Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Amina Yu and Version 1 by Alice Sychrová.
Flavonoids have been shown to be excellent natural agents useful in the treatment of various skin lesions with minimal side effects. Topical application is the best option for targeted use due to their lipophilic nature. On the other hand, the polyhydroxyl structure determines their antibacterial, antifibrotic, antioxidant, and anti-inflammatory properties. Prenylated Flavonoids are a subclass of flavonoids modified with at least one lipophilic side chain of varying length. They attract the attention of scientists because of their promising biological activities, such as antibacterial, antifungal, estrogenic, immunosuppressive, anticancer, anti-inflammatory, antioxidant, antiviral, larvicidal, osteogenic, antiallergic, and cytotoxic.
- antibacterial
- anti-inflammatory
- prenylated flavonoids
- skin
- wound healing
1. Flavonoids as Effective Anti-Infective and Wound Healing Agents
Various flavonoids were mentioned as promising therapeutic agents suitable for wound healing. