In traditional medicine, different parts of plants, including fruits, have been used for their anti-inflammatory and anti-oxidative properties. Plant-based foods, such as fruits, seeds and vegetables, are used for therapeutic purposes due to the presence of flavonoid compounds. Proanthocyanidins (PCs) and anthocyanins (ACNs) are the major distributed flavonoid pigments in plants, which have therapeutic potential against certain chronic diseases. PCs and ACNs derived from plant-based foods and/or medicinal plants at different nontoxic concentrations have shown anti-non-small cell lung cancer (NSCLC) activity in vitro/in vivo models through inhibiting proliferation, invasion/migration, metastasis and angiogenesis and by activating apoptosis/autophagy-related mechanisms.
- flavonoids
- proanthocyanidins
- anthocyanian
- NSCLC
1. Introduction
2. Proanthocyanidins in Nicotine-Induced NSCLC Treatment
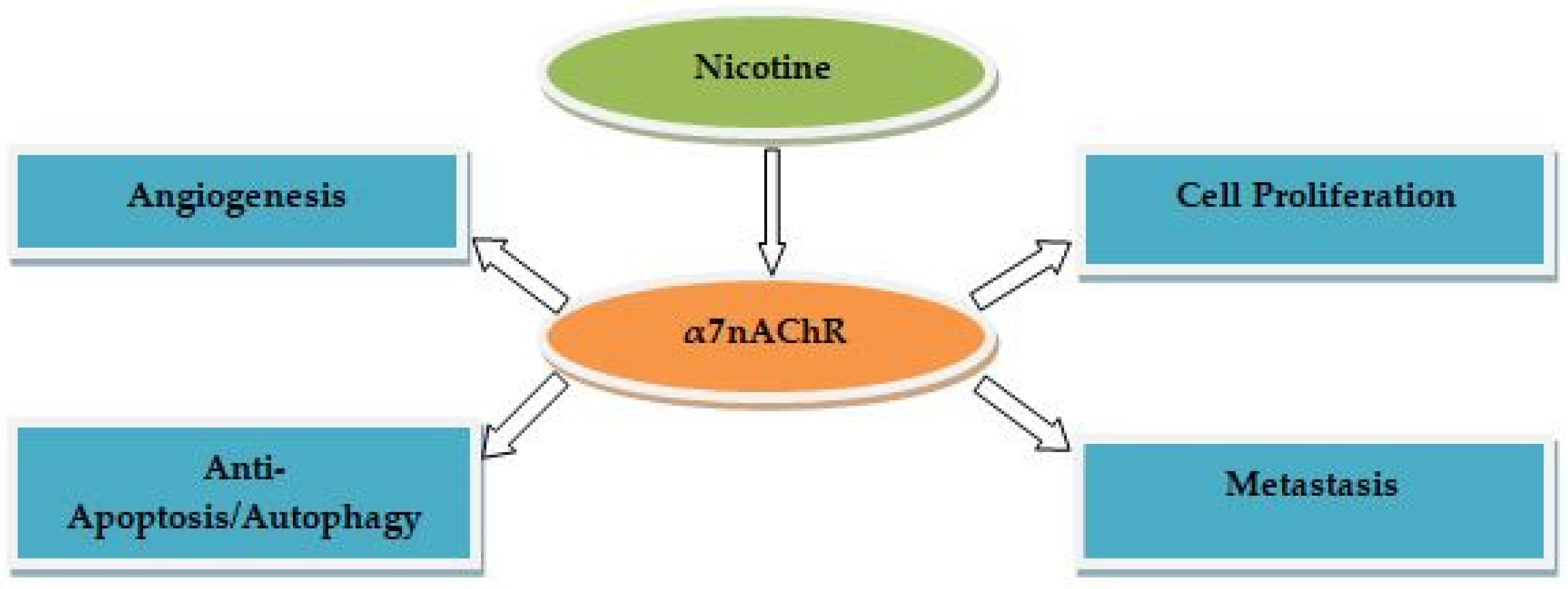
Several studies demonstrated the therapeutic effects of PC-rich extracts from plant-based foods and/or medicinal plants at different nontoxic concentration against nicotine-induced NSCLC. Cranberry-derived PCs suppress tumor cell growth in NSCLC cells [40[26][27],41], but the mechanisms for this action have not been well investigated [40][26]. Treatment with cranberry PCs resulted in a significant induction of apoptosis and cell cycle arrest in NSCLC cells via upregulating the expression of pro-apoptotic-related markers (e.g., cytochrome c and caspase 3) [42][28]. PCs from grape seed extract have shown promising results in nicotine-induced NSCLC treatment. For example, using the in vivo proteolysis/antitumor assay and the in vitro proteolysis/angiogenesis assay, PCs inhibit angiogenesis-mediated tumor growth in NSCLC cells, in part by suppressing vascular extracellular matrix (ECM) proteolysis byMMP-2 [43][29]. Treatment of NSCLC cells with PCs using the in vivo tumor xenograft assay and the in vitro 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay for cell proliferation/survival resulted in suppression of cell proliferation in vitro/vivo, and inhibition of angiogenesis and induction the apoptotic cell death of tumor cells in vivo. Such effects are mediated by upregulation of insulin-like growth factor binding protein-3 (IGFBP-3) levels and inhibition of proliferating cell nuclear antigen (PCNA) in the tumor microenvironment [44][30]. A study used the in vivo tumor xenograft assay and the in vitro cell deathenzyme-linked immunosorbent assay (ELISA) and MTT assay for assessing proliferation of NSCLC cells showed that PCs cause proliferation inhibition and apoptosis induction via the inhibition of cyclooxygenase-2 (COX-2) expression and prostaglandin-2 (PGI-2) receptors in NSCLC cells [45][31]. The mechanism underlying the anti-migration effect of PCs on NSCLC cells involve inhibiting nitric oxide (NO) synthase, N(G)-nitro-L-arginine methyl ester (L-NAME), and the ERK1/2 and MAPK signaling pathways [46][32]. The