPS tolerance-associated genes in KCs and monocytic macrophages, no classical signatures of LPS tolerance were found. Still, KCs revealed lower expression of MAPK signaling-related genes Erk2 and p38 [124][96], known to participate in pro-inflammatory cytokine synthesis and release by macrophages. Reduced production of cytokines by macrophages has been associated with tolerance [125,126][97][98].
It can be assumed that similar mechanisms can reduce KC tolerance to PAMPs other than endotoxin/LPS: for instance, higher expression of Tlr2, Tlr7 and Tlr8 genes in monocytes compared with KCs was observed [122][94]. It should be also noted that the benefit of re-colonization with monocytic macrophages with regard to prevention of asthma and resistance to bacterial pathogens observed in the lungs cannot be straightforwardly extrapolated to the liver. A similar replacement scenario applied to KCs in the liver may have a deeply damaging effect and result in a chronic inflammatory process in hepatic tissues and at systemic level [76,90][48][62].
The introduction of the tissue inflammatory status as one of the parameters ‘in charge’ of the counts and properties of macrophages in particular organ may also ease the apparent controversy concerning the cell fate of monocytes colonizing the liver after toxic injury. As has been mentioned, these monocytes colonize the injured liver to become totally eliminated later on, despite a sharp decline of the resident macrophage populations and the overwhelming abundance of vacant niches. Presumably, the inflammatory status, and notably the time window of elevated TNFa and IL-1 levels determine the permission for monocytes to occupy the vacant niches previously occupied by KCs [80,82][52][54]. Still, even under this assumption, it is difficult to explain subsequent disappearance of these new monocytic lineages after resolution of the inflammatory process, especially given that at early stages of recovery the immigrating monocytes outweigh the preserved KCs numerically. For the reasons as yet unknown, in the toxically injured liver, surviving KCs clearly outcompete the immigrating monocytes/macrophages in settling the vacant niches. Incidentally, despite the just-experienced toxic shock, resident macrophages enter proliferation much earlier. The comparative dynamics imply that to make the monocytic impact visible, 80% of the resident liver macrophages should be depleted, which is hardly possible to achieve with available experimental techniques [80,105][52][77].
Notably, in the rat model of subtotal live resection, expression of Tnfa and Il1 increased significantly by the end of regeneration only, whereas the content of TNFa protein in the remnant was reduced from the beginning and stayed low since. This observation can be related to the lack of immigration of CX3CR1+ macrophages to the remnant liver in this model [73,127][45][99].
With these important amendments to the fundamental macrophage niche concept, the macrophage populations of individual organs are perceived as complex systems engaged in specific functionalities depending on their localization and ensuring the communication among different compartments inside each organ [105][77]. Moreover, macrophages have been implicated in the inter-organ crosstalk as well [39][7]. For example, myocardial overload leads to activation of the sympathetic innervation of the kidneys, resulting in enhanced secretion of S100A8/A9 peptides by the collecting duct epithelium. These peptides stimulate kidney macrophages to release TNFa, which promotes secretion of GM-CSF by endothelial cells in the interstitium. In return, the increased blood levels of GM-CSF promote accumulation of Ly6Clo macrophages in the myocardium; these macrophages produce amphiregulin, which causes hypertrophy of cardiomyocytes [128][100]. Similar data on the relationship between macrophages of the heart, lungs and kidneys were obtained in a model of myocardial infarction [129][101].
This new paradigm provides an unexpected explanation to the experimentally observed increased expression of certain interleukins and growth factors in lungs and kidneys after subtotal hepatectomy in rats [127,130][99][102]. The effect is accompanied by an increase in CD68+ macrophage counts in the lungs [131][103]. The coherence between monocyte/macrophage populations of the spleen and the liver is more comprehensible given the anatomical connection between the two organs via portal circulation [132,133][104][105]. At the same time, the spleen has been implicated as a monocyte supply for other organs as well. For instance, monocytes deposited in the spleen have been shown to migrate to inflammation foci in myocardial infarction and cerebral ischemic stroke [38,134,135][106][107][108].
Thus, the accumulated body of evidence suggests a general scheme of acquisition of unique properties by a tissue macrophage population. The first step involves implementation of the core macrophage differentiation program represented by PU.1, MYB, C-MAF, MAFB and ZEB2 transcription factors [136][109]. This universal basis becomes subsequently adjusted and refined by the influence of particular tissue niche, which shapes a transcription program characteristic of particular type of resident macrophages [39][7]. For KCs, the tissue niche comprises Ito cells, sinusoidal endothelial cells and hepatocytes, as well as various ECM components of hepatic parenchyma and paracrine factors; the joint influence of these components shapes a specific transcription program involving the LXR-α/ID3/SPIC expression signature in macrophages [80,111][52][83]. Interestingly, the transcriptional program of Kupffer cells turned out to be similar in mammals (humans, mice, pigs, hamsters and macaques), chicken and zebrafish [40][8], whereas induction of most genes unique to macrophages in these animal species required the interaction of activin receptor-like kinase (ALK1) on Kupffer cells with BMP9/10 secreted by Ito cells, and was also more or less TGFb dependent [40,111][8][83].
The concept of a macrophage niche is consistent with data on the epigenetic regulation of the resident macrophages phenotype, including those of the liver [137,138][110][111]. It has already been mentioned that the expression of the Clec4f gene, which encodes a lectin required for the presentation of alpha-galactosylceramide to natural killer T, is specific for the liver [99][71]. Assessment of histone modification status showed the presence of unique poised and active enhancers in the region of the Clec4f gene, as well as open chromatin regions in the region of the transcription factor LXRa, specific for Kupffer cells [138][111]. Histone acetylation sites were also found in the region of LXR gene, as well as in RBPJ gene, which is consistent with studies that established the dependence of the formation of a specific transcriptional program of Kupffer cells on the NOTCH-ligand DLL4 secreted by liver sinusoid endotheliocytes [111][83]. The effect of LPS on histone acetylation in the area of DNA regulatory regions during the differentiation of monocytes into macrophages has been also shown, which, given the constant contact with LPS, is especially important for Kupffer cells [139][112]. However, more detailed studies in this respect of Kupffer cells have not been conducted.
2. Macrophage Populations of the Liver
An accumulating body of evidence reveals extraordinary heterogeneity of liver macrophages, explained by multiple intersecting diversities (diverse sources of origin, diverse hepatic functionalities and diverse pathophysiological circumstances) [39][7]. According to the current state of knowledge, a normal liver harbors at least three populations of cells constituted by monocytic-macrophage lineages: (1) the dominating KCs; (2) cells with intermediate phenotypes between monocytes and macrophages; and (3) non-KC macrophages, including liver capsule macrophages, peritoneal macrophages and biliary tree-associated macrophages [40][8] resembling the Gpnmb+Spp1+ lipid-associated macrophages, LAMs [41][9]. In terms of embryonic origin, liver macrophage populations almost entirely descend from the erythro-myeloid progenitors of the yolk-sac wall [9,11][10][11]. These progenitors migrate through the left vitelline vein and umbilical vein to the embryonic liver, colonize it and embark on differentiation into KCs till later stages of fetal development and after birth [42][12]. Macrophages derived from blood monocytes constitute a minor fraction within the liver, commonly estimated within 10% of total liver macrophages and probably accounting for up to 30% in certain models [7,43,44][13][14][15]. Considering the diversity of embryonic sources and to avoid confusion, many reseauthorchers stick to the following nomenclature of liver macrophages. The term ‘Kupffer cells’ is assigned exclusively to macrophages descending from the erythro-myeloid sources of the yolk-sac wall [45,46][16][17]. However, other reseauthorchers use this term less scrupulously to refer to all macrophages found within the liver independently of their origin. AuthoResearchers of current reviewsearch favor the first option, as it really allows avoiding confusion while acknowledging the diversity of embryonic sources contributing to the hepatic macrophage populations. Estimates of the proportion of bone marrow-derived (monocytic) macrophages in the liver vary greatly due to the lack of unified set of markers [47,48][18][19]. In ouresearchers' opinion, the most accurate estimates are obtained with Ly6C and CX3CR1 [7,8,9,44,49][10][13][15][20][21]. Most studies of macrophage ontogeny have been performed on laboratory mice. Murine macrophages of bone marrow origin, in postnatal development, express Ly6C protein on their surface, whereas KCs either lack this marker or express it at low levels [44,50][15][22]. In rats, Ly6C protein has not been identified and CX3CR1 is used as a protein marker of bone marrow-derived (monocytic) macrophages instead of Ly6C [44][15]. As estimated with these markers, about 5% of total liver macrophages originate from the bone marrow [7,8,9,44,49][10][13][15][20][21]. These estimates are considered with morphological data on the dimensional and topographical diversity of liver macrophages. According to these observations, liver macrophages fall into two morphological subtypes: ‘large’, associated with sinusoidal capillaries, and ‘small’, located in the vicinity of central veins and portal tracts. Both subtypes express CD68, but only ‘large’ macrophages express high levels of CD163 [51,52][23][24]. Furthermore, ‘small’ macrophages constitute about 8% of the total liver macrophage counts, which matches the estimates of monocytic macrophage content obtained using Ly6C marker [7,44][13][15]. Hence, the populations of ‘small’ liver macrophages and liver cells of bone marrow origin may prove to be the same. Although many studies demonstrate high expression levels of mannose receptor protein CD206 by resident liver macrophages, dedicated analysis shows that the degree of CD206 positivity varies. Accordingly, KCs can be subdivided into two subsets: a predominant CD206loESAM- (KC1) and a CD206hiESAM+ minority (KC2) [53][25]. The KC2 cells have been shown to express genes that regulate fatty acid metabolism under normal and pathological conditions; moreover, KC2, which express high levels of CD36, have been shown to participate in regulation of the obesity-related oxidative stress in the liver. In addition, KC2 cells under the action of IL-2 promote CD8+ T cell activation thereby supporting the antiviral immunity [54][26]. The most controversial findings so far have been obtained with CD11b and CD68 markers expressed by a wide variety of cell types including all leukocytes; in addition, CD68 is expressed by endothelial cells and fibroblasts [55][27]. CD11b and CD68 participate in cell adhesion, migration and phagocytosis; accordingly their expression can undergo rapid changes [55,56,57][27][28][29]. The use of CD11b and CD68 complemented with F4/80 helps identify at least three F4/80+ liver macrophage subpopulations with distinct functionalities: cytokine-producing F4/80+CD11b+, highly phagocytic F4/80+CD68+ and F4/80+CD11b-CD68- with yet unknown function. CD11b+ liver macrophages presumably contribute to anti-tumor immunity [58][30] (Table 1).Table 1.
Macrophage populations of the liver.
| Population | Markers | Functions |
|---|---|---|
| Kupffer cells | F4/80 [58][30] CD68 [58][30] CD11b [59,60][31][32] CD163 [51,52][23][24] Cd206 (lo/hi) [53][25] Clec4F [61][33] Tim4 [61][33] |
Homeostatic |
| Non-KCs Macrophages/monocytes | ||
| Monocytes | Ly6C+ [44,[1550]][22] | Inflammation |
| Capsule macrophages | F4/80, CD14, CD64, CD207 [62,63][34][35] | Protection against pathogens invasion from the abdominal cavity |
| Peritoneal macrophages | CD102, GATA6 [64][36] | Unclear |
| Biliary tree-associated macrophages | Gpnmb [41][9] | Unclear, lipid metabolism |
3. Components of the Liver Macrophage Population Dynamics: Cell Migration, Cell Proliferation and Cell Death
The foundations of contemporary views of mononuclear-phagocytic system were laid in the 1970s by the works of Ralph van Furth. He suggested a concept of continuous replacement of tissue macrophages (with a naturally limited life span) by immigrating monocytes originating in the red bone marrow and transported by circulation. As the highly differentiated resident macrophages were assumed to lack the mitotic capacity, this view was fully justified and consistent with certain experimental findings [67][39]. Nevertheless, it eventually became evident that a majority of resident macrophage populations, including KCs, are self-perpetuating rather than dependent on the bone marrow hemopoiesis [68][40]. At the same time, the canonical macrophage differentiation scheme involving blood monocytes is possible as well and actually dominates in many postnatal tissues (Figure 1). For the liver, the canonical scheme is used as an emergency fallback, i.e., the infiltration of the liver with monocytes can be observed under pathological conditions only [69][41], whereas physiological contribution of monocyte immigration to the liver macrophage counts is negligible.Figure 1.
Component processes of the liver macrophage population dynamics.
