Despite the fact that the incidence of gastric cancer has declined over the last decade, it is still the world’s leading cause of cancer-related death. The diagnosis of early gastric cancer is difficult, as symptoms of this cancer only manifest at a late stage of cancer progression. Thus, the prognosis of gastric cancer is poor, and the current treatment for improving patients’ outcomes involves the application of surgery and chemotherapy. Immunotherapy is one of the most recent therapies for gastric cancer, whereby the immune system of the host is programmed to combat cancer cells, and the therapy differs based upon the patient’s immune system. However, an understanding of the role of immune cells, namely the cell-mediated immune response and the humoral immune response, is pertinent for applications of immunotherapy. The roles of immune cells in the prognosis of gastric cancer have yielded conflicting results.
- gastric cancer
- immune cells
- immune response
- T cells
- B cells
- macrophages
- natural killer cells
- dendritic cells
1. Introduction
2. Functions of Immune Cells in Gastric Cancer
2.1. T Cells
T cells have different categories: cytotoxic T cells (CD8+ cells), T helper cells (CD4+ cells), and regulatory T cells. However, there are T cell categories that are yet to be characterized or discovered for their functions. Several types of T helper cells, based on their cytokine secretion, include T helper cell 1 (Th1), T helper cell 2 (Th2), T helper cell 9 (Th9), T helper cell 17 (Th17), T helper cell 22 (Th22), and T helper cells with high levels of expression for forkhead box 3 (FOXP3) and CD25 [52][16]. For instance, Th1 produces interferon-gamma (IFN-γ) and interleukin-2 (IL-2), while Th2 produces interleukin-4 (IL-4), interleukin-5 (IL-5), interleukin-6 (IL-6), and interleukin-10 (IL-10). Those cytokines play an essential role in activating cell-mediated immunity and orchestrating B-cell mediated humoral immunity [53][17]. Th9 cells secrete interleukin-9 (IL-9), which involves the orchestration of cell-mediated immunity [54][18], while Th17 cells produce interleukin-17 (IL-17), interleukin-21 (IL-21), and interleukin-22 (IL-22), which are pertinent in combating extracellular pathogens [55][19]. On the other hand, Th22 secretes interleukin-22 (IL-22), which is an essential activator of innate immunity in epithelial cells [56][20]. Generally, the functions of T cytotoxic cells include the neutralization of host cells that are infected with intracellular pathogens, such as viruses and some bacteria, and killing cancer cells. In contrast, the function of T helper cells primarily involves the crucial neutralization of cells that have been infected with intracellular pathogens through the mediation of B cells to produce antibodies, the activation of macrophages for bactericidal activity, and the facilitation of other immune cells’ activities in orchestrating immune defense. Conflicting reports have been made with regard to the role of T cytotoxic cells in the carcinogenesis of gastric cancer (Figure 1). At one end of the spectrum, the distinct expression of T cytotoxic cells