L-dopa is a precursor of dopamine used as the most effective symptomatic drug treatment for Parkinson’s disease. Most of the L-dopa isolated is either synthesized chemically or from natural sources, but only some plants belonging to the Fabaceae family contain significant amounts of L-dopa. Due to its low stability, the unambiguous determination of L-dopa in plant matrices requires appropriate technologies. Several analytical methods have been developed for the determination of L-dopa in different plants. The most used for quantification of L-dopa are mainly based on capillary electrophoresis or chromatographic methods, i.e., high-performance liquid chromatography (HPLC), coupled to ultraviolet-visible or mass spectrometric detection. HPLC is most often used.
- levodopa
- plant matrices
- extraction
- chromatographic methods
1. Introduction
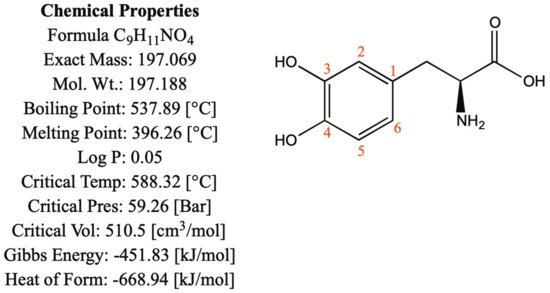
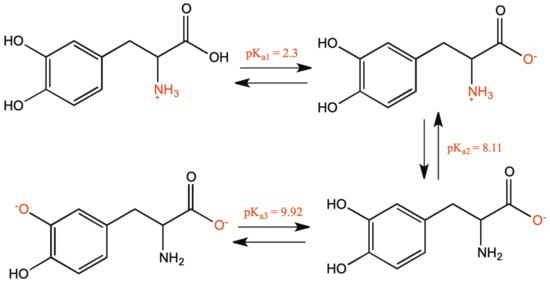
1.1. Chemical and Physical Properties
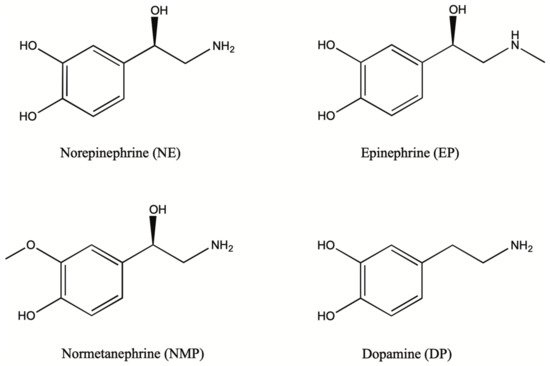
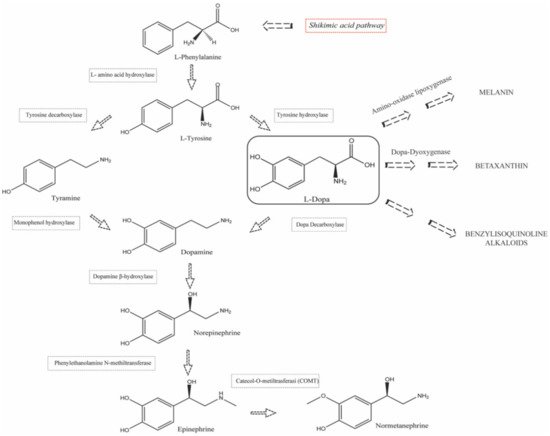
1.2. Biosynthesis and Conversion Routes of Levodopa in Plants
2. Levodopa Extraction Techniques
|
Extraction Technique |
Matrix |
Variety |
|---|
3. Levodopa Detection Methods
|
Methods |
Sample Source |
LOD Range | Solvents and Optimized Conditions |
Stationary Phase Recovery Percent (Mean Value Percent) |
References |
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Mobile Phase | Detection Mode |
Strengths |
Drawbacks |
References |
|||||||
|
Solid–liquid extraction (LSE) |
Mucuna pruriens dehulled and whole seed |
White and black var. utilis |
HCl 0.1 M; solvent: sample ratio 100:1 (v/w | ||||||||
|
HPLC-UV |
Broad bean, cocoa and beans | ), |
10 ng/mL–15 µg/mL |
extraction time 2 × (30 sec under homogenization and 1 h under stirring); extraction temperature 22 °C |
101.8% |
RP-C18 (mean particle diameter 5 µm, 125 × 3 mm I.D.) |
Solvent (A): acetate buffer, pH = 4.66; solvent (B): methanol [35] |
||||
Photodiode array detector (DAD) | It is highly reproducible, rapid and efficient | Sensitivity is rather limited so it is suitable for plant matrices with medium and high concentrations of LD. Selectivity is also limited since it does not allow the unambiguous identification of structurally similar molecules. |
[38] |
Broad bean, cocoa and beans |
|||||||
|
Mucuna pruriens dehulled and whole seed |
// |
Solvent (A): water/methanol/phosphoric acid 975.5:19.5:1 (v/v/v), pH = 2.0; solvent (B): 70% methanol. HClO4 0.2 M, solvent: sample ratio 5:1 (v/w), extraction time 24 h under shaking time for time; extraction temperature 25 °C |
[35] Within-day 84.4–96.0% Between-day 84.0–83.1% |
[38] |
|||||||
|
Vicia faba seeds (cotyledons and embryo axis) |
|||||||||||
|
Vicia faba seeds (cotyledons and embryo axis) | var. Alameda var. Brocal |
HClO4 0.83 mol/kg; |
Ammonium phosphate buffer (0.05 mol/kg, pH = 2.0) solvent: sample ratio 100:1 (v/w); extraction time 1 min under homogenization; extraction temperature 4 °C |
[25 // |
] [25] |
||||||
|
Vicia faba broad beans |
|||||||||||
|
Vicia faba broad beans |