4. KC-Specific Phenotypes
Gene and protein expression profiles identified with KCs are fairly flexible and depend on both the paracrine landscape and the long-range signals arriving from other organs [86][58]. Despite their pronounced molecular plasticity, KCs have been attributed with a particular immunophenotype [81,87][53][59]. Its hallmark proteins MARCO and CD163 are responsible for the recognition of bacterial pathogens and triggering of local immunity reactions; another KC marker, CD206, participates in antigen presentation, phagocytosis, cytokine production and pro-inflammatory mediator clearance [88][60]. In addition, resting KCs express the so-called tolerogenic phenotype variation with characteristic transcriptomic signature [60][32] including elevated expression of vascular permeability factors, ion channels, hemoglobin metabolism and complement system genes [81][53]. Such tolerogenic phenotypes are necessary for the active suppression of immune responses to the continuous influx of immunogenic agents accompanying the absorbed semi-metabolized nutrients and tissue debris [4,89][4][61]. Accordingly, KCs utilize enormous quantities of endotoxin/LPS without promoting inflammatory reactions. Moreover, LPS seems to trigger anti-inflammatory activation of KCs. Under these conditions, KCs suppress activation and proliferation of helper T cells while attracting regulatory T cells [90,91][62][63]. In addition, LPS may facilitate Fas-L expression by KCs, which triggers apoptosis in T cells [92][64]. At the same time, depletion of KCs causes fatal outcomes in bacterial infections with Listeria monocytogenes, Brucella burgdorferi or Staphylococcus aureus [93,94,95][65][66][67]. KCs also express the immunomodulatory PD-L1 and PD-1 shown, for example, to suppress the activity and proliferation of killer T cells in hepatocellular carcinoma or chronic hepatitis B [96][68]. The inhibition of PD-L1 expression by KCs in CMV infections stimulates the antiviral immunity [97][69]. KCs have been also shown to produce prostaglandins PGE2 and 15d-PGJ2 that interfere with the antigen-specific activation of T cells [98][70]. The loss of tolerogenic phenotype by Kupffer cells facilitates the development of inflammatory processes not only in the liver, but outside it as well [90][62]. Clec4F and Tim4 proteins and their corresponding transcripts are hallmarks for KC-specific molecular signatures [61][33]. Clec4F is a C-type lectin participating in antigen presentation of glycolipid antigens, as well as in the recognition and scavenging of desialylated platelets [99,100][71][72]. Tim4 is a phosphatidylserine-specific receptor allowing KCs to scavenge dying cells, also involved in triggering Th2 cell differentiation. Experimental inactivation of Tim4 improves the engraftment of the liver in allogeneic transplantations [101,102][73][74]. Overall, the diversity of KC phenotypes and gene expression signatures reflect the functional diversity characteristic of these liver-specific resident macrophages: KCs effectively combine the conventional macrophage functionalities, such as antigen presentation and phagocytosis, with organ-specific chores that involve elimination and scavenging of senescent formed elements, as well as tolerogenic influences.5. The Concept of Macrophage Niche and Its Application to KCs
The history of studies on the macrophage system in mammals is fairly long. The accumulating evidence has increasingly suggested that macrophages are extremely heterogeneous both phenotypically and functionally and thus must not be regarded as a single, uniform population [103][75]. Though specific prerequisites for such heterogeneity are uncertain, it apparently reflects the diversity of tissue microenvironments where macrophages differentiate and which are thought to largely define their phenotypes and functionalities [86][58]. These considerations eventually crystallized into the macrophage niche concept putting an emphasis on the unity of cellular components, extracellular matrix (ECM) and biologically active signaling molecules in providing the immediate environment for the tissue macrophage maturation [104][76]. The macrophage niche components ensure spatial compartmentalization/scaffolding and implement the trophic function; most notably in the context of this revisewarch, they provide macrophages with tissue-specific cellular identity by inducing particular sets of key transcription factor genes [80,105][52][77]. The macrophage niche concept is intended not just to explain the diversity of macrophage phenotypes and functionalities but also, very importantly, to account for the full-scale preservation of ‘relict’ macrophages of early hemopoietic origin in many organs and their non-replacement (resistance to replacement, as shown by experimental research) with descendants of fresher hemopoietic lineages [86,104][58][76]. The primary mechanistic explanation for this state of events in terms of macrophage niche involved three parameters: accessibility, vacancy and competition for the niche [86,104][58][76]. Accordingly, determination of macrophage composition at a particular location in the body could be reduced to interplay of these parameters. For instance, the persistence of microglia as the unique macrophage population in CNS could be explained by the non-accessibility of this location to monocytic precursors, due to the presence of the blood-brain barrier [86,104][58][76]. In contrast with the brain, the liver is accessible throughout postnatal development; however, by the time of full-fledged hemopoiesis in the bone marrow, all hepatic macrophage niches turn out to be occupied; upon injury, some of these niches go vacant which enables the immigration of monocytes from the blood and their subsequent differentiation into macrophages. In the lungs, the macrophage niches are constantly accessible and some of them vacant (free niches constantly appear physiologically), albeit on competitive terms, which explains the prolonged coexistence of macrophages from different sources in the lungs, with a gradual increase in the share of bone marrow-derived lung macrophages during postnatal life [86][58]. At the same time, some experimentally observed features of local macrophage populations in mammalian organs are only partially consistent with the fundamental macrophage niche concept. For example, monocytes arriving in the lungs are capable of differentiation into macrophages indistinguishable from the resident [106][78]. In a similar study, the liver depleted of resident macrophages was colonized by monocytes of bone marrow origin arriving from the blood. The macrophages differentiating from these monocytes were functionally similar to KC, but expressed a different profile of transcription factors [61,81][33][53]. Furthermore, a straightforward implementation of a lung-like scheme of macrophage colonization in the liver would imply a gradual increase in the content of bone marrow-derived macrophages as the liver grows. In laboratory rodents, the liver grows continuously throughout life, but the proportion of bone marrow-derived liver macrophages stays low—reaching 2–5% soon after birth and remaining at this level later on [8,9][10][20]. Complete ousting of the newly arrived bone marrow-derived (monocytic) macrophages by proliferating resident macrophages of the liver was demonstrated in a murine model of toxic liver injury [50][22]. A similar wave of monocyte immigration to the liver is observed after 70% liver resection in mice (although the fate of these cells has not been studied) but intriguingly not after subtotal resection (over 80% of the organ volume). According to the basic concept, the appearance of new macrophage niches during organ growth should promote monocyte/macrophage immigration as the cheapest route of replenishment [86][58]. The negligible rates of monocyte immigration in murine subtotal liver resection model may be related to low levels of MCP-1 production in the remnant [107][79]. The accumulation of new findings has fostered an attempt to modify the macrophage niche concept (Figure 2). The reseauthorchers dismiss the assumption that each organ comprises a single macrophage niche as inaccurate, and such opinion is consistent with representation of multiple macrophage lineages, including KCs, liver capsule macrophages and probably also peritoneal macrophages, in the liver [64][36]. A similar situation has been described for other organs, e.g., lungs, and even CNS microglia shows region-dependent heterogeneity [108,109][80][81]. Special types of macrophage populations have been identified in the so-called border zones, for example, the already mentioned macrophage population of the liver capsule, biliary tree-associated macrophages or macrophages found in the vicinity of mammary ducts [40,110][8][82].Figure 2. Characterization of the hepatic macrophage niche. KCs—Kupffer cells, HSCs—hepatic stellate cells (Ito cells), LSECs—liver sinusoidal endothelial cells, Hep—hepatocytes.