has been associated with a different prognosis of gastric cancer, in which a high density of T cells in patients with gastric cancer is associated with a poor prognosis of gastric cancer. A study conducted by Thompson et al. [57][21] showed that patients with a poor prognosis demonstrated a higher infiltration of CD8+ T cells in gastric tumors compared to patients with a better prognosis. They also found that gastric cancer cells that had been isolated from patients with a poor prognosis also expressed high levels of programmed cell death ligand 1 (PDL-1), which is a critical ligand that deactivates T cells via the stimulation of T regulatory cells, and subsequently encourages autoimmunity and the incidence of autoimmune diseases [58][22]. Of note, a high expression of CD8+ T cells has been shown in mixtures of cells that have been isolated from the tumors of patients with gastric cancer who were infected with the Epstein–Barr virus (EBV) [59][23]. At the other end of the spectrum, a study conducted by Lee et al. [60][24] showed that a high level of infiltration by CD8+ T cells in patients with gastric cancer was associated with a better rate of survival than for patients with a low degree of infiltration by CD8+ T cells. In the mouse model, a high infiltration of CD8+ T cells, with the absence of CD4+ T cells, has been associated with severe gastritis. This finding concludes that CD4+ T cells are vital for the surveillance of CD8+ T cell activity [61][25].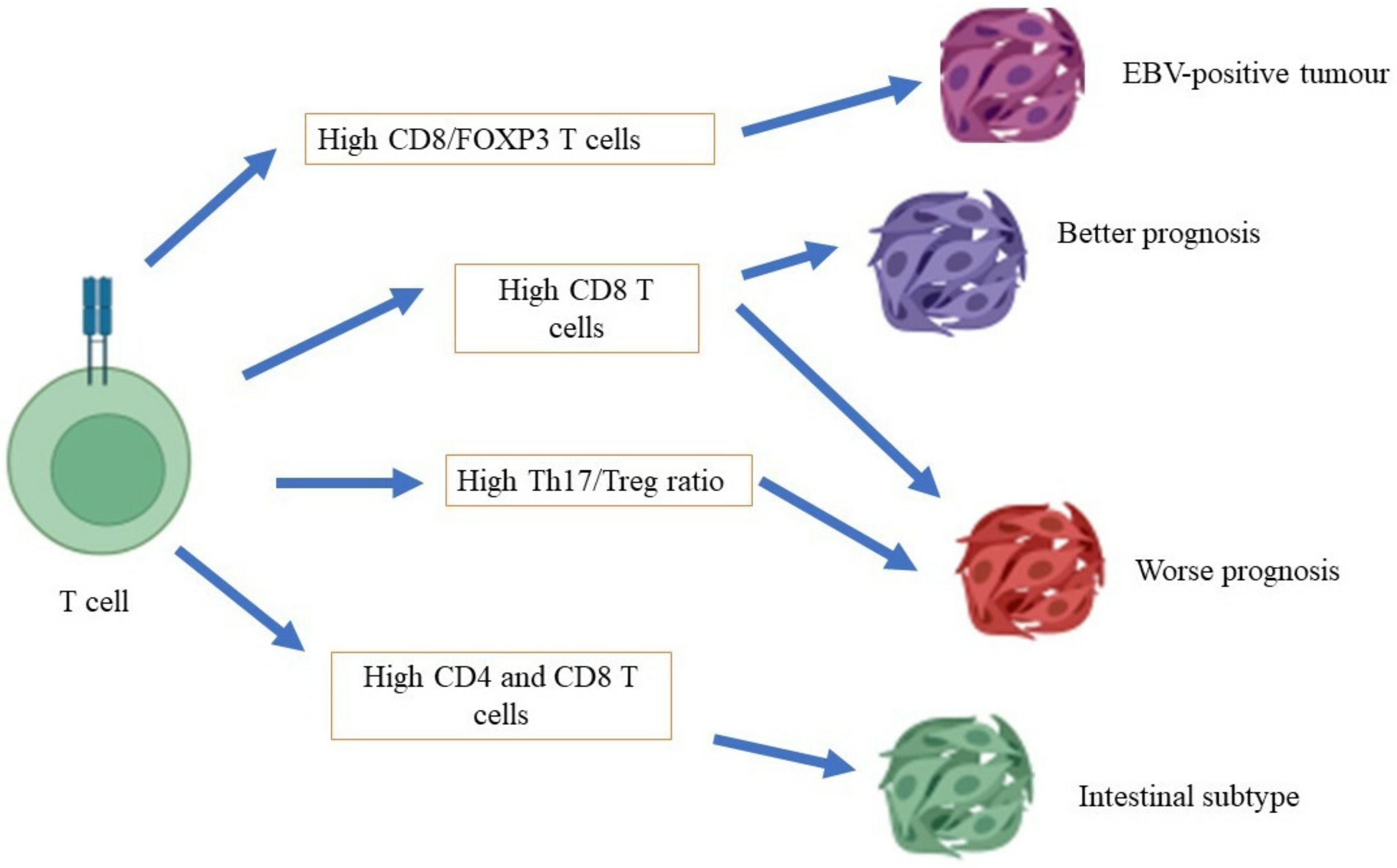
2.2. B Cells
B cells function as antigen-presenting cells, and they can differentiate into plasma cells to produce antibodies and to secrete cytokines such as IL-6, IFN-γ, and tumor necrosis α (TNF-α), for the development of CD4+ effector and memory T cells [70][34]. A subset of B regulatory cells (Breg) has been shown to inhibit antitumoral activity that is mediated by T cells and other immune cells, namely effector T cells, natural killer cells, and tumor-associated macrophages [71][35]. A comprehensive immunophenotype of gastric cancer cells using an antibody microarray showed a higher expression of B-cell markers, including CD19, CD38, and surface immunoglobulin (sIg), in patients with H. pylori-infected gastric cancer than in patients without infections [72][36].2.3. Macrophages
Macrophages are phagocytes that play a pivotal role in the clearance of erythrocytes and tissue modeling [73][37]. Furthermore, these cells also have an essential role in recognizing pathogens via receptor recognition and the neutralization of apoptotic cells [74][38]. A high density of macrophages is associated with a low rate of survival in patients with gastric cancer and increased gastric tumor invasiveness through the β-catenin pathway (Figure 2) [75][39]. A high density of tumor-associated macrophages (TAM) of phenotype M2 was found in patients with gastric cancer with invasive gastric peritoneum compared to patients without invasive gastric peritoneum. Additionally, a gastric cancer cell line co-cultured with the TAM phenotype M2 also showed increased degrees of invasiveness and cell division in the