Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Dipendra Mahato and Version 3 by Theodoros Varzakas.
Mycotoxins are secondary metabolites produced by fungi that infect a wide range of foods worldwide. Nivalenol (NIV), a type B trichothecene produced by numerous Fusarium species, has the ability to infect a variety of foods both in the field and during post-harvest handling and management. NIV is frequently found in cereal and cereal-based goods, and its strong cytotoxicity poses major concerns for both human and animal health.
- nivalenol
- food contamination
- human and animal health
1. Introduction
Mycotoxins, which are secondary metabolites produced by many fungi, can be detrimental to both human and animal’s health when ingested through contaminated food [1][2][3][4][5][6][1,2,3,4,5,6]. The contamination of crops and their processed products is a major public health and economic concern. More than 80% of the agricultural produce is contaminated by at least one mycotoxin. Several genera of fungi, including Cephalosporium, Cyclindrocarpon, Myrothecium, Phomopsis, Stachybotrys, Trichoderma, Trichotecium, and Verticimonosporium species, are mainly responsible for the mycotoxin production among which the trichothecene group of mycotoxins share a major proportion, and hence pose wider concerns [7]. The trichothecene group of mycotoxins are categorized as type A, B, C, and D, which includes more than 170 different toxins, but their toxicity potential depends upon the type of mycotoxin being produced by the fungi [8].
Nivalenol (NIV) is one of the major mycotoxins, produced by several Fusarium species and belongs to the type B trichothecene. It is a natural contaminant found worldwide in various foodstuffs. Due to the potent cytotoxicity, NIV constitutes a serious health risk to both humans and animals. Its contamination is observed very frequently in agricultural products. Crops, such as wheat, barley, and corn, are usually contaminated with NIV, as these crops are more prone to the growth of NIV-producing fungal species [9][10][11][9,10,11]. Furthermore, NIV is also resistant against food processing conditions, thereby leading to the further spread of NIV-induced mycotoxicosis in consumers [12]. Controlling NIV exposure is especially important because of how frequently it contaminates foods and how easily it can harm mammals through immunotoxicity and hemotoxicity [13]. NIV can produce certain complications, such as inhibition of cell proliferation, induction of CXCL8/interleukin (IL) -8 secretions, and the involvement of stress-activated mitogen-activated protein kinases [14][15][16][14,15,16].
Chemically, the structure of NIV is very similar to DON (deoxynivalenol) mycotoxin of the type B trichothecene group. The only difference between NIV and DON is one oxygen atom at the C-4 position (Figure 1); however, the toxicity of these oxides of carbon completely differs from each other. The co-occurrence of NIV and DON has been reported extensively, but the oxidative stress and toxicity induced by NIV contamination are higher than that described for DON [7][12][7,12]. Several studies have reported the contamination of NIV in foods, including wheat [17][18][17,18], barley [19][20][19,20], groundnuts [21], maize and their by-products [22][23][24][22,23,24], but the level of contamination varies with the type of crop and ecological factors, such as temperature and humidity. Magan et al. [25] reported that environmental factors, including farm practices, handling, storage, processing, and distribution of food grains, largely influence the spread of NIV contamination. Therefore, the knowledge of climatic conditions on the growth of Fusarium species, along with its behavior and mycotoxin production, is essential to control the risk of NIV contamination in crops [26].
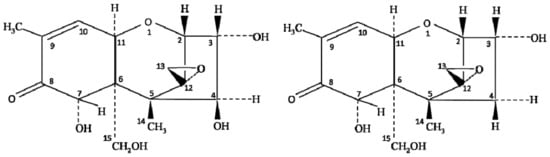
Figure 1.
Chemical structure of nivalenol (NIV) and deoxynivalenol (DON).
2. Chemistry and Biosynthesis of Nivalenol
Trichothecenes are sesquiterpenoids with a variable pattern of oxygen and ester groups on a core tricyclic skeleton containing an epoxide function, also known as the 12,13-epoxytrichothec-9-ene (EPT) [27]. A large variety of trichothecene have been found in diverse fungal taxa and are grouped into four classes (types A-D) based on the substitution pattern on the fundamental EPT core [28][29][28,29]. Numerous research has been conducted to understand the various impacts of trichothecenes and to study the underlying mechanisms to mitigate the huge losses caused by Fusarium diseases globally [30][31][32][33][30,31,32,33], which endanger the viability of farms and entire rural communities.