References
- Liaskou, E.; Wilson, D.V.; Oo, Y.H. Innate immune cells in liver inflammation. Mediat. Inflamm. 2012, 2012, 949157.
- Nguyen-Lefebvre, A.T.; Horuzsko, A. Kupffer Cell Metabolism and Function. J. Enzymol. Metab. 2015, 1, 101.
- Protzer, U.; Maini, M.K.; Knolle, P.A. Living in the liver: Hepatic infections. Nat. Rev. Immunol. 2012, 12, 201–213.
- Thomson, A.W.; Knolle, P.A. Antigen-presenting cell function in the tolerogenic liver environment. Nat. Rev. Immunol. 2010, 10, 753–766.
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186.
- Kolios, G.; Valatas, V.; Kouroumalis, E. Role of Kupffer cells in the pathogenesis of liver disease. World J. Gastroenterol. 2006, 12, 7413–7420.
- Blériot, C.; Chakarov, S.; Ginhoux, F. Determinants of Resident Tissue Macrophage Identity and Function. Immunity 2020, 52, 957–970.
- Guilliams, M.; Bonnardel, J.; Haest, B.; Vanderborght, B.; Wagner, C.; Remmerie, A.; Bujko, A.; Martens, L.; Thoné, T.; Browaeys, R.; et al. Spatial proteogenomics reveals distinct and evolutionarily conserved hepatic macrophage niches. Cell 2022, 185, 379–396.e38.
- Remmerie, A.; Martens, L.; Thoné, T.; Castoldi, A.; Seurinck, R.; Pavie, B.; Roels, J.; Vanneste, B.; De Prijck, S.; Vanhockerhout, M.; et al. Osteopontin Expression Identifies a Subset of Recruited Macrophages Distinct from Kupffer Cells in the Fatty Liver. Immunity 2020, 53, 641–657.e14.
- Perdiguero, E.G.; Geissmann, F. The development and maintenance of resident macrophages. Nat. Immunol. 2016, 17, 2–8.
- Hoeffel, G.; Ginhoux, F. Fetal monocytes and the origins of tissue-resident macrophages. Cell. Immunol. 2018, 330, 5–15.
- Naito, M.; Hasegawa, G.; Kiyoshi, T. Development, differentiation, and maturation of Kupffer cells. Microsc. Res. Tech. 1997, 39, 350–364.
- Chazaud, B. Macrophages: Supportive cells for tissue repair and regeneration. Immunobiology 2014, 219, 172–178.
- Kinoshita, M.; Uchida, T.; Sato, A.; Nakashima, M.; Nakashima, H.; Shono, S.; Habu, Y.; Miyazaki, H.; Hiroi, S.; Seki, S. Characterization of two F4/80-positive Kupffer cell subsets by their function and phenotype in mice. J. Hepatol. 2010, 53, 903–910.
- Zigmond, E.; Samia-Grinberg, S.; Pasmanik-Chor, M.; Brazowski, E.; Shibolet, O.; Halpern, Z.; Varol, C. Infiltrating Monocyte-Derived Macrophages and Resident Kupffer Cells Display Different Ontogeny and Functions in Acute Liver Injury. J. Immunol. 2014, 193, 344–353.
- Guillot, A.; Tacke, F. Liver Macrophages: Old Dogmas and New Insights. Hepatol. Commun. 2019, 3, 730–743.
- Shan, Z.; Ju, C. Hepatic Macrophages in Liver Injury. Front. Immunol. 2020, 11, 322.
- Wen, Y.; Lambrecht, J.; Ju, C.; Tacke, F. Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell. Mol. Immunol. 2020, 18, 45–56.
- Ju, C.; Tacke, F. Hepatic macrophages in homeostasis and liver diseases: From pathogenesis to novel therapeutic strategies. Cell. Mol. Immunol. 2016, 13, 316–327.
- Gomez Perdiguero, E.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; De Bruijn, M.F.; Geissmann, F.; et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 2015, 518, 547–551.
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and Functions of Tissue Macrophages. Immunity 2014, 41, 21–35.
- You, Q.; Holt, M.; Yin, H.; Li, G.; Hu, C.J.; Ju, C. Role of hepatic resident and infiltrating macrophages in liver repair after acute injury. Biochem. Pharmacol. 2013, 86, 836–843.
- Armbrust, T.; Ramadori, G. Functional characterization of two different Kupffer cell populations of normal rat liver. J. Hepatol. 1996, 25, 518–528.
- He, Y.; Sadahiro, T.; Noh, S.I.; Wang, H.; Todo, T.; Chai, N.N.; Klein, A.S.; Wu, G.D. Flow cytometric isolation and phenotypic characterization of two subsets of ED2+ (CD163) hepatic macrophages in rats. Hepatol. Res. 2009, 39, 1208–1218.
- Blériot, C.; Barreby, E.; Dunsmore, G.; Ballaire, R.; Chakarov, S.; Ficht, X.; De Simone, G.; Andreata, F.; Fumagalli, V.; Guo, W.; et al. A subset of Kupffer cells regulates metabolism through the expression of CD36. Immunity 2021, 54, 2101–2116.e6.
- De Simone, G.; Andreata, F.; Bleriot, C.; Fumagalli, V.; Laura, C.; Garcia-Manteiga, J.M.; Di Lucia, P.; Gilotto, S.; Ficht, X.; De Ponti, F.F.; et al. Identification of a Kupffer cell subset capable of reverting the T cell dysfunction induced by hepatocellular priming. Immunity 2021, 54, 2089–2100.e8.
- Gottfried, E.; Kunz-Schughart, L.A.; Weber, A.; Rehli, M.; Peuker, A.; Müller, A.; Kastenberger, M.; Brockhoff, G.; Andreesen, R.; Kreutz, M. Expression of CD68 in Non-Myeloid Cell Types. Scand. J. Immunol. 2008, 67, 453–463.
- Sanchez-Madrid, F.; Simon, P.; Thompson, S.; Springer, T.A. Mapping of antigenic and functional epitopes on the alpha- and beta-subunits of two related mouse glycoproteins involved in cell interactions, LFA-1 and Mac-1. J. Exp. Med. 1983, 158, 586–602.
- Schittenhelm, L.; Hilkens, C.M.; Morrison, V.L. β2 Integrins As Regulators of Dendritic Cell, Monocyte, and Macrophage Function. Front. Immunol. 2017, 8, 1866.
- Ikarashi, M.; Nakashima, H.; Kinoshita, M.; Sato, A.; Nakashima, M.; Miyazaki, H.; Nishiyama, K.; Yamamoto, J.; Seki, S. Distinct development and functions of resident and recruited liver Kupffer cells/macrophages. J. Leukoc. Biol. 2013, 94, 1325–1336.