xenographic model [76][40]. Osteopontin, a protein that is involved in inflammation and cancer pathogenesis [77][41], recruits macrophages into the tumor microenvironment and polarizes macrophages to the M2 phenotype, which facilitates gastric cancer development [78][42]. On the other hand, macrophages induce invasiveness, migration, and gastric cancer cell movement through epidermal growth factor and the phosphorylation of Akt, Erk ½, and c-Src [79][43]. H. pylori-infected macrophages demonstrated the activation of immune-regulated genes such as Cd44, Cd40, Cd86, and Cd274, and the inhibition of macrophage cell line division, primary macrophage cells, and the transcription of genes that are involved in DNA synthesis and the cell cycle [80][44].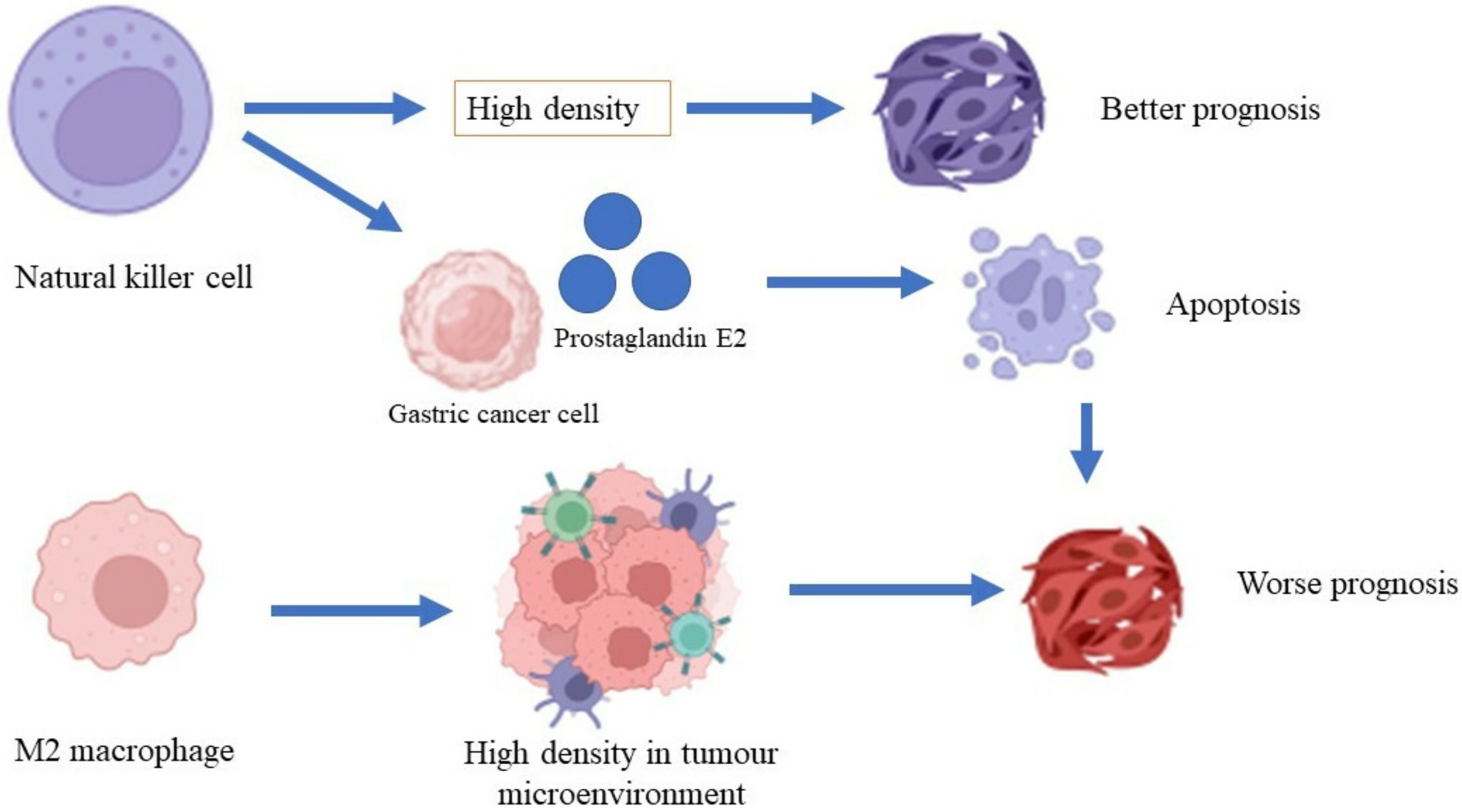
2.4. Natural Killer Cells
Natural killer cells are lymphocytes that are important in mediating innate immunity and adaptive immunity for the surveillance of the immune system against invading intracellular viruses and cancer cells [81][45]. Natural killer cells can recognize cancer cells because they express natural killer group-2 (NKG2D), which is expressed by cancer cells [82][46]. The expressions of CD antigens such as CD56 and CD16 play essential roles in mediating the activation of immune cells, including macrophages, T cells, and dendritic cells [83][47]. A study conducted by Ishigami et al. [84][48] revealed that a high density of natural killer cells in patients with gastric cancer is associated with a better prognosis of gastric cancer compared to a low density of natural killer cells. A lysis activity that is greater than 25% in natural killer cells is significantly associated with a better five-year survival rate compared to lysis activity that is less than 25% [85][49]. A high apoptosis rate for natural killer cells was shown in patients with advanced-stage gastric cancer. However, the rate of apoptosis was reduced after the patients underwent gastrectomy [86][50]. Gastric cancer cells produce prostaglandin E2, which inhibits natural killer cell division and encourages apoptosis [87][51].2.5. Dendritic Cells
Dendritic cells, or antigen-presenting cells, are master regulators of innate immunity and adaptive immunity because they can present antigens to major histocompatibility complex (MHC) class I and MHC class II. Immature dendritic cells express low levels of CD54, CD58, CD80, CD86, CD40, CD25, CD83, and IL-12, whereas mature dendritic cells express those CD antigens at high levels. IL-12 is an essential cytokine for activating natural killer cells, T cells, and B cells [88][52]. There are several types of dendritic cells, including myeloid dendritic cells, plasmacytoid dendritic cells, Langerhans cells, microglia cells, and CD14+ dendritic cells, which are based on the cell markers that are expressed on their surfaces [89][53]. In ovarian cancer cells, dendritic cells express high levels of B7-H1 and PD-1, which inhibit T-cell activation and deactivate the function of dendritic cells in recognizing tumors [90][54]. Factors that are secreted by tumor cells inhibit the differentiation of dendritic cells, which deactivates their antitumoral properties [91][55]. Furthermore, factors that are produced by tumor cells leads to the high expression of CD11b, which reduces the expression of MHC class II and is associated with a worse prognosis in patients with gastric cancer [92][56].3. Immune Checkpoints in Gastric Cancer
Immune checkpoints are essential to maintain the self-tolerance of the immune system to body cells and prevent autoimmune reactions. Tumour cells exploit the immune checkpoint molecules to their advantage by downregulating the surveillance of immune cells toward the cancer cells [93][57]. Immune checkpoint molecules are often expressed on immune cells as master regulators to keep the immune system in check. Immune checkpoint molecules that have been extensively studied include programmed cell death receptor-1 (PD-1), programmed cell death receptor ligand-1 (PD-L1), programmed cell death receptor ligand-2 (PD-L2), lymphocyte activation-gene-3 (LAG-3), cytotoxic T lymphocyte antigen-4 (CTLA-4), B and T lymphocyte attenuator (BTLA) [94][58]. Furthermore, an immune checkpoint score based on twenty immune checkpoints has been studied and can be applied to predict the prognosis of gastric cancer patients [95][59]. Cancer cells express PD-L1 to escape immune surveillance through binding to PD-1 expressed on activated immune cells, namely T cells, monocytes, macrophages, and natural killer cells. The binding inhibits paramount immune signal transduction pathways and renders immune cells to be unresponsive to cancer cells and causes the downregulation of immune cell proliferation, cytokine secretion, and activation [96][60]. Treatments of gastric cancer include curative resection and adjuvant chemotherapy depending on the staging of gastric cancer [97][61]. Immunotherapy through the identification of immune checkpoints that can be manipulated to attack cancer cells has emerged as an alternative treatment for gastric cancer. Immune checkpoint inhibitors have been introduced to activate immune cells in gastric cancer patients as part of immunotherapy. Several monoclonal antibodies, namely nivolumab and pembrolizumab, that target the PD-1/PD-L1 immune checkpoint have undergone clinical trial phases with promising results. In a phase 3 clinical trial conducted among gastric cancer patients at an advanced stage from East Asian countries (Japan, South Korea, and Taiwan), the administration of nivolumab increased median follow-up in surviving patients, median overall survival, and the 1-year overall survival rate in the nivolumab group compared to the placebo group. However, adverse events were more frequently observed in the group that received nivolumab than placebo [98][62]. Another monoclonal antibody that targets the PD-1/PD-L1 checkpoint is pembrolizumab. A phase 1b clinical trial conducted among advanced gastric cancer patients who were positive for PD-L1 found that pembrolizumab had manageable toxicity safety among patients and elicited antitumoral activity against gastric cancer [99][63]. Another drug that targets the PD-1 checkpoint is avelumab. A phase 3 clinical