Fusarium’s two primary chemotypes are connected to organisms that either generate deoxynivalenol or nivalenol [34]. The functioning of two genes, TRI13 that expresses a trichothecene 4-hydroxylase and TRI7 that encodes a trichothecene 4-acetyltransferase, has been shown to be the genetic difference between deoxynivalenol and nivalenol chemotypes. Both genes are present but inactive in Fusarium chemotypes that produce deoxynivalenol [35][36][35,36]. Compared to its type B companion, DON and NIV, which are more abundant in Asia, contaminate grains at greater levels and cause more severe acute and chronic toxicities. In F. graminearum, twelve trichothecene genes are grouped within a 25-kb area on chromosome II that is inherited as a single core TRI cluster [37][38][37,38]. Two more loci are involved, including the single gene Tri101 and the two-gene Tri1-Tri16 locus, both of which are situated in unrelated chromosomal areas [39]. In F. graminearum, the presence of both functioning Tri7 and Tri13 genes is needed for NIV-producing chemotypes [35][40][35,40].
Biosynthetic clusters are used in fungi to produce secondary or specialized metabolites [41]. As a result, the proteins that catalyze the biosynthesis of distinct trichothecenes are likewise arranged in biosynthetic clusters (TRI) [37][38][37,38], where many of the genes appear to be coordinately expressed and controlled. Intermediates from two separate trichothecene routes, deoxynivalenol biosynthesis and T-2 toxin generation, can feed the nivalenol process. The entry processes from the deoxynivalenol biosynthesis into the nivalenol biosynthesis are mediated by TRI13, which codes for a trichothecene 4-hydroxylase, and TRI7, which codes for a trichothecene 4-acetyltransferase and is non-functional in Fusarium species of the deoxynivalenol chemotype.
The gene TRI13 encodes a trichothecene 4-hydroxylase that has been identified and described from Fusarium graminearum strain 88-1, which represents the nivalenol chemotype. This enzyme catalyzes the conversion of 3,15-acetyldeoxynivalenol to 3,15-diacetylnivalenol, hence functioning as a switch between deoxynivalenol-producing and nivalenol-producing Fusarium graminearum strains [35][42][35,42]. The TRI7-encoded trichothecene C-4-acetyltransferase then performs C-4 acetylation, leading to the formation of 3,4,15-triacetylnivalenol [35][42][43][35,42,43]. The same intermediate can also be produced via the trichothecene 7,8-dihydroxylase (TRI1), which acts as 3,4,15-triacetoxyscirpenol 7,8-dihydroxylase and converts 3,4,15-triacetoxyscirpenol, an intermediate of the T-2 toxin biosynthesis, into 7,8-dihydroxy-3,4,15-triacetoxyscirpenol, followed by a currently unknown protein that confers the carbonyl function to the C-8 of 3,4,15-triacetylnivalenol [44][45][44,45]. In Fusarium graminearum strain 79A1 [46], the trichothecene C-3 esterase (TRI8) deacylates 3,4,15-triacetylnivalenol, generating 4,15-diacetylnivalenol, which is subsequently deacylated at C-4 by an unknown protein. Furthermore, in the NIV chemotype, Tri13 hydroxylates 3,15- diacetyldeoxynivalenol to yield 3,15-diacetylnivalenol, which is further transformed into NIV [47].
3. Occurrence of Nivalenol in Food and Feed
Several species of Fusarium, including F. poae, F. culmorum, F. grainearum, F. cortaderae, F. asiaticum, F. nivale, F crookwellense etc., are mainly responsible for producing NIV and its acetyle derivatives in foods [18][24][48][49][18,24,55,56]. The Fusarium species easily grow on cereal grain and produce Fusarium head blight disease under favorable environmental conditions. In general, moderate temperature and high air humidity are ideal for the infection and growth of these Fusarium species. However, sometimes it may vary from species to species, such as F. graminearum are usually found in the warmer and more humid regions (Australia, Eastern Europe, North America, South China), whereas cold climatic regions (Western Europe) are suitable for the growth of F. culmorum species [50][57]. NIV is reported in various cereals and cereal-based products worldwide (Table 1). It is also interesting to note that NIV mostly co-occurs with DON and is detected simultaneously in different food matrices. Furthermore, most of the methods mentioned in Table 1 were used for the multi-mycotoxin analysis.Table 1. Occurrence of nivalenol in food and feed around the world.