anti-proliferative/apoptotic effects of PCs onNSCLC cells are mediated via the activation of caspase 3 expression, prostacyclin synthase (PTGIS)/PGI2 (as measured by 6-keto PGF1α), and 15-lipoxigenase-2/15(S)-hydroxyeicosatetraenoic acid (15-LOX-2)/15-HETE production [47][33]. PCs showed the inhibitory effects on the cigarette smoke condensate (CSC)-induced migration of NSCLC cells through inhibition of NADPH oxidase (NOX)-induced oxidative stress and EMT transition [48][34]. Treatment with PCs using the colorimetric caspase-3 activity assay in vivo and in vitro showed apoptotic effects through increased expression of pro-apoptotic markers (e.g., poly ADP ribose polymerase (PARP); Bcl-2-associated X protein (Bax)), and decreased expression of apoptotic markers (e.g., Bcl-2 and cyclins) [49][35]. A study used the in vivo tumor xenograft assay and the in vitro MTT and miR-106b ISH assays showed that PCs promote anti-proliferative/invasive effects on NSCLC cells via downregulating miR-106b expression and upregulating cyclin-dependent kinase inhibitor 1A (CDKN1A) mRNA and p21 expression [50][36]. A few studies on NSCLC cells after treatment with the Cinnamomi Cortex extract PCs showed a significant reduction in nuclear factor-E2-related factor 2(Nrf2) expression, and insulin-like growth factor-1 receptors (IGF-1R) were responsible for induced proliferation [51,52][37][38]. Cinnamomi Cortex extract procyanidin C1 exert anti-metastatic activity by suppressing TGF-β-induced EMT in NSCLC cells [53][39]. Treatment with PCs inhibits hydrogen peroxide (H2O2)-induced NSCLC cell viability, as shown by reduced reactive oxygen species (ROS) and malondialdehyde (MDA) production, hydrogen peroxide(H2O2)-induced oxidative stress, and promoted the expression of Nrf2 target genes [54][40]. PCs inhibit proliferation, viability, along with induction of apoptosis and G2/M cell cycle arrest in NSCLC cells. This is triggered by inhibiting the EMT-related molecules (e.g., N-cadherin and vimentin), expression of apoptotic markers (e.g., Bcl-2), and increasing expression of pro-apoptotic markers (e.g., Bax) via downregulating the Janus kinase/signal transducer and activator of transcription3 (JAK2/STAT3) signaling pathway [55][41]. Treatment with prodelphinidin B-2 3′-O-gallate, a proanthocyanidin gallate, resulted in the upregulation of key transcription factors such as the soluble Fas ligand (sFasL) and membrane-bound Fas ligand (mFasL), which are responsible for the anti-proliferative and apoptotic activities in NSCLC cells [56,57][42][43]. Cinnamtannin D1, an A-type procyanidin trimer, from Rhododendron formosanum extracts has been found to exhibit autophagic effects on NSCLC cells via inhibition of cellular signaling pathways (e.g., mTOR) [58][44]. These results suggest that plant-derived natural PCs may play a significant role as anti-NSCLC agents by suppressing proliferation, migration, invasion, viability, metastasis, angiogenesis, and promoting apoptosis/autophagy via inhibition/activation of transcription factors and/or multiple cellular signaling pathways induced by α7nAChR in NSCLC cells. Table 1 highlights the molecular mechanisms of PCs in nicotine-induced NSCLC treatment.| Study Type | NSCLC Cell Type | Extract/Compound | Concentrations | Activity | Mechanisms of Action | Reference | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| In vitro | DMS114 | Cranberry presscake | 200–300 µmol/L | Anti-proliferative, apoptosis | NA | [40] | [26] | |||||||||
| In vitro | H460 | Cranberry ( | Vaccinium macrocarpon | ) | 20–80 µg/mL | Anti-proliferative | MMP2, MMP9↓ | [41] | [27] | |||||||
| [ | 61 | ] | [ | 47 | ] | In vitro | ||||||||||
| In vitro | H460 | Cranberry ( | Vaccinium macrocarpon | ) | 50 µg/mL | Apoptosis, cell cycle arrest | P21, P73, PARP, cytochrome | c | , caspase3/4/8↑ Bcl-2↓ |
[42] | [28] | |||||
| A549, H1299 | Cyanidin-3-glucoside | 5, 10, 20, 40, and 80 µM | Anti-proliferative, anti-migration, anti-invasive, apoptosis | TP53I3 andPI3K/Akt/mTOR↓ | [ | 62] | [48] | In vitro/vivo | A549 | Grape seed | 100 µg/mL (in vitro) 30 mg PC/kg bodyweight (in vivo) |
Inhibition of tumor angiogenesis | ||||
| In vitro/vivo | A549, H441, SK-MES-1 | Delphinidin | 5–100 µM (in vitro) 1, 2 mg PC/kg bodyweight (in vivo) |
Inhibition of tumor growth, anti-proliferative, anti-angiogenic, apoptosis | MMP2↓ | EGFR, Bcl-2, PCNA, cyclin D1, VEGFA, Akt/PI3K/MAPK↓ Bax, caspase-3/9↑ | [43] | [29] | ||||||||
| [ | 63 | ] | [ | 49 | ] | In vitro/vivo | A549, H1299 | Grape seed | 60, 80 µg/mL (in vitro) Administration of PCs (0.1%, 0.2%, and 0.5%, bodyweight) (in vivo) |
Anti-proliferative, anti-angiogenic, apoptosis | PCNA↓ IGFBP-3↑ |
|||||
| [ | ||||||||||||||||
| 36 | ||||||||||||||||
| In vitro/vivo | A549 | Delphinidin | 10, 20, and 40 µM (in vitro) 80 µM (in vivo) | [ | 44 | Anti-angiogenic | ] | [ | EGF, CoCl2, HIF-1α, ERK, VEGF mRNA, Akt/mTOR/PI3K/p70S6K↓30] | |||||||
| [ | 64 | ] | [ | 50 | ] | In vitro/vivo | H157, H226, H460, H1299, A549 | Grape seed | 20, 40, and 60 µg/mL (in vitro) Administration of PCs (0.5%, bodyweight) (in vivo) |
Anti-proliferative, apoptosis | COX-2, PGI-2↓ | [45] | [31] | |||
| In vitro | A549, H1299 | Grape seed | 10, 20, 40, and 60 µg/mL | |||||||||||||
| In vitro/vivo | A549, H1299 | Bilberry and blueberry | 3.125–12.5 µM (in vitro) | Anti-migration | NO, L-NAME, MAPK, ERK1/2↓ | [46] | [32] | |||||||||
| ] | ||||||||||||||||
| In vitro | A549 | Cinnamomi Cortex | 2.5 µg/mL | Inhibition of cell viability and proliferation | Nrf2↓ | [51] | [37] | |||||||||
| 1.5 mg PC/kg bodyweight (in vivo) | Anti-metastatic, anti-invasive, apoptosis, cell cyclearrest | TNFα-induced NF- | k | B, Notch, Wnt/β-catenin, cyclinD1/B1, VEGF, p-ERK, bcl-2, MMP9 ↓ | [ | 65] | [51] | |||||||||
| In vitro | A549 | Syzygium cumini | L. | 2.5, 5, 10, 20, and 25 µM | Anti-proliferative | NA | [66] | [ | In vitro | A549 | Grape seed | 6 µg/mL | Anti-proliferative, apoptosis | caspase 3, PTGIS/PGI2↑ | [47] | [33] |
| 52 | ] | In vitro | A549, H1299, H460 | Grape seed | 20 and 40 µg/mL | Anti-migration | E-cadherin, NOX, p22/p47(phox)↓ N-cadherin, fibronectin, vimentin↑ |
[48] | [34] | |||||||
| In vitro/vivo | A549, H1299 | Grape seed | 20, 40, and 60 µg/mL (in vitro) 50, 100, and 200 mg PC/kg bodyweight (in vivo) |
Apoptosis | G1arrest, Bax, caspases-3/9, Cdki, PARP↑ Bcl-2,Bcl-xl, Cdk2/4/6, cyclins↓ |
[49] | [35] | |||||||||
| In vitro/vivo | A549 | Grape seed | 45 µg/mL (in vitro) 112 mg PC/kg bodyweight (in vivo) |
Anti-proliferative, anti-invasive | CDKN1A, p21↑, miR-106b↓ |
[50] | In vitro | A549 | Cinnamomi Cortex | 10 µg/mL | Inhibition of cell proliferation | Nrf2, IGF-1R↓ | [52] | [38] | ||
| In vitro | A549 | Cinnamomi Cortex | 12.5, 25, 50, and 100 µg/mL | Anti-metastatic | TGF-β, snail, E-cadherin, smad2↓ | [53] | [39] | |||||||||
| In vitro | A549 | PCs | ≥100 mg/L | Inhibition of cell viability | ROS, MDA, Nrf2↓ HO-1, NQO1, TXNRD1, glutathione, catalase, superoxide dismutase↑ |
[54] | [40] | |||||||||
| In vitro | A549 | PCs | 12.5, 25, 50, 100, and 200 µM | Inhibition of cell viability and proliferation, apoptosis, cell cycle arrest | N-cadherin, vimentin, Bcl-2, MMP2/9, JAK2/STAT3↓, Bax↑ |
[55] | [41] | |||||||||
| In vitro | A549 | Green tea leaf | 1, 5, 10, 20 µM | Anti-proliferative, apoptosis, cell cycle arrest | P21, P53, Fas/sFasL, Fas/APO-1↑ | [56] | [42] | |||||||||
| In vitro | A549 | Myrica rubra | 0.5, 2.5, 5, and 10 µM | Anti-proliferative, apoptosis, cell cycle arrest | Fas/APO-1, P21/WAF1, P53, Fas/sFasL↑ | [57] | [43] | |||||||||
| In vitro | A549, H460 | Rhododendron formosanum | 125, 150, and 175 µM | Autophagy | Akt/mTOR↓ | [58] | [44] |
3. Anthocyanins in Nicotine-Induced NSCLC Treatment
ACNs-rich extracts from plant-based foods/medicinal plants at various nontoxic concentrations have exerted therapeutic effects against nicotine-induced NSCLC, as demonstrated by several studies. Delphinidin and cyanidin have been shown to inhibit platelet-derived growth factor (PDGF)(AB)-induced VEGF expression in vascular smooth muscle cells through attenuation of the p38 MAPK and c-JUN NH2-terminal kinase (JNK) signaling pathways [59][45], known as α7nAChR-mediated cascades involved in NSCLC.ACNs extracted from Vitis coignetiae Pulliat inhibit the expression of several transcription and growth factors (e.g., MMP2/9 and VEGF) involved in proliferation, angiogenesis, invasion, and migration of NSCLC cells [60][46]. The extracts from Morus alba L. (cyanidin 3-rutinoside and cyanidin 3-glucoside) inhibit the migratory and invasive activities of NSCLC cells [61][47]. Cyanidin-3-glucoside not only inhibits the proliferation, invasion, and migration, but also induces apoptosis. The mechanism underlying such an effect is associated with inhibiting p53-induced gene 3 (TP53I3) expression in NSCLC cells and the downregulation of cellular signaling pathways (PI3K/Akt/mTOR) involved in NSCLC progression [62][48]. Delphinidin inhibits tumor growth, angiogenesis, proliferation, and induces apoptosis