1. The Infectious Cycle of S. aureus in Non-Professional Phagocytes
Intracellular survival of
S. aureus has been shown in vitro in almost every cell type, including epithelial cells, endothelial cells, osteoclasts, keratinocytes, and fibroblasts
[1][2][3][4][5][23,24,25,26,27]. However, the infectious cycle caused by the bacteria depends on the cell type that is infected (
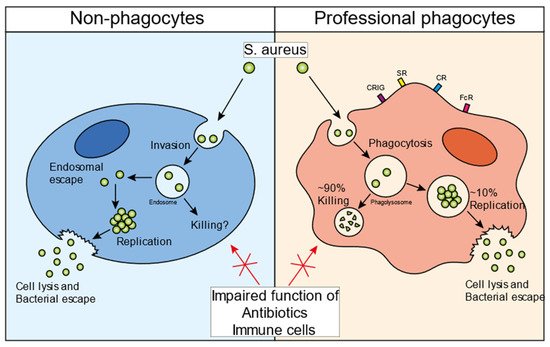
Figure 1). In general, for the invasion of non-professional phagocytes,
S. aureus employs a zipper-like mechanism that utilizes fibronectin-binding proteins A and B. These proteins attract fibronectin to the bacterial surface and this bacterial-protein complex is subsequently recognized by integrin α5β1, leading to the internalization of staphylococci in non-professional phagocytes
[6][28]. Several additional staphylococcal surface proteins have been shown to promote internalization in mammalian cells, such as clumping factor A (ClfA), iron-regulated surface determinant B (IsdB), and efflux pump tet38
[6][28].
S. aureus can escape from the endosome into the cytosol of the host cell
[7][29]. The bacteria can replicate inside the cytosol, eventually causing cell lysis and escaping into the extracellular environment (
Figure 1, left panel). Various bacterial mediators have been suggested to play a role in this escape and survival mechanism. The bacterial two-component system (TCS) accessory gene regulator (Agr), a quorum-sensing regulatory system, has been found to be a key mediator in the process of the endosomal escape of bacteria
[8][9][10][30,31,32]. In HeLa cells, Agr is required for autophagy-mediated cytotoxicity and is essential for bacterial escape into the cytoplasm, intracellular replication, and host cell killing
[10][32]. Importantly, this intracellular
S. aureus survival mechanism was not observed with an Agr mutant strain. The Agr system is a strict regulator for phenol-soluble modulin (PSM)-α, and -β production
[11][33]. Using an elegant method of
S. aureus detection in the cytosol,
thi
t iss study showed that PSM-α is crucial for the destruction of the endosomal membrane in epithelial and endothelial cells
[12][34]. An intracellular role of PSMs is supported by the fact that serum lipoproteins can protect against PSM-induced host cell lysis, both in vitro and in vivo
[13][14][35,36]. Thus, Agr and PSMs play important roles in the intracellular survival of
S. aureus in non-professional phagocytes. However, evidence for intracellular survival in these cell types has only been shown in vitro and there has rarely been any direct visualization of staphylococci persisting in non-professional phagocytes in animal models of infection or in tissues dissected from infected patients.
Figure 1. S. aureus infectious cycle. Schematic overview of the staphylococcal infectious life cycle in non-professional phagocytes (left panel) and professional phagocytes (right panel). S. aureus invades non-professional phagocytes and ends up inside the endosome. Various bacterial factor regulators, such as Agr and SigmaB, are important for the endosomal escape of the bacteria into the cytosol. Once inside the cytosol, bacteria can persist inside the host cells, maintaining an intracellular bacterial reservoir. Additionally, bacteria can start replicating, eventually lysing the host cell, and escaping into the extracellular environment. Professional phagocytes recognize bacteria with different receptors and take up bacteria by means of phagocytosis (right panel). Once inside the phagolysosome, ~90% of the bacteria are successfully killed. However, the remaining ~10% can evade the intraphagolysosomal killing strategies and then use the cells as a niche to start replicating. Similar to non-professional phagocytes, the bacteria can eventually lyse the host cell and escape into the extracellular environment. The intracellular bacteria in all cell types are protected from immune cells as well as antibiotic treatment. Abbreviations: CRIg, complement receptor of immunoglobulin superfamily; SR, scavenger receptor; CR, complement receptor; FcR, Fc receptor.