Iambola, San Francesco, FV5, Cegliese, Extra-early purple and Aguadulce supersimonia |
Water (H2O) and acetonitrile (ACN) both containing 0.1% (v/v%) acid formic 5% w/v HClO4 solution; solvent: sample ratio 10:1 (v/w); extraction time 5 min under homogenization; extraction temperature 4 °C |
[39] // |
[39] |
|||||||
|
Vicia Faba roots, sprouts and seeds |
|||||||||||
|
Vicia faba |
// |
roots, sprouts, seedling, leaf, flower, pod, stem Formic acid:ethanol (1:1 v/v); solvent: sample ratio (10–40):1 (v/w); extraction time 5 × 120 min at 120 rpm; extraction temperature 4 °C |
94.1–116.6% |
Solvent (A): 0.1% acetic (98%); [40] |
|||||||
Solvent (B): methanol (2%). |
[43] |
Mucuna pruriens seed cooked and raw |
|||||||||
|
Vicia faba sprouts |
// |
Solvent (A): 82% buffer solution (32 mM citric acid, 54.3 mM sodium acetate, 0.074 mM Na2EDTA, 0.215 mM octyl sulphate pH = 4); Solvent (B): 18% methanol Water; solvent: sample ratio 400:1 (v/w); extraction time 20 min under stirring; extraction temperature 25 °C |
[44 // |
] [41] |
|||||||
|
Avena sativa seeds |
GK Iringo, GK Kormorán and GK Zalán |
||||||||||
|
Mucuna and | Aqueous solution of 0.1% (m/v) ascorbic acid and 1% (v/v) MeOH; solvent: sample ratio 6:1 (v/w); extraction time 5 h under shaking; extraction temperature 25 °C |
95.2–99.6% |
[42] |
||||||||
|
Vicia Faba roots, sprouts, leaf, seediling, pod, flower, stem |
// |
Ethanol solution 95% (v/v); solvent: sample ratio //; extraction time 72 h in freezer; extraction temperature −18 °C |
// |
[43] |
|||||||
Stizolobium pruriens seed |
Solvent (A): 0.1 N acetic acid (90%); Solvent (B): methanol (10%) |
[45] |
Vicia Faba | ||||||||
|
Mucuna pruriens seed |
Solvent (A): 0.1% formic acid (98%); Solvent (B): methanol (2%) |
[48] |
sprouts |
// |
Ethanol solution 95% (v/ | ||||||
|
Vicia faba flowers, fruits and leaves |
Solution of 50 mM potassium dihydrogen phosphate (pH = 2.3) | v); solvent: sample ratio //; |
[36] extraction time 48–72 h; extraction temperature −18 °C |
// |
[44] |
||||||
|
Mucuna and Stizolobium pruriens seed |
M. sempervirens, M. birdwoodiana, M. macrocarpa, M. interrupta, M. paohwashanica, Stizolobium pruriens var. pruriens, S. pruriens var. utilis |
||||||||||
|
Vicia faba sprouts, leaves, flowers, pods, roots |
Solvent (A): water with 0.3% formic acid; Solvent (B): acetonitrile with 0.3% formic acid | HCl 0.1 M; solvent: sample ratio 20:1 (v/w); extraction time 2 × 5–10 min; extraction temperature 100 °C with a steam bath. |
// |
[51] [45] |
|||||||
|
Mucuna pruriens seed |
// |
0.1 M phosphate-buffered solution (pH = 7.0); | |||||||||
|
Vicia faba seeds |
Solvent (A): 97% v/v of an aqueous solution of 0.2% v/v acetic acid; Solvent (B): 3% v/v methanol | solvent: sample ratio 5000:1( v/w); extraction time 5 h; |
[ extraction temperature 25 °C under stirring. |
15] 99.35% |
[46] |
||||||
|
Mucuna pruriens leaves |
// |
||||||||||
|
M. pruriens seeds |
Water, methanol and acetonitrile (5:3:2) containing 0.2% triethylamine, pH = 3.3 | 0.1 M phosphate-buffered solution (pH = 7.0); |
[54 solvent: sample ratio 500:1 (v/w); extraction time 5 h; extraction temperature 25 °C under stirring |