in NSCLC cells using the in vivo Matrigel plug assay and the in vitro MTT/ELISA assay by upregulating pro-apoptotic expression (e.g., Bax and caspase 3/9), along with downregulating epidermal growth factor receptor (EGFR), cobalt chloride (CoCl2)-induced HIF-1α, Bcl-2, PCNA, cyclin D1, and VEGF mRNA expression via inhibiting of several signaling pathways [63,64][49][50]. A combination of five ACN extracts (i.e., delphinidin, peonidin, petunidin, cyanidin, and malvidin) from bilberry and blueberry resulted in inhibited growth and metastasis, and induced apoptosis and cell cycle arrest of NSCLC cells in vitro. The mechanism of action of ACNs involves inhibiting activation of multiple signaling pathways, including TNFα-induced NF-kB, Notch, Wnt/β-catenin, and their key transcription factors (i.e., cyclin D1/B1, VEGF, p-ERK, bcl-2, and MMP9). Furthermore, when evaluated with an in vivo model using the in vivo xenograft assay, delphinidin alone, and the ACN mixture, resulted in significantly inhibited growth of H1299 cells [65][51]. ACNs derived from Syzygium cumini L. (known as Indian blackberry) showed significant anti-proliferative effects on NSCLC cells, but the mechanisms for this action have not been observed [66][52]. These results suggest that ACNs may have a significant role in anti-proliferative, anti-invasive, anti-angiogenic, anti-metastatic, and apoptotic/autophagic effects in NSCLC cells by suppressing the activation of key transcription/growth factors and α7nAChR-mediatedcellular signaling pathways. The molecular mechanisms of ACNs in nicotine-induced NSCLC treatment are summarized in Table 2.| Study Type | NSCLC Cell Type | Extract/Compound | Concentrations | Activity | Mechanisms of Action | Reference | |
|---|---|---|---|---|---|---|---|
| In vitro | A549 | Vitis coignetiae Pulliat | 200 µg/mL | Anti-proliferative, anti-invasive, anti-angiogenic, anti-migration | MMP2/9, cyclin D1, C-myc, COX-2, VEGF↓ | [60] | [46] |
| In vitro | A549 | Morus alba | L. | 25, 50, and 100 µM | Anti-migration, anti-invasive | MMP2, c-Jun, C-fos, NF-kB↓ |
References
- Bray, F.; Jacques, F.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424.
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. 2008, 83, 584–594.
- Alsharairi, N.A. The effects of dietary supplements on asthma and lung cancer risk in smokers and non-smokers: A review of the literature. Nutrients 2019, 11, 725.
- Xue, J.; Yang, S.; Seng, S. Mechanisms of cancer induction by tobacco-specific NNK and NNN. Cancers 2014, 6, 1138–1156.
- Hecht, S.S. Lung Carcinogenesis by tobacco smoke. Int. J. Cancer 2012, 131, 2724–2732.
- Hecht, S.S.; Stepanov, I.; Carmella, S.G. Exposure and metabolic activation biomarkers of carcinogenic tobacco-spesficnitrosommines. ACS Chem. Res. 2016, 49, 106–114.
- Tournier, J.-M.; Birembaut, P. Nicotinic acetylcholine receptors and predisposition to lung cancer. Curr. Opin. Oncol. 2011, 23, 83–87.
- Warren, G.W.; Singh, A.K. Nicotine and lung cancer. J. Carcinog. 2013, 12, 1.