2. The Infectious Cycle of S. aureus in Professional Phagocytes
Intracellular survival has also been observed in professional phagocytes such as neutrophils and macrophages
[15][16][37,38]. There are important differences between these immune cells. Neutrophils are bone marrow-derived, short-lived, and mostly found in the bloodstream, although very limited numbers can be found extravasated in the peripheral tissues. Neutrophils are typically the first cells to arrive at the site of infection and have an incredible capacity for killing bacteria
[17][39]. Macrophages, on the other hand, populate many different tissues and body cavities in large numbers. In homeostasis, macrophages can be derived from yolk-sac or fetal liver progenitors and are self-maintained by local proliferation or by bone marrow-derived monocytes. Upon inflammation, the turnover of macrophages is primarily monocyte-dependent, as monocytes are recruited and differentiate into macrophages. Depending on the tissue they populate, macrophages have very different gene expression profiles and can exert specific cellular functions such as clearance of dead cells, antigen presentation, and coordination of responses to infection
[18][40].
In 1956, a seminal study by David Rogers showed that upon bloodstream infection in rabbits, a small number of bacteria was not removed from the blood by macrophages in the liver, but was internalized by neutrophils
[19][41]. These intracellular bacteria were predicted to be responsible for the large wave of dissemination later in infection. Since then, others have shown that live
S. aureus can survive in both mouse and human neutrophils and have proposed the “Trojan Horse” theory of dissemination by these immune cells
[20][42]. Intravenous infection of
S. aureus in mice leads to the uptake of bacteria by neutrophils that carry them in the blood for hours
[21][43]. Isolated neutrophils from an infected animal were sufficient to cause full-blown disease in healthy animals
[15][37]. Furthermore, patients with normal neutrophil counts may be more susceptible to bacterial dissemination compared to patients with reduced neutrophil counts
[22][44]. Although neutrophils have been demonstrated to enable the persistence of
S. aureus during infection, eventually leading to bacterial escape, there is limited experimental in vivo evidence that replication occurs inside these cells
[15][37].
In contrast to neutrophils, there is an increasing body of in vivo evidence that intracellular replication occurs in macrophages. During bloodstream infection, the vast majority of staphylococci are taken up in the liver by the largest population of resident macrophages in the body, called Kupffer cells (KCs). KCs are a self-sustaining population of macrophages strategically situated within the liver sinusoids and in direct contact with the blood circulation
[23][45]. In contrast to many other tissue macrophages and neutrophils, KCs are able to catch bacteria directly out of the bloodstream and overcome the high shear forces of blood flow
[24][46]. KCs are crucial cells during bloodstream
S. aureus infections, and KC depletion leads to massive, sustained bacteremia, and a greatly increased mortality due to
S. aureus infections. Since KCs can kill ~90% of the injected inoculum, they are regarded as an important initial immune bottleneck during staphylococcal bloodstream infections, whereas neutrophils are essential at later time points
[25][47]. Various studies have shown that not all staphylococci succumb to the intracellular bactericidal phagolysosome of KCs, and high-resolution intravital microscopy in combination with replication-reporter bacteria showed that staphylococci can replicate within these cells over time
[26][27][28][29][30][15,19,20,21,22].
Using intravital microscopy in living animals, it was shown that
S. aureus replicates in approximately 10% of infected KCs. Microcolonies of approximately 50–70 bacteria can be observed to lead to the destruction of the KC, which is followed by dissemination to various other internal organs, primarily the kidney (
Figure 1, right panel). Mechanistically,
S. aureus was found to replicate in phagolysosomes from KCs that did not produce sufficient reactive oxygen species (ROS) to kill the intracellular bacteria
[26][15]. Interestingly, these replicating staphylococci induced α-toxin expression, which is released back into the circulation, thereby causing platelet aggregation, microthrombus formation, vascular occlusion, and organ dysfunction
[31][48]. Even though α-toxin was upregulated by
S. aureus in KCs, it remains to be determined which staphylococcal toxin is responsible for the lysis of Kupffer cells. Jorch et al. showed that after
S. aureus escapes from the KCs, they relocate from the liver into the peritoneal cavity, where they get phagocytosed by Gata6
+ peritoneal macrophages
[32][49]. Similar to KC infections,
S. aureus can persist inside the peritoneal macrophages and disseminate to organs in the peritoneal cavity, including the kidneys. Collectively, these data show that in the murine bloodstream infection model,
S. aureus can infect and escape from multiple macrophage subtypes, ultimately seeding into the kidney, where it establishes classical abscesses 3–4 days post-infection. In a murine airway infection model, macrophages were also essential for the clearance of