] 98.30% |
|||||||
|
Mucuna pruriens seed |
|||||||||||
|
Mucuna pruriens powder formulation |
// |
Solvent (A): water 80% v/v; Solvent (B): methanol 20% v/v Citric acid 58% (wt%); solvent: sample ratio 7:1; extraction time 90 min; extraction temperature 60 °C |
80–84% |
[ |
[47] |
||||||
] |
Ultrasound-assisted solvent extraction (UASE) |
Mucuna pruriens seed |
Arka Dhanwantri |
Water acidified with 0.1 M HCl (pH: 2.6); | |||||||
|
Mucuna pruriens powder and extracts | solvent: sample ratio 10:1 ( | v / |
RP-C18 (mean particle diameter 5 µm) | w |
Water: Methanol: Acetonitrile (100:60:40) containing 0.2% Triethylamine, pH = 3.3 | ); frequency 35 kHz; extraction time 5, 10, 15 min; extraction temperature 25 °C. |
(5 min) 30.7% |
[55] (10 min) 25.6% (15 min) 31.5% |
[48] |
||
|
Arka Ashwini |
|||||||||||
|
Mucuna sanjappae Seed |
(5 min) 29.0% (10 min) 27.7% (15 min) 26.8% |
||||||||||
|
RP-C18 (250 × 4.6 mm I.D.) |
Methanol |
[53] |
White |
(5 min) 29.3% (10 min) 31.4% (15 min) 30.8% |
|||||||
|
Brown |
(5 min) 23.9% (10 min) 28.7% (15 min) 30.6% |
||||||||||
|
Vicia faba sprouts and seeds |
// |
MeOH and water mixture (80:20); solvent: sample ratio 1:5 (v/w); frequency //; extraction time 30 min; extraction temperature 25 °C. |
// |
[49] |
|||||||
|
Mucuna utilis seed |
RP-C18 (250 × 4.0 mm I.D.) |
Solvent (A):0.5% v/v of acetic acid 30%; Solvent (B): methanol 70% |
[59] |
||||||||
|
LC-MS |
Vicia faba roots, sprouts and seeds |
18 µg/Kg |
RP-C18 (mean particle diameter 2.6 µm, 100 × 4.6 mm I.D.) |
Solvent (A): ultrapure water with 0.5% (v/v) formic acid 50%; Solvent (B): methanol 50% |
Photo diode array detector (DAD) and triple quadrupole (TQ) mass spectrometer |
Robust analytical technique that provides higher sensitivity and selectivity than LC-UV methods. It allows to unambiguously identify the compounds under analysis, through the possibility of fragmentation. |
It is a technique susceptible to matrix effects: co-eluting compounds could interfere with the ionization of the analyte under examination. Detection in MRM mode is to be preferred. |
[40] |
|||
|
0.01 µg/mL |
Not reported |
Not reported |
Photo diode array detector (DAD) and quadrupole-time-of-flight (QTOF)- mass spectrometer |
[49] |
Vicia faba flowers, fruits and leaves |
||||||
|
Avena sativa seeds | // |
RP-C18 (mean particle diameter 4 µm, 250 × 2 mm I.D.) Water boiling deionized; solvent: sample ratio 50:1 (v/w); frequency //; extraction time 15 min; extraction temperature 100 °C. |
Solvent (A): solution 0.1% ( 100.32% |
v/v) of formic acid (97%); Solvent (B): ACN/MeOH 75/25 containing 0.1% (v/v) formic acid (3%) |
Ion Trap mass spectrometer [36] |
||||||
[ | ] |
Vicia Faba seeds |
// |
HCl 10 mM 5 mL; | |||||||
|
HPTLC |
M. pruriens seeds | solvent: sample ratio // ( v/w); frequency //; extraction time 2 × 60 min; extraction temperature 25 °C. |
Not reported 99.8% |
Silica-coated aluminum sheet (10 cm × 10 cm with 0.2 mm thickness) |
[50] |
||||||
n-butanol, | acetic acid and water were used as mobile phase at 4:1:1 |
UV-Vis thin layer scanner |
It makes it possible to obtain a preliminary separation of the analytes in a fast, efficient, easy and low cost analysis |
It is generally employed only for qualitative analysis. It is poorly reproducible, as it works in an open system, whose environmental conditions could alter the results. |
[54] |
Lens culinaris seeds |
105.0% |
||||
|
CE-UV |
Vicia faba seeds |
LOD value 0.7 µg/mL. |
47 cm (40 cm from inlet to the detector) × 75 µm i.d. fused-silica capillary |
35 mM NaH2PO4, pH = 4.55, 17.5 kV and 30 °C. |
Photo diode array detector (DAD) |
It allows faster analysis and higher efficiency than LC-UV |
It is less sensitive than HPLC-UV |
[50] |
Vicia Faba sprouts, leaves, flowers, pods, roots |
// |
|
|
Lens culinaris seeds | Aqueous MeOH 50% (v/v); solvent: sample ratio 200:1 (v/w); frequency //; extraction time 30 min; extraction temperature below 40 °C. |
// |
[51] |
||||||||
|
Vicia faba seeds |
Bachus, Bolero White, Windsor Bonus, Rambo Amigo, Olga Granit, Albus Fernando, Amulet |
Aqueous CH3COOH 0.2% (v/v); solvent: sample ratio 25:1 (v/w); frequency 40 kHz; extraction time 2 × 20 min; extraction temperature 25 °C. |
|||||||||
|
Electrochemical methods |
Mucuna pruriens seed, leaves |
LOD value 1.54 µM |
// |
Working electrode: [15] |
|||||||
gold modified pencil graphite | Supporting electrolyte: |
0.1 M phosphate-buffered solution (pH = 7.0) |
Differential pulse voltammetry (DPV) |
It makes it possible to identify and quantify the analyte in a fast and economical way, through the use of conventional or modified nanostructured electrodes, which permits a better selectivity and sensitivity of analysis. |
The technique still shows limitations especially related to the problem of electrode poisoning and oxidizable interfering compounds in the same range of anode potential. |
[46] |
Wild type legume grain |
Acacia nilotica, Bauhinia purpurea, Canavalia ensiformis, Cassia hirsuta, Caesalpinia bonducella, Erythrina indica, Mucuna gigantea, Pongamia pinnata, Sebania sesban, Xylia xylocarpa |
HCl 0.1 M; solvent: sample ratio 10:1 ( | ||
|
Mucuna pruriens seed cooked and raw |
LOD value 5.12 ng/mL | v /w); frequency // kHz; extraction time 30 min and stirring for 1 h. extraction temperature 25 °C. |
RP-C18 (mean particle diameter 3.5 µm, 150 × 2.1 mm I.D.) Working electrode: Glassy carbon // |
Eluent/supporting electrolyte: 103 mM sodium acetate, 0.88 mM citric acid, 2.14 mM 1-octanesulphonic acid sodium salt with pH adjusted to 2.38 by orthophosphoric acid [52] |
|||||||
Amperometric detection at a potential of +0.7 V after micro-high performance | liquid chromatography separation |
[41] |
Mucuna sanjappae seed |
// |
HCl 0.1 M; solvent: sample ratio 300:1 (v | ||||||
|
Sunflower seed, sesame seed, pumpkin seed and fava bean seed |
LOD value 14.3 nmol/L | / w); frequency // kHz; |
Working electrode: glassy carbon modified by graphene quantum dots decorated with Fe3O extraction time 20 min; extraction temperature 25 °C. |
4 // |
nanoparticles/functionalized multiwalled carbon [53] |
||||||
nanotubes | Supporting electrolyte: 0.1 mol/L PBS at pH = 5.5 |