trial that involved advanced gastric cancer patients found that avelumab did not improve overall survival and progression-free survival of the patients compared to chemotherapy, although the toxicity of avelumab was more manageable than chemotherapy [100][64]. A combination of monoclonal antibodies that target PD-1 (nivolumab) and CTLA-4 (ipilimumab) have also been used in clinical trials. Recent global phase 3 clinical trials (CheckmMate 649 study) that compared the combination of nivolumab and ipilimumab to that of nivolumab and chemotherapy revealed that the combination of nivolumab and chemotherapy resulted in better overall survival of the patients than that of nivolumab and ipilimumab [101][65].References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249.
- Yusefi, A.R.; Bagheri Lankarani, K.; Bastani, P.; Radinmanesh, M.; Kavosi, Z. Risk factors for gastric cancer: A systematic review. Asian Pac. J. Cancer Prev. 2018, 19, 591–603.
- Necula, L.; Matei, L.; Dragu, D.; Neagu, A.I.; Mambet, C.; Nedeianu, S.; Bleotu, C.; Diaconu, C.C.; Chivu-Economescu, M. Recent advances in gastric cancer early diagnosis. World J. Gastroenterol. 2019, 25, 2029–2044.
- Orditura, M.; Galizia, G.; Sforza, V.; Gambardella, V.; Fabozzi, A.; Laterza, M.M.; Andreozzi, F.; Ventriglia, J.; Savastano, B.; Mabilia, A.; et al. Treatment of gastric cancer. World J. Gastroenterol. 2014, 20, 1635–1649.
- Swann, J.B.; Smyth, M.J. Immune surveillance of tumors. J. Clin. Investig. 2007, 117, 1137–1146.
- Airoldi, I.; Di Carlo, E.; Cocco, C.; Sorrentino, C.; Fais, F.; Cilli, M.; D’Antuono, T.; Colombo, M.P.; Pistoia, V. Lack of Il12rb2 signaling predisposes to spontaneous autoimmunity and malignancy. Blood 2005, 106, 3846–3853.
- Clementi, R.; Locatelli, F.; Dupré, L.; Garaventa, A.; Emmi, L.; Bregni, M.; Cefalo, G.; Moretta, A.; Danesino, C.; Comis, M.; et al. A proportion of patients with lymphoma may harbor mutations of the perforin gene. Blood 2005, 105, 4424–4428.
- Mittal, D.; Gubin, M.M.; Schreiber, R.D.; Smyth, M.J. New insights into cancer immunoediting and its three component phases--elimination, equilibrium and escape. Curr. Opin. Immunol. 2014, 27, 16–25.
- Dighe, A.S.; Richards, E.; Old, L.J.; Schreiber, R.D. Enhanced in vivo growth and resistance to rejection of tumor cells expressing dominant negative IFN gamma receptors. Immunity 1994, 1, 447–456.
- Shankaran, V.; Ikeda, H.; Bruce, A.T.; White, J.M.; Swanson, P.E.; Old, L.J.; Schreiber, R.D. IFNgamma and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature 2001, 410, 1107–1111.
- Müller-Hermelink, N.; Braumüller, H.; Pichler, B.; Wieder, T.; Mailhammer, R.; Schaak, K.; Ghoreschi, K.; Yazdi, A.; Haubner, R.; Sander, C.A.; et al. TNFR1 signaling and IFN-gamma signaling determine whether T cells induce tumor dormancy or promote multistage carcinogenesis. Cancer Cell 2008, 13, 507–518.
- Teng, M.W.; Galon, J.; Fridman, W.H.; Smyth, M.J. From mice to humans: Developments in cancer immunoediting. J. Clin. Investig. 2015, 125, 3338–3346.
- Matsueda, S.; Graham, D.Y. Immunotherapy in gastric cancer. World J. Gastroenterol. 2014, 20, 1657–1666.