S. aureus since the loss of alveolar macrophages inhibited the killing of bacteria and significantly enhanced mortality
[33][50]. Some studies have shown that
S. aureus is actively taken up by alveolar macrophages, although it is currently unclear whether
S. aureus replication occurs in this population of resident macrophages.
3. Intraphagolysosomal Evasion Strategies of S. aureus
Unlike bacterial invasion observed upon the infection of non-professional phagocytes, professional phagocytes engage in a variety of phagocytic receptors to actively engulf and internalize bacteria via phagocytosis (
Figure 1, right panel). During phagocytosis, the microbe is enclosed in a bacteria-containing phagosome within the phagocytic cell. These phagosomes undergo several maturation steps to ultimately become phagolysosomes, a process in which the interior of the vesicle progressively increases its acidity. Phagolysosomes contain a variety of antimicrobial peptides and degradative lysosomal enzymes, and are bombarded with ROS, all of which contribute to the killing of ingested bacteria
[34][51]. The microbicidal activity of the phagolysosome is incredibly disruptive to most bacteria, and in order to establish an infection, many pathogens have evolved various strategies to neutralize or resist the effector components of the phagolysosome. Nonetheless, for
S. aureus, there is some controversy on how these bacteria can overcome the phagosomal effector functions. Some literature suggests that the phagosomal maturation pathway is hijacked by
S. aureus [30][35][22,52], while most papers found normal phagosomal maturation suggesting that
S. aureus can withstand the harmful environment of the phagolysosome
[12][26][29][36][37][38][39][3,15,21,34,53,54,55].
S. aureus, therefore, must overcome these defensive mechanisms in order to enable intracellular replication. Unsurprisingly, the bacteria express a plethora of evasion molecules inside the phagolysosome (
Figure 2).
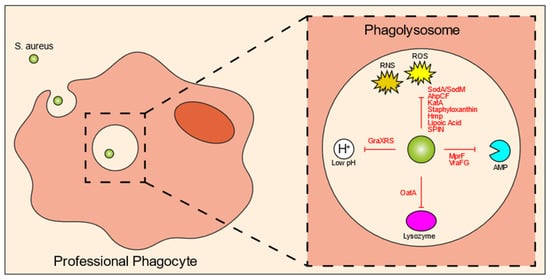
Figure 2. S. aureus intraphagolysosomal evasion strategies. Once inside the phagolysosome of professional phagocytes, the bacteria get exposed to various antimicrobial molecules designed to kill the bacteria. Co-evolution of host and pathogen has led to a variety of evasion molecules produced by S. aureus to counteract these phagolysosomal killing mechanisms. These mechanisms include evasion molecules against ROS and RNS, AMPs, lysozyme, and acidic pH. Abbreviations: ROS, reactive oxygen species; RNS, reactive nitrogen species; Sod, superoxide dismutase; AhpCF, alkyl hydroperoxidase reductase; KatA, catalase; Hmp, flavohemoglobin; SPIN, staphylococcal peroxidase inhibitor; AMP, antimicrobial peptide; MprF, multiple peptide resistance factor; OatA, O-acetyltransferase.
One of the most important killing mechanisms of the host cells is ROS, which is produced by NADPH oxidase (NOX2) via catalyzation of superoxide (O
2−). Oxygen radicals have great antimicrobial activity, targeting various bacterial components, such as DNA, proteins, and the bacterial cell membrane. Nitrogen radicals can also be created by immune cells, termed reactive nitrogen species (RNS). It is not surprising that most
S. aureus evasion molecules are directed against these molecules (
Figure 2). SodA and SodM are staphylococcal superoxide dismutases that directly eliminate ROS
[40][41][57,58]. Similarly, alkyl hydroperoxidase reductase (AhpCF) and catalase (KatA) directly incapacitate peroxides and H
2O
2, respectively
[42][43][59,60]. Staphyloxanthin is a carotenoid pigment with strong antioxidant properties
[44][61]. For nitric oxide resistance,
S. aureus can produce flavohemoglobin (Hmp), which acts like a denitrosylase
[45][62]. ROS and RNS production can be inhibited by lipoic acid
[46][63], and staphylococcal peroxidase inhibitor (SPIN) can bind to and block myeloperoxidase (MPO), which is the enzyme that converts H
2O
2 and chloride into hypochlorous acid
[47][64].