Differential Pulse Voltammetry (DPV) |
M. pruriens seeds |
Macrocarpa |
HCl 0.1 M; solvent: sample ratio 300:1 (v/w); frequency // kHz; | ||||||
|
Sweet potato |
17 nM |
extraction time 20 min; extraction temperature 25 °C. |
Working electrode: nitrogen-doped graphene supported with nickel oxide nanocomposite // |
[54] |
|||||||
Supporting electrolyte: 0.05 M PBS at pH = 7 | Differential pulse voltammetry (DPV) |
Microwave-assisted solvent extraction (MASE) |
|||||||||
|
Spectrophotometry UV-Vis |
Mucuna pruriens seed |
Wild type legume grain Arka Dhanwantri |
LOD 1.12 µg/mL |
Water acidified with 0.1 M HCl (pH = 2.6); solvent: sample ratio 10:1 (v/w); MW power 400 W; irradiation time 5, 10, 15 min; extraction temperature 60 °C. |
// |
(5 min) 53.5% (10 min) 58.7% (15 min) 58.4% |
// |
[48] |
|||
// | It is an easy to use and low-cost technique that allows both qualitative and quantitative evaluation | It is generally not preceded by a separation step. This implies that the sample can contain interfering compounds causing potential false positives. |
[52] |
Arka Ashwini |
|||||||
|
Phaseolus vulgaris dried seed, seeding and callus |
(5 min) 50.6% (10 min) 59.6% (15 min) 54.0% |
||||||||||
[58] |
White |
||||||||||
|
NMR | (5 min) 50.5% (10 min) 49.6% (15 min) 58.5% |
||||||||||
Mucuna pruriens seed |
LOD value 0.0175 mg/g |
// |
// |
// |
It is a highly reproducible technique. It makes it possible to get structural details of the compounds under examination. |
Requires expensive equipment and provides low sensitivity compared to LC-MS. It is hardly used for quantification, due to the chemical noise and signal overlapping. |
[56] |
Brown |
(5 min) 56.1% (10 min) 54.9% (15 min) 54.8% |
||
|
Reflux extraction |
Mucuna pruriens seed |
Arka Dhanwantri |
Water acidified with 0.1 M HCl (pH = 2.6); solvent: sample ratio 10:1 (v/w); extraction time 300 min, extraction temperature 100 °C |
60.2% |
[48] |
||||||
|
Arka Ashwini |
65.7% |
||||||||||
|
White |
57.2% |
||||||||||
|
Brown |
59.8% |
||||||||||
|
Mucuna pruriens powder and extracts |
// |
MeOH and 0.1 M HCl mixture (70:30); solvent: sample ratio 100:1 (v/w); extraction time 30 min; extraction temperature 25 °C |
98.83% |
[55] |
|||||||
|
Mucuna pruriens seed |
Preta Kaunch |
HCl 0.1 M; solvent: sample ratio 2:1 (v/w); extraction time 180 min; extraction temperature 25 °C |
98.1–106.7% |
[56] |
|||||||
|
Maceration |
Mucuna pruriens powder formulation |
// |
Water and EtOH mixture (30:70); solvent: sample ratio //; extraction time 7 days; extraction temperature cold |
94.5% |
[57] |
||||||
|
Phaseolus vulgaris dried seed, seeding and callus |
// |
HCl 0.1 M and EtOH mixture (1:1); solvent: sample ratio 1:10; extraction time 5 days; extraction temperature 25 °C |
99.55–100.27% |
[58] |
|||||||
|
Soxhlet extraction |
Mucuna utilis seed |
// |
MeOH; solvent: sample ratio //; extraction Soxhlet time //; extract obtained sonication for 60 min with 100 mL HCl 0.1 M; extraction temperature 25 °C |
98.67–100.4% |
[59] |
// This indicates that values were not reported.
// This indicates that values were not reported.