- Haldar, M.; Murphy, K.M. Origin, development, and homeostasis of tissue-resident macrophages. Immunol. Rev. 2014, 262, 25–35.
- Movita, D.; Kreefft, K.; Biesta, P.; van Oudenaren, A.; Leenen, P.J.M.; Janssen, H.L.A.; Boonstra, A. Kupffer cells express a unique combination of phenotypic and functional characteristics compared with splenic and peritoneal macrophages. J. Leukoc. Biol. 2012, 92, 723–733.
- Scott, C.L.; Zheng, F.; De Baetselier, P.; Martens, L.; Saeys, Y.; De Prijck, S.; Lippens, S.; Abels, C.; Schoonooghe, S.; Raes, G.; et al. Bone marrow-derived monocytes give rise to self-renewing and fully differentiated Kupffer cells. Nat. Commun. 2016, 7, 10321.
- Sierro, F.; Evrard, M.; Rizzetto, S.; Melino, M.; Mitchell, A.J.; Florido, M.; Beattie, L.; Walters, S.B.; Tay, S.S.; Lu, B.; et al. A Liver Capsular Network of Monocyte-Derived Macrophages Restricts Hepatic Dissemination of Intraperitoneal Bacteria by Neutrophil Recruitment. Immunity 2017, 47, 374–388.e6.
- David, B.A.; Rezende, R.M.; Antunes, M.M.; Santos, M.M.; Freitas Lopes, M.A.; Diniz, A.B.; Sousa Pereira, R.V.; Marchesi, S.C.; Alvarenga, D.M.; Nakagaki, B.N.; et al. Combination of Mass Cytometry and Imaging Analysis Reveals Origin, Location, and Functional Repopulation of Liver Myeloid Cells in Mice. Gastroenterology 2016, 151, 1176–1191.
- Wang, J.; Kubes, P. A Reservoir of Mature Cavity Macrophages that Can Rapidly Invade Visceral Organs to Affect Tissue Repair. Cell 2016, 165, 668–678.
- Nishiyama, K.; Nakashima, H.; Ikarashi, M.; Kinoshita, M.; Nakashima, M.; Aosasa, S.; Seki, S.; Yamamoto, J. Mouse CD11b+Kupffer cells recruited from bone marrow accelerate liver regeneration after partial hepatectomy. PLoS ONE 2015, 10, e0136774.
- Krenkel, O.; Hundertmark, J.; Abdallah, A.T.; Kohlhepp, M.; Puengel, T.; Roth, T.; Branco, D.P.P.; Mossanen, J.C.; Luedde, T.; Trautwein, C.; et al. Myeloid cells in liver and bone marrow acquire a functionally distinct inflammatory phenotype during obesity-related steatohepatitis. Gut 2019, 69, 551–563.
- Van Furth, R. Monocyte origin of Kupffer cells. Blood Cells 1980, 6, 87–92.
- Jenkins, S.J.; Ruckerl, D.; Cook, P.C.; Jones, L.H.; Finkelman, F.D.; Van Rooijen, N.; MacDonald, A.S.; Allen, J.E. Local macrophage proliferation, rather than recruitment from the blood, is a signature of T H2 inflammation. Science 2011, 332, 1284–1288.
- Karlmark, K.R.; Weiskirchen, R.; Zimmermann, H.W.; Gassler, N.; Ginhoux, F.; Weber, C.; Merad, M.; Luedde, T.; Trautwein, C.; Tacke, F. Hepatic recruitment of the inflammatory Gr1+ monocyte subset upon liver injury promotes hepatic fibrosis. Hepatology 2009, 50, 261–274.
- Hashimoto, D.; Chow, A.; Noizat, C.; Teo, P.; Beasley, M.B.; Leboeuf, M.; Becker, C.D.; See, P.; Price, J.; Lucas, D.; et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity 2013, 38, 792–804.
- Ajami, B.; Bennett, J.L.; Krieger, C.; Tetzlaff, W.; Rossi, F.M.V. Local self-renewal can sustain CNS microglia maintenance and function throughout adult life. Nat. Neurosci. 2007, 10, 1538–1543.
- Jenkins, S.J.; Ruckerl, D.; Thomas, G.D.; Hewitson, J.P.; Duncan, S.; Brombacher, F.; Maizels, R.M.; Hume, D.A.; Allen, J.E. IL-4 directly signals tissue-resident macrophages to proliferate beyond homeostatic levels controlled by CSF-1. J. Exp. Med. 2013, 210, 2477–2491.
- Elchaninov, A.V.; Fatkhudinov, T.K.; Usman, N.Y.; Kananykhina, E.Y.; Arutyunyan, I.V.; Makarov, A.V.; Lokhonina, A.V.; Eremina, I.Z.; Surovtsev, V.V.; Goldshtein, D.V.; et al. Dynamics of macrophage populations of the liver after subtotal hepatectomy in rats. BMC Immunol. 2018, 19, 23.
- Elchaninov, A.; Nikitina, M.; Vishnyakova, P.; Lokhonina, A.; Makarov, A.; Sukhikh, G.; Fatkhudinov, T. Macro- and microtranscriptomic evidence of the monocyte recruitment to regenerating liver after partial hepatectomy in mouse model. Biomed. Pharmacother. 2021, 138, 111516.
- Ait Ahmed, Y.; Fu, Y.; Rodrigues, R.M.; He, Y.; Guan, Y.; Guillot, A.; Ren, R.; Feng, D.; Hidalgo, J.; Ju, C.; et al. Kupffer cell restoration after partial hepatectomy is mainly driven by local cell proliferation in IL-6-dependent autocrine and paracrine manners. Cell. Mol. Immunol. 2021, 18, 2165–2176.
- Tran, S.; Baba, I.; Poupel, L.; Dussaud, S.; Moreau, M.; Gélineau, A.; Marcelin, G.; Magréau-Davy, E.; Ouhachi, M.; Lesnik, P.; et al. Impaired Kupffer Cell Self-Renewal Alters the Liver Response to Lipid Overload during Non-alcoholic Steatohepatitis. Immunity 2020, 53, 627–640.e5.
- Jin, H.; Liu, K.; Tang, J.; Huang, X.; Wang, H.; Zhang, Q.; Zhu, H.; Li, Y.; Pu, W.; Zhao, H.; et al. Genetic fate-mapping reveals surface accumulation but not deep organ invasion of pleural and peritoneal cavity macrophages following injury. Nat. Commun. 2021, 12, 2863.
- Blériot, C.; Ginhoux, F. Understanding the Heterogeneity of Resident Liver Macrophages. Front. Immunol. 2019, 10, 2694.