| Food Matrix | Country | Range (μg/kg) | Detection Technique | References |
|---|---|---|---|---|
| Food | ||||
| Adlay millet | South Korea | 12.6–337.6 | HPLC-UV | [51][58] |
| Baby foods | Spain | 75–100 | HPLC-MS/MS | [52][59] |
| Baby formula | South Korea | 4.4–1000 | HPLC-UV | [53][60] |
| Barley | England | 10–1088 | GC/MS | [54][61] |
| South Korea | 10.4–110.3 | HPLC-UV | [51][58] | |
| Italy | 21.7–106 | LC-MS/MS | [55][62] | |
| Spain | 12.47 | GC-MS | [56][63] | |
| Baked snacks | Spain | 55.7 | GC-MS | [57][64] |
| Barley grain | Poland | 5 | TLC and HPLC | [58][65] |
| Barley and barley products | Germany | 0.87–19 | LC-MS/MS | [59][66] |
| Beer | Czech Republic | 4–6 | UHPLC-APCI-Orbitrap MS | [60][67] |
| Beer | Spain | 10–15 | UHPLC-APCI-Orbitrap MS | [61][68] |
| Black bean paste (Chunjang) | South Korea | 83.8 | HPLC-UV | [51][58] |
| Breakfast cereals | Spain | 51.1–106.5 | GC-MS | [57][64] |
| Breakfast cereals | South Korea | 1096.8 | GC-MS | [62][69] |
| Brown rice | South Korea | 47.4 | HPLC-UV | [51][58] |
| Cereals | Finland | 185–300 | LC-MS/MS | [63][70] |
| Cereals | Czech Republic | 50 | UHPLC-ESI-ToF-MS | [64][71] |
| Cereal based products | Czech Republic | 25 | UHPLC-APCI-MS/MS | [65][72] |
| Cereal based products | Switzerland | 100 | HPLC-ESI-MS/MS | [66][73] |
| Cereals and cereal-based products | Spain | 121–176 | LC-MS/MS | [67][74] |
| Corn | South Korea | 0–181.41 * | HPLC-PAD | [68][75] |
| Corn | France | 7–340 | HPLC | [69][76] |
| De-hulled and naked barley | Spain | 1.1–7.6 | LC-MS/MS | [59][66] |
| Durum wheat | France | 60 | HPLC | [69][76] |
| Durum wheat flour | Denmark | 83–440 | GC-ECD | [70][77] |
| Flour bread | Italy | 5–8 | LC-MS/MS | [55][62] |
| Foxtail millet | South Korea | 27.4–370.8 | HPLC-UV | [51][58] |
| Ground wheat | Italy | 3.5–63.5 | LC-APCI-MS/MS | [71][78] |
| Groundnut-maize based snacks | Nigeria | 1.8–2.5 | LC-MS/MS | [72][79] |
| Grain-based product | Italy | 30 | GC-MS | [73][80] |
| Groundnut | Nigeria | 1.0 | LC-MS/MS | [72][79] |
| Malting barley | Spain | 35 | LC-MS/MS | [59][66] |
| Maize | South Korea | 51.3 | HPLC-UV | [51][58] |
| Nigeria | 0.8 | LC-MS/MS | [72][79] | |
| China | 2.1–15.3 | UHPLC-MS/MS | [74][81] | |
| Austria | 22.3–250 | LC-MS/MS | [75][82] | |
| Spain | 6.4 | GC-MS/MS | [76][83] | |
| Germany | 4.41–20 | GC | [77][84] | |
| UK | 5–10 | HPLC | [78][85] | |
| Poland | 2 | TLC and HPLC | [58][65] | |
| Maize flour | Germany | 39 | GC-MS | [79][86] |
| Maize-based breakfast cereal | Spain | 16–60.2 | GC-MS | [57][64] |
| Multicereal Flour | Spain | 75 | LC-MS/MS | [80][87] |
| Mixed paste | South Korea | 15.9–100.6 | HPLC-UV | [51][58] |
| Mixed grains | South Korea | 88.9 | GC-MS | [62][69] |
| Oats | South Korea | 23.5 | HPLC-UV | [51][58] |
| Italy | 45.5–50.4 | LC-MS/MS | [55][62] | |
| Germany | 17 | LC-MS/MS | [81][88] | |
| Italy | 8–20 | LC-MS/MS | [82][89] | |
| Austria | 100 | HPLC | [69][76] | |
| England | 10–112 | GC/MS | [83][90] | |
| Oats grain | Poland | 6 | TLC and HPLC | [58][65] |
| Pearl barley | Spain | 0.18 | LC-MS/MS | [59][66] |
| Popcorn | South Korea | 68.7 | HPLC-UV | [51][58] |
| Red chili paste (Gochujang) | South Korea | 8.5–120.2 | HPLC-UV | [51][58] |
| Rice | South Korea | 10 | HPLC | [84][91] |
| Rice | Thailand | 0.50–15.00 | UHPLC-MS/MS | [85][92] |