- Schaal, S.; Chellappan, S.P. Nicotine-mediated cell proliferation and tumor progression in smoking-related cancers. Mol. Cancer Res. 2014, 12, 14–23.
- Wang, S.; Hu, Y. α7 nicotinic acetylcholine receptors in lung cancer (Review). Oncol. Lett. 2018, 16, 1375–1382.
- Zhao, H.; Wang, Y.; Ren, X. Nicotine promotes the development of non-small cell lung cancer through activating LINC00460 and PI3K/Akt signaling. Biosci. Rep. 2019, 39, BSR20182443.
- Hajiasgharzadeh, K.; Sadigh-Eteghad, S.; Mansoori, B.; Mokhtarzadeh, A.; Shanehbandi, D.; Doustvandi, M.A.; Asadzadeh, Z.; Baradaran, B. Alpha7 nicotinic acetylcholine receptors in lung inflammation and carcinogenesis: Friends or foes? J. Cell Physiol. 2019, 234, 14666–14679.
- Alsharairi, N.A. Scutellaria baicalensis and their natural flavone compounds as potential medicinal drugs for the treatment of nicotine-induced non-small-cell lung cancer and asthma. Int. J. Environ. Res. Public Health 2021, 18, 5243.
- Alsharairi, N.A. Supplements for smoking-related lung diseases. Encyclopedia 2021, 1, 76–86.
- Dafni, U.; Tsourti, Z.; Vervita, K.; Peters, S. Immune checkpoint inhibitors, alone or in combination with chemotherapy, as first-line treatment for advanced non-small cell lung cancer. A systematic review and network meta-analysis. Lung Cancer 2019, 134, 127–140.
- Zanoaga, O.; Braicu, C.; Jurj, A.; Rusu, A.; Buiga, R.; Berindan-Neagoe, I. Progress in research on the role of flavonoids in lung cancer. Int. J. Mol. Sci. 2019, 20, 4291.
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive compounds and antioxidant activity in different types of berries. Int. J. Mol. Sci. 2015, 16, 24673–24706.
- Sayago-Ayerdi, S.; García-Martínez, D.L.; Ramírez-Castillo, A.C.; Ramírez-Concepción, H.R.; Viuda-Martos, M. Tropical fruits and their co-products as bioactive compounds and their health effects: A review. Foods 2021, 10, 1952.
- Samtiya, M.; Aluko, R.E.; Dhewa, T.; Moreno-Rojas, J.M. Potential health benefits of plant food-derived bioactive components: An overview. Foods 2021, 10, 839.
- Smeriglio, A.; Barreca, A.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262.
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A comprehensive review encompassing structure elucidation via mass spectrometry. Phytochem. Rev. 2017, 17, 1–16.
- Mannino, G.; Chinigò, G.; Serio, G.; Genova, T.; Gentile, C.; Munaron, L.; Bertea, C.M. Proanthocyanidins and where to find them: A meta-analytic approach to investigate their chemistry, biosynthesis, distribution, and effect on human health. Antioxidants 2021, 10, 1229.
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779.
- Lin, B.-W.; Gong, C.-C.; Song, H.-F.; Cui, Y.-Y. Effects of anthocyanins on the prevention and treatment of cancer. Br. J. Pharmacol. 2017, 174, 1226–1243.
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A comprehensive review of their chemical properties and health effects on cardiovascular and neurodegenerative diseases. Molecules 2020, 25, 3809.
- Ferguson, P.J.; Kurowska, E.; Freeman, D.J.; Chambers, A.F.; Koropatnick, D.J. A flavonoid fraction from cranberry extract inhibits proliferation of human tumor cell lines. J. Nutr. 2004, 134, 1529–1535.
- Neto, C.C.; Krueger, C.G.; Lamoureaux, T.L.; Kondo, M.; Vaisberg, A.J.; Hurta, R.A.R.; Curtis, S.; Matchett, M.D.; Yeung, H.; Sweeney, M.I.; et al. Maldi-TOF MS characterization of proanthocyanidins from cranberry fruit (Vaccinium macrocarpon) that inhibit tumor cell growth and matrix metalloproteinase expression in vitro. J. Sci. Food. Agric. 2006, 86, 18–25.