- Guilliams, M.; Svedberg, F.R. Does tissue imprinting restrict macrophage plasticity? Nat. Immunol. 2021, 22, 118–127.
- Bonnardel, J.; T’Jonck, W.; Gaublomme, D.; Browaeys, R.; Scott, C.L.; Martens, L.; Vanneste, B.; De Prijck, S.; Nedospasov, S.A.; Kremer, A.; et al. Stellate Cells, Hepatocytes, and Endothelial Cells Imprint the Kupffer Cell Identity on Monocytes Colonizing the Liver Macrophage Niche. Immunity 2019, 51, 638–654.e9.
- Beattie, L.; Sawtell, A.; Mann, J.; Frame, T.C.M.; Teal, B.; de Labastida Rivera, F.; Brown, N.; Walwyn-Brown, K.; Moore, J.W.J.; MacDonald, S.; et al. Bone marrow-derived and resident liver macrophages display unique transcriptomic signatures but similar biological functions. J. Hepatol. 2016, 65, 758–768.
- Blériot, C.; Dupuis, T.; Jouvion, G.; Eberl, G.; Disson, O.; Lecuit, M. Liver-Resident Macrophage Necroptosis Orchestrates Type 1 Microbicidal Inflammation and Type-2-Mediated Tissue Repair during Bacterial Infection. Immunity 2015, 42, 145–158.
- DiPaolo, N.C.; Doronin, K.; Baldwin, L.K.; Papayannopoulou, T.; Shayakhmetov, D.M. The Transcription Factor IRF3 Triggers “Defensive Suicide” Necrosis in Response to Viral and Bacterial Pathogens. Cell Rep. 2013, 3, 1840–1846.
- Lai, S.M.; Sheng, J.; Gupta, P.; Renia, L.; Duan, K.; Zolezzi, F.; Karjalainen, K.; Newell, E.W.; Ruedl, C. Organ-Specific Fate, Recruitment, and Refilling Dynamics of Tissue-Resident Macrophages during Blood-Stage Malaria. Cell Rep. 2018, 25, 3099–3109.e3.
- Ginhoux, F.; Bleriot, C.; Lecuit, M. Dying for a Cause: Regulated Necrosis of Tissue-Resident Macrophages upon Infection. Trends Immunol. 2017, 38, 693–695.
- Guilliams, M.; Scott, C.L. Does niche competition determine the origin of tissue-resident macrophages? Nat. Rev. Immunol. 2017, 17, 451–460.
- Lokhonina, A.; Elchaninov, A.; Fatkhudinov, T.; Makarov, A.; Arutyunyan, I.; Grinberg, M.; Glinkina, V.; Surovtsev, V.; Bolshakova, G.; Goldshtein, D.; et al. Activated Macrophages of Monocytic Origin Predominantly Express Proinflammatory Cytokine Genes, whereas Kupffer Cells Predominantly Express Anti-Inflammatory Cytokine Genes. Biomed Res. Int. 2019, 2019, 3912142.
- Martinez-Pomares, L. The mannose receptor. J. Leukoc. Biol. 2012, 92, 1177–1186.
- Gorezynski, R.M. Immunosuppression induced by hepatic portal venous immunization spares reactivity in IL-4 producing T lymphocytes. Immunol. Lett. 1992, 33, 67–77.
- Heymann, F.; Peusquens, J.; Ludwig-Portugall, I.; Kohlhepp, M.; Ergen, C.; Niemietz, P.; Martin, C.; van Rooijen, N.; Ochando, J.C.; Randolph, G.J.; et al. Liver inflammation abrogates immunological tolerance induced by Kupffer cells. Hepatology 2015, 62, 279–291.
- Li, X.; Wang, Z.; Zou, Y.; Lu, E.; Duan, J.; Yang, H.; Wu, Q.; Zhao, X.; Wang, Y.; You, L.; et al. Pretreatment with lipopolysaccharide attenuates diethylnitrosamine-caused liver injury in mice via TLR4-dependent induction of Kupffer cell M2 polarization. Immunol. Res. 2015, 62, 137–145.
- Uchikura, K.; Wada, T.; Hoshino, S.; Nagakawa, Y.; Aiko, T.; Bulkley, G.B.; Klein, A.S.; Sun, Z. Lipopolysaccharides induced increases in Fas ligand expression by Kupffer cells via mechanisms dependent on reactive oxygen species. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G620–G626.
- Lee, W.Y.; Moriarty, T.J.; Wong, C.H.Y.; Zhou, H.; Strieter, R.M.; Van Rooijen, N.; Chaconas, G.; Kubes, P. An intravascular immune response to Borrelia burgdorferi involves Kupffer cells and iNKT cells. Nat. Immunol. 2010, 11, 295–302.
- Ebe, Y.; Hasegawa, G.; Takatsuka, H.; Umezu, H.; Mitsuyama, M.; Arakawa, M.; Mukaida, N.; Naito, M. The role of Kupffer cells and regulation of neutrophil migration into the liver by macrophage inflammatory protein-2 in primary listeriosis in mice. Pathol. Int. 1999, 49, 519–532.
- Helmy, K.Y.; Katschke, K.J.; Gorgani, N.N.; Kljavin, N.M.; Elliott, J.M.; Diehl, L.; Scales, S.J.; Ghilardi, N.; Van Lookeren Campagne, M. CRIg: A macrophage complement receptor required for phagocytosis of circulating pathogens. Cell 2006, 124, 915–927.
- Ke, W.; Kryczek, I.; Chen, L.; Zou, W.; Welling, T.H. Kupffer cell suppression of CD8+ T cells in human hepatocellular carcinoma is mediated by B7-H1/programmed death-1 interactions. Cancer Res. 2009, 69, 8067–8075.
- Dolina, J.S.; Sung, S.S.J.; Novobrantseva, T.I.; Nguyen, T.M.; Hahn, Y.S. Lipidoid Nanoparticles Containing PD-L1 siRNA Delivered In Vivo Enter Kupffer Cells and Enhance NK and CD8(+) T Cell-mediated Hepatic Antiviral Immunity. Mol. Ther. Nucleic Acids 2013, 2, e72.
- You, Q.; Cheng, L.; Kedl, R.M.; Ju, C. Mechanism of T cell tolerance induction by murine hepatic Kupffer cells. Hepatology 2008, 48, 978–990.
- Yang, C.; Chen, J.B.; Tsai, T.F.; Tsai, Y.C.; Tsai, C.Y.; Liang, P.H.; Hsu, T.L.; Wu, C.Y.; Netea, M.G.; Wong, C.H.; et al. CLEC4F is an inducible C-type lectin in F4/80-positive cells and is involved in alpha-galactosylceramide presentation in liver. PLoS ONE 2013, 8, e65070.