| Rice wine | South Korea | 2.5 | HPLC-UV | [53][60] |
| Rye | Italy | 33.9–34.4 | LC-MS/MS | [55][62] |
| Germany | 1.8 | LC-MS/MS | [81][88] | |
| France | 2–48 | HPLC | [69][76] | |
| Rye flour | Denmark | 38–48 | GC-ECD | [70][77] |
| Rye grain | Poland | 5 | TLC and HPLC | [58][65] |
| Sesame butter | China | 0.05–7.25 | UHPLC-MS/MS | [86][93] |
| Semolina | Germany | 36 | GC-MS | [79][86] |
| Wheat | Italy | 12–106 | LC-MS/MS | [55][62] |
| Japan | 0.2 | HPLC-AAPI-MS/MS | [87][94] | |
| Spain | 53.6 | GC-MS/MS | [76][83] | |
| Germany | 33 | GC-MS | [88][95] | |
| Poland | 10 | GC-GC-ToF-MS | [89][96] | |
| Argentina | 0.11–0.40 | HPLC | [90][97] | |
| England | 10–330 | GC/MS | [91][98] | |
| Wheat flour | South Korea | 31.8 | GC-MS | [62][69] |
| Denmark | 10–189 | GC-ECD | [70][77] | |
| Spain | 30 | HPLC-ESI-MS/MS | [92][99] | |
| Wheat semolina | Spain | 8.8–13.6 | GC-MS/MS | [93][100] |
| Winter barley | Spain | 5.6–6.5 | LC-MS/MS | [59][66] |
| Winter wheat | Italy | 70 | HPLC-MS/MS | [94][101] |
| Spelt | Italy | 23 | LC-MS/MS | [55][62] |
| Feed | ||||
| Bran | South Korea | 11.1–36.9 | HPLC-UV | [84][91] |
| Cattle feed | South Korea | 0–111.52 * | HPLC-PAD | [68][75] |
| Chicken feed | South Korea | 0–101.23 * | HPLC-PAD | [68][75] |
| Maize silages | Denmark | 122 | LC-MS/MS | [95][102] |
| Pig feed | South Korea | 0–84.21 * | HPLC-PAD | [68][75] |
| Wheat germ | Germany | 26 | GC-MS | [79][86] |
| Wheat bran | Germany | 37 | GC-MS | [79][86] |
* ng/kg of samples. GC: gas chromatography; GC-MS: gas chromatography–mass spectrometry; GC-ECD: gas chromatography with electron capture detection; GC-GC-ToF-MS: two-dimensional gas chromatography/time-of-flight-mass spectrometry; GC-MS/MS: gas chromatography coupled with tandem mass spectrometry; HPLC-AAPI-MS/MS: high performance liquid chromatography/atmospheric pressure ionization/tandem mass spectrometry; HPLC-APCI-MS/MS: high-performance liquid chromatography–atmospheric pressure chemical ionization–tandem mass spectrometry; HPLC-ESI-MS/MS: high-performance liquid chromatography/electrospray ionization tandem mass spectrometry; HPLC-MS/MS: high performance liquid chromatography and tandem mass spectrometry; HPLC-PAD: high-performance liquid chromatography-photodiode array detector; HPLC-UV: high performance liquid chromatographic method coupled with ultraviolet detector; LC-APCI-MS/MS: liquid chromatography coupled with atmospheric pressure chemical ionization triple quadrupole mass spectrometry; LC-MS/MS: liquid chromatography–tandem mass spectrometry; TLC: thin layer chromatography; HPLC: high performance liquid chromatography; UFLC-MS/MS: ultra-fast liquid chromatography tandem mass spectrometry; UHPLC-APCI-MS/MS: ultra-performance liquid chromatography–atmospheric pressure chemical ionization tandem mass spectrometry; UHPLC-APCI-Orbitrap MS: ultrahigh-performance liquid chromatography electrospray ionization quadrupole Orbitrap high-resolution mass spectrometry; UHPLC-ESI-ToF-MS: ultra-high-performance liquid chromatography coupled with electrospray ionization quadrupole time-of-flight mass spectrometry; UHPLC-MS/MS: ultra-high performance liquid chromatography tandem mass spectrometry; UHPLC-QqLIT-MS: ultra-high-performance liquid chromatography quadrupole linear ion trap mass spectrometry.