A recent study by Leliefeld et al. using an in vitro fibrinogen gel model to increase the lifespan of neutrophils, showed that neutrophils limit staphylococcal growth in a pH-dependent manner
[48][65]. Contrarily, in macrophages, it was shown that the intraphagolysosomal acidic pH is important for
S. aureus to enable replication. This was orchestrated by GraXRS, which has been shown both in vitro as well as in vivo
[49][50][66,67]. Similar to survival inside non-professional phagocytes, SigmaB could play a role in intraphagosomal survival, because a wild-type
S. aureus strain showed increased resistance to acidic pH and hydrogen peroxide, when compared to a SigmaB-inactivated mutant in vitro
[51][68]. In line with this observation, the SigmaB activating protein RsbU has been shown to be necessary for the intracellular growth of
S. aureus within the phagolysosomes of THP-1 macrophages and HUVEC cells
[52][69]. Various host cell-derived enzymes are released into the phagolysosome, among which are proteases. Lysozyme, which can cleave bacterial peptidoglycan and subsequently induce bacterial cell lysis, is particularly harmful. In response,
S. aureus produces O-acetyltransferase (OatA) that acetylates peptidoglycan so it can no longer be targeted by lysozyme
[53][54][70,71]. Finally, there are other antimicrobial peptides released into the phagolysosomal environment, many of which are positively charged. Multiple peptide resistance factors (MprF) of
S. aureus catalyze a reaction that attaches a positively-charged lysine to the negatively-charged lipids of the bacterial membrane, thereby reducing interaction with antimicrobial peptides, defensins and protegrins in particular
[55][56][57][72,73,74]. VraFG is a transporter of antimicrobial peptides and helps with resistance against cationic antimicrobial peptides
[58][75].
Not all studies suggest that
S. aureus replicates within the phagolysosome. Kubica et al. reported that
S. aureus escaped into the cytosol of macrophages using α-toxin, where they replicated for a few days followed by host cell lysis. Several
S. aureus factors were seen to be important for this survival, including SigmaB and Agr
[59][76]. Another study suggested that the
S. aureus USA300 strain can disturb normal phagolysosome formation and can sense the intraphagolysosomal pH in order to escape into the cytosol and commence intracellular replication
[29][21]. Staphylococcus-produced PSMs have been shown to be important for killing neutrophils after phagocytosis. These toxins are actively upregulated by the bacteria within the phagolysosome by Agr and the stringent response system, and both systems appear to be crucial for the intracellular production of PSMs and escape from neutrophils
[60][61][77,78].
43. Bacterial Specificity of the Intracellular Reservoir
Collectively, research has shown that
S. aureus can persist and replicate in multiple cell types, maintaining the bacterial population during infection, and leading to the dissemination of bacteria throughout the body. Commensal bacteria are typically cleared by host phagocytes after infection, whereas non-aureus staphylococci can persist in macrophages but do not replicate
[62][79]. Co-infection of
S. aureus and commensal bacteria only led to the proliferation of
S. aureus within KCs
[63][80]. The commensal bacteria augment
S. aureus infection by acting as a sink for ROS production, thereby increasing the chance that
S. aureus can initiate replication inside KCs
[64][81]. Many other bacterial, fungal, or parasitic species have been shown to use circulating phagocytes as an opportunity for dissemination, such as
Mycobacterium tuberculosis,
Listeria monocytogenes,
Leishmania, and
Cryptococcus neoformans [65][66][67][68][82,83,84,85].
The ability to persist and replicate intracellularly might be
S. aureus strain specific. For example, where the Newman strain has been shown to be acid sensitive and is cleared in an intracellular infection
[69][86], the more virulent USA300 strain can survive in phagocytes and has even been suggested to require acidification of the phagolysosome for persistence
[50][67]. In a study where different
S. aureus strains (Cowan I, 6850, Novel, Wood 46, BS 507, BS 513, BS 800, BS 890, BS 891, and RN4220) were tested in vitro with different cell lines (HEK-293, HeLa, and EA.hy926 cells), it was observed that all strains survived inside fully acidified vesicles except for
S. aureus 6850, which was present in vesicles with reduced acidification
[39][55]. Xiong et al. compared genotypes of persistent
S. aureus strains with strains associated with rapid clearance in the blood and showed that persistence was associated with overexpression of Agr, staphylococcal cassette chromosome mec type II, clonal complex 30, and spa 16
[70][87]. Hence, only some
S. aureus strains have the unique ability to persist and grow inside host phagocytes. As outlined above, various virulence mechanisms have implicated the ability of
S. aureus to withstand the innate defenses of neutrophils and macrophages, even though not all studies are in agreement. The discrepancy between studies could be due to staphylococcal strain variation, use of distinct cells and or cell lines, and methodological differences. It should also be noted that in vitro generated data might be far from the actual in vivo survival mechanisms of
S. aureus within macrophages. Nevertheless, it is safe to assume that there is a redundancy in evasion strategies that provides the bacteria time to adapt to the intraphagolysosomal environment to facilitate the intracellular reservoir.