- Jiang, Y.; Tang, Y.; Hoover, C.; Kondo, Y.; Huang, D.; Restagno, D.; Shao, B.; Gao, L.; McDaniel, J.M.; Zhou, M.; et al. Kupffer cell receptor CLEC4F is important for the destruction of desialylated platelets in mice. Cell Death Differ. 2021, 28, 3009–3021.
- Wu, H.; Xu, X.; Li, J.; Gong, J.; Li, M. TIM-4 blockade of KCs combined with exogenous TGF-β injection helps to reverse acute rejection and prolong the survival rate of mice receiving liver allografts. Int. J. Mol. Med. 2018, 42, 346–358.
- Martina, M.; McGrath, M. Diverse roles of TIM4 in immune activation: Implications for alloimmunity. Curr. Opin. Organ Transplant. 2018, 23, 44–50.
- Murray, P.J.J.; Allen, J.E.E.; Biswas, S.K.K.; Fisher, E.A.A.; Gilroy, D.W.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.A.; Ivashkiv, L.B.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20.
- T’Jonck, W.; Guilliams, M.; Bonnardel, J. Niche signals and transcription factors involved in tissue-resident macrophage development. Cell. Immunol. 2018, 330, 43–53.
- Guilliams, M.; Thierry, G.R.; Bonnardel, J.; Bajenoff, M. Establishment and Maintenance of the Macrophage Niche. Immunity 2020, 52, 434–451.
- Kulikauskaite, J.; Wack, A. Teaching Old Dogs New Tricks? The Plasticity of Lung Alveolar Macrophage Subsets. Trends Immunol. 2020, 41, 864–877.
- Wyler, S.L.; D’Ingillo, S.L.; Lamb, C.L.; Mitchell, K.A. Monocyte chemoattractant protein-1 is not required for liver regeneration after partial hepatectomy. J. Inflamm. 2016, 13, 28.
- Chakarov, S.; Lim, H.Y.; Tan, L.; Lim, S.Y.; See, P.; Lum, J.; Zhang, X.M.; Foo, S.; Nakamizo, S.; Duan, K.; et al. Two distinct interstitial macrophage populations coexist across tissues in specific subtissular niches. Science 2019, 363, eaau0964.
- Böttcher, C.; Schlickeiser, S.; Sneeboer, M.A.M.; Kunkel, D.; Knop, A.; Paza, E.; Fidzinski, P.; Kraus, L.; Snijders, G.J.L.; Kahn, R.S.; et al. Human microglia regional heterogeneity and phenotypes determined by multiplexed single-cell mass cytometry. Nat. Neurosci. 2018, 22, 78–90.
- Dawson, C.A.; Pal, B.; Vaillant, F.; Gandolfo, L.C.; Liu, Z.; Bleriot, C.; Ginhoux, F.; Smyth, G.K.; Lindeman, G.J.; Mueller, S.N.; et al. Tissue-resident ductal macrophages survey the mammary epithelium and facilitate tissue remodelling. Nat. Cell Biol. 2020, 22, 546–558.
- Sakai, M.; Troutman, T.D.; Seidman, J.S.; Ouyang, Z.; Spann, N.J.; Abe, Y.; Ego, K.M.; Bruni, C.M.; Deng, Z.; Schlachetzki, J.C.M.; et al. Liver-Derived Signals Sequentially Reprogram Myeloid Enhancers to Initiate and Maintain Kupffer Cell Identity. Immunity 2019, 51, 655–670.e8.
- Pallett, L.J.; Burton, A.R.; Amin, O.E.; Rodriguez-Tajes, S.; Patel, A.A.; Zakeri, N.; Jeffery-Smith, A.; Swadling, L.; Schmidt, N.M.; Baiges, A.; et al. Longevity and replenishment of human liver-resident memory T cells and mononuclear phagocytes. J. Exp. Med. 2020, 217, e20200050.
- Elchaninov, A.; Lokhonina, A.; Vishnyakova, P.; Soboleva, A.; Poltavets, A.; Artemova, D.; Makarov, A.; Glinkina, V.; Goldshtein, D.; Bolshakova, G.; et al. Marco+ macrophage dynamics in regenerating liver after 70% liver resection in mice. Biomedicines 2021, 9, 1129.
- Kangas, M.; Brännström, A.; Elomaa, O.; Matsuda, Y.; Eddy, R.; Shows, T.B.; Tryggvason, K. Structure and chromosomal localization of the human and murine genes for the macrophage MARCO receptor. Genomics 1999, 58, 82–89.
- Kraal, G.; Van Der Laan, L.J.W.; Elomaa, O.; Tryggvason, K. The macrophage receptor MARCO. Microbes Infect. 2000, 2, 313–316.
- Knolle, P.A.; Germann, T.; Treichel, U. Endotoxin down-regulates T cell activation by antigen-presenting liver sinusoidal endothelial cells. J. Immunol. 1999, 162, 1401–1407.
- Aegerter, H.; Kulikauskaite, J.; Crotta, S.; Patel, H.; Kelly, G.; Hessel, E.M.; Mack, M.; Beinke, S.; Wack, A. Influenza-induced monocyte-derived alveolar macrophages confer prolonged antibacterial protection. Nat. Immunol. 2020, 21, 145–157.
- Yao, Y.; Jeyanathan, M.; Haddadi, S.; Barra, N.G.; Vaseghi-Shanjani, M.; Damjanovic, D.; Lai, R.; Afkhami, S.; Chen, Y.; Dvorkin-Gheva, A.; et al. Induction of Autonomous Memory Alveolar Macrophages Requires T Cell Help and Is Critical to Trained Immunity. Cell 2018, 175, 1634–1650.e17.
- Machiels, B.; Dourcy, M.; Xiao, X.; Javaux, J.; Mesnil, C.; Sabatel, C.; Desmecht, D.; Lallemand, F.; Martinive, P.; Hammad, H.; et al. A gammaherpesvirus provides protection against allergic asthma by inducing the replacement of resident alveolar macrophages with regulatory monocytes. Nat. Immunol. 2017, 18, 1310–1320.
- Lokhonina, A.V.; Makarov, A.V.; Elchaninov, A.V.; Arutyunyan, I.V.; Shmakova, T.V.; Grinberg, M.V.; Usman, N.Y.; Surovtsev, V.V.; Chernikov, V.P.; Fatkhudinov, T.K. Quantitative and Qualitative Characterization of Phagocytic Activity of Macrophages of Bone Marrow and Fetal Origin. Bull. Exp. Biol. Med. 2019, 167, 154–158.