- Kresty, L.A.; Howell, A.B.; Baird, M. Cranberry proanthocyanidins mediate growth arrest of lung cancer cells through modulation of gene expression and rapid induction of apoptosis. Molecules 2011, 16, 2375–2390.
- Zhai, W.-Y.; Jia, C.-P.; Zhao, H.; Xu, Y.-S. Procyanidins inhibit tumor angiogenesis by crosslinking extracellular matrix. Chin. J Cancer Res. 2011, 23, 99–106.
- Akhtar, S.; Meeran, S.M.; Katiyar, N.; Katiyar, S.K. Grape seed proanthocyanidins inhibit the growth of human non-small cell lung cancer xenografts by targeting insulin-like growth factor binding protein-3, tumor cell proliferation, and angiogenic factors. Clin. Cancer Res. 2009, 15, 821–831.
- Sharma, S.D.; Meeran, S.M.; Katiyar, S.K. Proanthocyanidins inhibit in vitro and in vivo growth of human non-small cell lung cancer cells by inhibiting the prostaglandin E(2) and prostaglandin E(2) receptors. Mol. Cancer Ther. 2010, 9, 569–580.
- Punathil, T.; Katiyar, S.K. Inhibition of non-small cell lung cancer cell migration by grape seed proanthocyanidins is mediated through the inhibition of nitric oxide, guanylate cyclase, and ERK1/2. Mol. Carcinog. 2009, 48, 232–242.
- Mao, J.T.; Smoake, J.; Park, H.K.; Lu, Q.-Y.; Xue, B. Grape seed procyanidin extract mediates antineoplastic effects against lung cancer via modulations of prostacyclin and 15-HETE eicosanoid pathways. Cancer Prev. Res. 2016, 9, 925–932.
- Vaid, M.; Katiyar, S.K. Grape seed proanthocyanidins inhibit cigarette smoke condensate-induced lung cancer cell migration through inhibition of NADPH oxidase and reduction in the binding of p22(phox) and p47(phox) proteins. Mol. Carcinog. 2015, 54, E61–E71.
- Singh, T.; Sharma, S.D.; Katiyar, S.K. Grape proanthocyanidins induce apoptosis by loss of mitochondrial membrane potential of human non-small cell lung cancer cells in vitro and in vivo. PLoS ONE 2011, 6, e27444.
- Xue, B.; Lu, Q.-Y.; Massie, L.; Qualls, C.; Mao, J.T. Grape seed procyanidin extract against lung cancer: The role of microrna-106b, bioavailability, and bioactivity. Oncotarget 2018, 9, 15579–15590.
- Ohnuma, T.; Anzai, E.; Suzuki, Y.; Shimoda, M.; Saito, S.; Nishiyama, T.; Ogura, K.; Hiratsuka, A. Selective antagonization of activated Nrf2 and inhibition of cancer cell proliferation by procyanidins from Cinnamomi Cortex extract. Arch. Biochem. Biophys. 2015, 585, 17–24.
- Ohnuma, T.; Sakamoto, K.; Shinoda, A.; Takagi, C.; Ohno, S.; Nishiyama, T.; Ogura, K.; Hiratsuka, A. Procyanidins from Cinnamomi Cortex promote proteasome-independent degradation of nuclear Nrf2 through phosphorylation of insulin-like growth factor-1 receptor in A549 cells. Arch. Biochem. Biophys. 2017, 635, 66–73.
- Kin, R.; Kato, S.; Kaneto, N.; Sakurai, H.; Hayakawa, Y.; Li, F.; Tanaka, K.; Saiki, I.; Yokoyama, S. Procyanidin C1 from Cinnamomi Cortex inhibits TGF-β-induced epithelial-to-mesenchymal transition in the A549 lung cancer cell line. Int. J. Oncol. 2013, 43, 1901–1906.
- Sun, C.; Jin, W.; Shi, H. Oligomeric proanthocyanidins protects A549 cells against H2O2-induced oxidative stress via the Nrf2-ARE pathway. Int. J. Mol. Med. 2017, 39, 1548–1554.
- Wu, Y.; Liu, C.; Niu, Y.; Xia, J.; Fan, L.; Wu, Y.; Gao, W. Procyanidins mediates antineoplastic effects against non-small cell lung cancer via the JAK2/STAT3 pathway. Transl. Cancer Res. 2021, 10, 2023–2035.