- Slamon, D.J.; Leyland-Jones, B.; Shak, S.; Fuchs, H.; Paton, V.; Bajamonde, A.; Fleming, T.; Eiermann, W.; Wolter, J.; Pegram, M.; et al. Use of chemotherapy plus a monoclonal antibody against HER2 for metastatic breast cancer that overexpresses HER2. N. Engl. J. Med. 2001, 344, 783–792.
- Jiang, J.; Xu, N.; Wu, C.; Deng, H.; Lu, M.; Li, M.; Xu, B.; Wu, J.; Wang, R.; Xu, J.; et al. Treatment of advanced gastric cancer by chemotherapy combined with autologous cytokine-induced killer cells. Anticancer Res. 2006, 26, 2237–2242.
- Bailey, S.R.; Nelson, M.H.; Himes, R.A.; Li, Z.; Mehrotra, S.; Paulos, C.M. Th17 cells in cancer: The ultimate identity crisis. Front. Immunol. 2014, 5, 276.
- Larussa, T.; Leone, I.; Suraci, E.; Imeneo, M.; Luzza, F. Helicobacter pylori and T Helper Cells: Mechanisms of Immune Escape and Tolerance. J. Immunol. Res. 2015, 2015, 981328.
- Kaplan, M.H. Th9 cells: Differentiation and disease. Immunol. Rev. 2013, 252, 104–115.
- Eisenstein, E.M.; Williams, C.B. The T(reg)/Th17 cell balance: A new paradigm for autoimmunity. Pediatr Res. 2009, 65, 26R–31R.
- Eyerich, S.; Eyerich, K.; Pennino, D.; Carbone, T.; Nasorri, F.; Pallotta, S.; Cianfarani, F.; Odorisio, T.; Traidl-Hoffmann, C.; Behrendt, H.; et al. Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling. J. Clin. Investig. 2009, 119, 3573–3585.
- Thompson, E.D.; Zahurak, M.; Murphy, A.; Cornish, T.; Cuka, N.; Abdelfatah, E.; Yang, S.; Duncan, M.; Ahuja, N.; Taube, J.M.; et al. Patterns of PD-L1 expression and CD8 T cell infiltration in gastric adenocarcinomas and associated immune stroma. Gut 2017, 66, 794–801.
- Francisco, L.M.; Sage, P.T.; Sharpe, A.H. The PD-1 pathway in tolerance and autoimmunity. Immunol. Rev. 2010, 236, 219–242.
- Kuzushima, K.; Nakamura, S.; Nakamura, T.; Yamamura, Y.; Yokoyama, N.; Fujita, M.; Kiyono, T.; Tsurumi, T. Increased frequency of antigen-specific CD8(+) cytotoxic T lymphocytes infiltrating an Epstein-Barr virus-associated gastric carcinoma. J. Clin. Investig. 1999, 104, 163–171.
- Lee, H.E.; Chae, S.W.; Lee, Y.J.; Kim, M.A.; Lee, H.S.; Lee, B.L.; Kim, W.H. Prognostic implications of type and density of tumour-infiltrating lymphocytes in gastric cancer. Br. J. Cancer 2008, 99, 1704–1711.
- Tan, M.P.; Pedersen, J.; Zhan, Y.; Lew, A.M.; Pearse, M.J.; Wijburg, O.L.; Strugnell, R.A. CD8+ T cells are associated with severe gastritis in Helicobacter pylori-infected mice in the absence of CD4+ T cells. Infect. Immun. 2008, 76, 1289–1297.
- Smythies, L.E.; Waites, K.B.; Lindsey, J.R.; Harris, P.R.; Ghiara, P.; Smith, P.D. Helicobacter pylori-induced mucosal inflammation is Th1 mediated and exacerbated in IL-4, but not IFN-gamma, gene-deficient mice. J. Immunol. 2000, 165, 1022–1029.
- Akhiani, A.A.; Pappo, J.; Kabok, Z.; Schön, K.; Gao, W.; Franzén, L.E.; Lycke, N. Protection against Helicobacter pylori infection following immunization is IL-12-dependent and mediated by Th1 cells. J. Immunol. 2002, 169, 6977–6984.
- Yang, P.; Qiu, G.; Wang, S.; Su, Z.; Chen, J.; Wang, S.; Kong, F.; Lu, L.; Ezaki, T.; Xu, H. The mutations of Th1 cell-specific T-box transcription factor may be associated with a predominant Th2 phenotype in gastric cancers. Int. J. Immunogenet. 2010, 37, 111–1115.