- Horst, A.K.; Neumann, K.; Diehl, L.; Tiegs, G. Modulation of liver tolerance by conventional and nonconventional antigen-presenting cells and regulatory immune cells. Cell. Mol. Immunol. 2016, 13, 277–292.
- Elchaninov, A.; Lokhonina, A.; Nikitina, M.; Vishnyakova, P.; Makarov, A.; Arutyunyan, I.; Poltavets, A.; Kananykhina, E.; Kovalchuk, S.; Karpulevich, E.; et al. Comparative analysis of the transcriptome, proteome, and mirna profile of kupffer cells and monocytes. Biomedicines 2020, 8, 627.
- Nikitina, M.P.; Elchaninov, A.V.; Lokhonina, A.V.; Makarov, A.V.; Tagirova, M.K.; Grinberg, M.V.; Bolshakova, G.B.; Glinkina, V.V.; Goldshtein, D.V.; Fatkhudinov, T.K. Analysis of the Expression of Regulator Genes in Kupffer Cells and Monocytes. Bull. Exp. Biol. Med. 2020, 168, 556–560.
- Lokhonina, A.V.; Elchaninov, A.V.; Makarov, A.V.; Nikitina, M.P.; Goldshtein, D.V.; Paltsev, M.A.; Fatkhudinov, T.K. Comparative characteristics of the susceptibility of kupffer cells and macrophages of bone-background origin to activation factors. Mol. Med. 2019, 17, 1032.
- Liu, D.; Cao, S.; Zhou, Y.; Xiong, Y. Recent advances in endotoxin tolerance. J. Cell. Biochem. 2019, 120, 56–70.
- Nimah, M.; Zhao, B.; Denenberg, A.G.; Bueno, O.; Molkentin, J.; Wong, H.R.; Shanley, T.P. Contribution of MKP-1 regulation of p38 to endotoxin tolerance. Shock 2005, 23, 80–87.
- Elchaninov, A.; Fatkhudinov, T.; Usman, N.; Kananykhina, E.; Arutyunyan, I.; Makarov, A.; Bolshakova, G.; Goldshtein, D.; Sukhikh, G. Molecular survey of cell source usage during subtotal hepatectomy-inducedliver regeneration in rats. PLoS ONE 2016, 11, e0162613.
- Fujiu, K.; Shibata, M.; Nakayama, Y.; Ogata, F.; Matsumoto, S.; Noshita, K.; Iwami, S.; Nakae, S.; Komuro, I.; Nagai, R.; et al. A heart-brain-kidney network controls adaptation to cardiac stress through tissue macrophage activation. Nat. Med. 2017, 23, 611–622.
- Hoyer, F.F.; Naxerova, K.; Schloss, M.J.; Hulsmans, M.; Nair, A.V.; Dutta, P.; Calcagno, D.M.; Herisson, F.; Anzai, A.; Sun, Y.; et al. Tissue-Specific Macrophage Responses to Remote Injury Impact the Outcome of Subsequent Local Immune Challenge. Immunity 2019, 51, 899–914.e7.
- Elchaninov, A.V.; Fatkhudinov, T.K.; Usman, N.Y.; Arutyunyan, I.V.; Makarov, A.V.; Kananykhina, E.Y.; Glinkina, V.V.; Bolshakova, G.B.; Sukhikh, G.T. Expression of cytokine genes and growth factors in rat lungs and kidneys after subtotal hepatectomy. Bull. Exp. Biol. Med. 2016, 161, 373–377.
- El’chaninov, A.V.; Fatkhudinov, T.K.; Arutyunyan, I.V.; Makarov, A.V.; Usman, N.Y.; Mikhailova, L.P.; Lokhonina, A.V.; Botchey, V.M.; Glinkina, V.V.; Bol’shakova, G.B. Dynamics of Expression of Cytokine Genes and Macrophage Content in the Lungs and Kidneys after Subtotal Hepatectomy in Rats. Bull. Exp. Biol. Med. 2018, 165, 136–141.
- Elchaninov, A.V.; Fatkhudinov, T.K.; Vishnyakova, P.A.; Nikitina, M.P.; Lokhonina, A.V.; Makarov, A.V.; Arutyunyan, I.V.; Kananykhina, E.Y.; Poltavets, A.S.; Butov, K.R.; et al. Molecular mechanisms of splenectomy-induced hepatocyte proliferation. PloS One 2020, 15, e0233767.
- Elchaninov, A.; Vishnyakova, P.; Sukhikh, G.; Fatkhudinov, T. Spleen: Reparative Regeneration and Influence on Liver. Life 2022, 12, 626.
- Teh, Y.C.; Ding, J.L.; Ng, L.G.; Chong, S.Z. Capturing the Fantastic Voyage of Monocytes Through Time and Space. Front. Immunol. 2019, 10, 834.
- Kim, E.; Yang, J.; Beltran, C.D.; Cho, S. Role of spleen-derived monocytes/macrophages in acute ischemic brain injury. J. Cereb. Blood Flow Metab. 2014, 34, 1411–1419.
- Leuschner, F.; Rauch, P.J.; Ueno, T.; Gorbatov, R.; Marinelli, B.; Lee, W.W.; Dutta, P.; Wei, Y.; Robbins, C.; Iwamoto, Y.; et al. Rapid monocyte kinetics in acute myocardial infarction are sustained by extramedullary monocytopoiesis. J. Exp. Med. 2012, 209, 123.
- Scott, C.L.; T’Jonck, W.; Martens, L.; Todorov, H.; Sichien, D.; Soen, B.; Bonnardel, J.; De Prijck, S.; Vandamme, N.; Cannoodt, R.; et al. The Transcription Factor ZEB2 Is Required to Maintain the Tissue-Specific Identities of Macrophages. Immunity 2018, 49, 312–325.e5.
- Gosselin, D.; Link, V.M.; Romanoski, C.E.; Fonseca, G.J.; Eichenfield, D.Z.; Spann, N.J.; Stender, J.D.; Chun, H.B.; Garner, H.; Geissmann, F.; et al. Environment Drives Selection and Function of Enhancers Controlling Tissue-Specific Macrophage Identities. Cell 2014, 159, 1327–1340.
- Lavin, Y.; Winter, D.; Blecher-Gonen, R.; David, E.; Keren-Shaul, H.; Merad, M.; Jung, S.; Amit, I. Tissue-resident macrophage enhancer landscapes are shaped by the local microenvironment. Cell 2014, 159, 1312–1326.
- Saeed, S.; Quintin, J.; Kerstens, H.H.D.; Rao, N.A.; Aghajanirefah, A.; Matarese, F.; Cheng, S.C.; Ratter, J.; Berentsem, K.; Van Der Ent, M.A.; et al. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science 2014, 345, 1251086.
More