- Kuo, P.L.; Hsu, Y.L.; Lin, T.C.; Lin, C.C. The antiproliferative activity of prodelphinidin B-2 3’-O-gallate from green tea leaf is through cell cycle arrest and Fas-mediated apoptotic pathway in A549 cells. Food Chem. Toxicol. 2005, 43, 315–323.
- Kuo, P.L.; Hsu, Y.L.; Lin, T.A.; Lin, C.C. Prodelphinidin B-2 3,3’-di-O-gallate from Myrica rubra inhibits proliferation of A549 carcinoma cells via blocking cell cycle progression and inducing apoptosis. Eur. J. Pharmacol. 2004, 501, 41–48.
- Way, T.-D.; Tsai, S.-J.; Wang, C.-M.; Jhan, Y.-L.; Ho, C.-T.; Chou, C.-H. Cinnamtannin D1 from Rhododendron formosanum induces autophagy via the inhibition of Akt/mTOR and activation of ERK1/2 in non-small-cell lung carcinoma cells. J. Agric. Food Chem. 2015, 63, 10407–10417.
- Oak, M.-H.; Bedoui, J.E.; Madeira, S.V.F.; Chalupsky, K.; Schini-Kerth, V.B. Delphinidin and cyanidin inhibit PDGF(AB)-induced VEGF release in vascular smooth muscle cells by preventing activation of p38 MAPK and JNK. Br. J. Pharmacol. 2006, 149, 283–290.
- Lu, J.N.; Panchanathan, R.; Lee, W.S.; Kim, H.J.; Kim, D.H.; Choi, Y.H.; Kim, G.; Shin, S.C.; Hong, S.C. Anthocyanins from the fruit of Vitis coignetiae Pulliatinhibit TNF-augmented cancer proliferation, migration, and invasion in A549 cells. Asian Pac. J. Cancer Prev. 2017, 18, 2919–2923.
- Chen, P.-N.; Chu, S.-C.; Chiou, H.-L.; Kuo, W.-H.; Chiang, C.-L.; Hsieh, Y.-S. Mulberry anthocyanins, cyanidin 3-rutinoside and cyanidin 3-glucoside, exhibited an inhibitory effect on the migration and invasion of a human lung cancer cell line. Cancer Lett. 2006, 235, 248–259.
- Chen, X.; Zhang, W.; Xu, X. Cyanidin-3-glucoside suppresses the progression of lung adenocarcinoma by downregulating TP53I3 and inhibiting PI3K/AKT/mTOR pathway. World J. Surg. Oncol. 2021, 19, 232.
- Pal, H.C.; Sharma, S.; Strickland, L.R.; Agarwal, J.; Athar, M.; Elmets, C.A.; Afaq, F. Delphinidin reduces cell proliferation and induces apoptosis of non-small-cell lung cancer cells by targeting EGFR/VEGFR2 signaling pathways. PLoS ONE. 2013, 8, e77270.
- Kim, M.-H.; Jeong, Y.-J.; Cho, H.-J.; Hoe, H.-S.; Park, K.-K.; Park, Y.-Y.; Choi, Y.-H.; Kim, C.-H.; Chang, H.-W.; Park, Y.-J.; et al. Delphinidin inhibits angiogenesis through the suppression of HIF-1α and VEGF expression in A549 lung cancer cells. Oncol. Rep. 2017, 37, 777–784.
- Kausar, H.; Jeyabalan, J.; Aqil, F.; Chabba, D.; Sidana, J.; Singh, I.P.; Gupta, R.C. Berry anthocyanidins synergistically suppress growth and invasive potential of human non-small-cell lung cancer cells. Cancer Lett. 2012, 325, 54–62.
- Aqil, F.; Gupta, A.; Munagala, R.; Jeyabalan, J.; Kausar, H.; Sharma, R.J.; Singh, I.P.; Gupta, R.C. Antioxidant and antiproliferative activities of anthocyanin/ellagitannin-enriched extracts from Syzygium cumini L. (Jamun, the Indian Blackberry). Nutr. Cancer 2012, 64, 428–438.