- Corthay, A. How do regulatory T cells work? Scand. J. Immunol. 2009, 70, 326–336.
- Ichihara, F.; Kono, K.; Takahashi, A.; Kawaida, H.; Sugai, H.; Fujii, H. Increased populations of regulatory T cells in peripheral blood and tumor-infiltrating lymphocytes in patients with gastric and esophageal cancers. Clin. Cancer Res. 2003, 9, 4404–4408.
- Kim, K.J.; Lee, K.S.; Cho, H.J.; Kim, Y.H.; Yang, H.K.; Kim, W.H.; Kang, G.H. Prognostic implications of tumor-infiltrating FoxP3+ regulatory T cells and CD8+ cytotoxic T cells in microsatellite-unstable gastric cancers. Hum. Pathol. 2014, 45, 285–293.
- Nagase, H.; Takeoka, T.; Urakawa, S.; Morimoto-Okazawa, A.; Kawashima, A.; Iwahori, K.; Takiguchi, S.; Nishikawa, H.; Sato, E.; Sakaguchi, S.; et al. ICOS+ Foxp3+ TILs in gastric cancer are prognostic markers and effector regulatory T cells associated with Helicobacter pylori. Int. J. Cancer 2017, 140, 686–695.
- Li, Q.; Li, Q.; Chen, J.; Liu, Y.; Zhao, X.; Tan, B.; Ai, J.; Zhang, Z.; Song, J.; Shan, B. Prevalence of Th17 and Treg cells in gastric cancer patients and its correlation with clinical parameters. Oncol. Rep. 2013, 30, 1215–1222.
- Shen, P.; Fillatreau, S. Antibody-independent functions of B cells: A focus on cytokines. Nat. Rev. Immunol. 2015, 15, 441–451.
- Schwartz, M.; Zhang, Y.; Rosenblatt, J.D. B cell regulation of the anti-tumor response and role in carcinogenesis. J. Immunother Cancer 2016, 4, 40.
- Sukri, A.; Hanafiah, A.; Kosai, N.R.; Mohamed Taher, M.; Mohamed Rose, I. Surface Antigen Profiling of Helicobacter pylori-Infected and -Uninfected Gastric Cancer Cells Using Antibody Microarray. Helicobacter 2016, 21, 417–427.
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969.
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623.
- Wu, M.H.; Lee, W.J.; Hua, K.T.; Kuo, M.L.; Lin, M.T. Macrophage infiltration induces gastric cancer invasiveness by activating the β-catenin pathway. PLoS ONE 2015, 10, e0134122.
- Yamaguchi, T.; Fushida, S.; Yamamoto, Y.; Tsukada, T.; Kinoshita, J.; Oyama, K.; Miyashita, T.; Tajima, H.; Ninomiya, I.; Munesue, S.; et al. Tumor-associated macrophages of the M2 phenotype contribute to progression in gastric cancer with peritoneal dissemination. Gastric Cancer 2016, 19, 1052–1065.
- Lund, S.A.; Giachelli, C.M.; Scatena, M. The role of osteopontin in inflammatory processes. J. Cell Commun. Signal 2009, 3, 311–322.
- Lin, C.N.; Wang, C.J.; Chao, Y.J.; Lai, M.D.; Shan, Y.S. The significance of the co-existence of osteopontin and tumor-associated macrophages in gastric cancer progression. BMC Cancer 2015, 15, 128.
- Cardoso, A.P.; Pinto, M.L.; Pinto, A.T.; Oliveira, M.I.; Pinto, M.T.; Gonçalves, R.; Relvas, J.B.; Figueiredo, C.; Seruca, R.; Mantovani, A.; et al. Macrophages stimulate gastric and colorectal cancer invasion through EGFR Y(1086), c-Src, Erk1/2 and Akt phosphorylation and smallGTPase activity. Oncogene 2014, 33, 2123–2133.
- Tan, G.M.; Looi, C.Y.; Fernandez, K.C.; Vadivelu, J.; Loke, M.F.; Wong, W.F. Suppression of cell division-associated genes by Helicobacter pylori attenuates proliferation of RAW264.7 monocytic macrophage cells. Sci. Rep. 2015, 5, 11046.
- Cerwenka, A.; Lanier, L.L. Natural killer cells, viruses and cancer. Nat. Rev. Immunol. 2001, 1, 41–49.
- Nausch, N.; Cerwenka, A. NKG2D ligands in tumor immunity. Oncogene 2008, 27, 5944–5958.
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510.
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer 2000, 88, 577–583.
- Takeuchi, H.; Maehara, Y.; Tokunaga, E.; Koga, T.; Kakeji, Y.; Sugimachi, K. Prognostic significance of natural killer cell activity in patients with gastric carcinoma: A multivariate analysis. Am. J. Gastroenterol. 2001, 96, 574–578.
- Saito, H.; Takaya, S.; Osaki, T.; Ikeguchi, M. Increased apoptosis and elevated Fas expression in circulating natural killer cells in gastric cancer patients. Gastric Cancer 2013, 16, 473–479.
- Li, T.; Zhang, Q.; Jiang, Y.; Yu, J.; Hu, Y.; Mou, T.; Chen, G.; Li, G. Gastric cancer cells inhibit natural killer cell proliferation and induce apoptosis via prostaglandin E2. Oncoimmunology 2015, 5, e1069936.
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252.
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 15, 3–20.
- Krempski, J.; Karyampudi, L.; Behrens, M.D.; Erskine, C.L.; Hartmann, L.; Dong, H.; Goode, E.L.; Kalli, K.R.; Knutson, K.L. Tumor-infiltrating programmed death receptor-1+ dendritic cells mediate immune suppression in ovarian cancer. J. Immunol. 2011, 186, 6905–6913.
- Hargadon, K.M. Tumor-altered dendritic cell function: Implications for anti-tumor immunity. Front. Immunol. 2013, 4, 192.
- Okita, Y.; Tanaka, H.; Ohira, M.; Muguruma, K.; Kubo, N.; Watanabe, M.; Fukushima, W.; Hirakawa, K. Role of tumor-infiltrating CD11b+ antigen-presenting cells in the progression of gastric cancer. J. Surg. Res. 2014, 186, 192–200.
- He, X.; Xu, C. Immune checkpoint signaling and cancer immunotherapy. Cell Res. 2020, 30, 660–669.
- McBride, M.A.; Patil, T.K.; Bohannon, J.K.; Hernandez, A.; Sherwood, E.R.; Patil, N.K. Immune checkpoints: Novel therapeutic targets to attenuate sepsis-induced immunosuppression. Front. Immunol. 2021, 11, 624272.
- Wang, J.B.; Li, P.; Liu, X.L.; Zheng, Q.L.; Ma, Y.B.; Zhao, Y.J.; Xie, J.W.; Lin, J.X.; Lu, J.; Chen, Q.Y.; et al. An immune checkpoint score system for prognostic evaluation and adjuvant chemotherapy selection in gastric cancer. Nat. Commun. 2020, 11, 6352.
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742.
- Joshi, S.S.; Badgwell, B.D. Current treatment and recent progress in gastric cancer. CA Cancer J. Clin. 2021, 71, 264–279.
- Kang, Y.K.; Boku, N.; Satoh, T.; Ryu, M.H.; Chao, Y.; Kato, K.; Chung, H.C.; Chen, J.S.; Muro, K.; Kang, W.K.; et al. Nivolumab in patients with advanced gastric or gastro-oesophageal junction cancer refractory to, or intolerant of, at least two previous chemotherapy regimens (ONO-4538-12, ATTRACTION-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 390, 2461–2471.
- Muro, K.; Chung, H.C.; Shankaran, V.; Geva, R.; Catenacci, D.; Gupta, S.; Eder, J.P.; Golan, T.; Le, D.T.; Burtness, B.; et al. Pembrolizumab for patients with PD-L1-positive advanced gastric cancer (KEYNOTE-012): A multicentre, open-label, phase 1b trial. Lancet Oncol. 2016, 17, 717–726.
- Bang, Y.J.; Ruiz, E.Y.; Van Cutsem, E.; Lee, K.W.; Wyrwicz, L.; Schenker, M.; Alsina, M.; Ryu, M.H.; Chung, H.C.; Evesque, L.; et al. Phase III, randomised trial of avelumab versus physician’s choice of chemotherapy as third-line treatment of patients with advanced gastric or gastro-oesophageal junction cancer: Primary analysis of JAVELIN Gastric 300. Ann. Ocol. 2018, 29, 2052–2060.
- Janjigian, Y.Y.; Shitara, K.; Moehler, M.; Garrido, M.; Salman, P.; Shen, L.; Wyrwicz, L.; Yamaguchi, K.; Skoczylas, T.; Campos Bragagnoli, A.; et al. First-line nivolumab plus chemotherapy versus chemotherapy alone for advanced gastric, gastro-oesophageal junction, and oesophageal adenocarcinoma (CheckMate 649): A randomised, open-label, phase 3 trial. Lancet 2021, 398, 27–40.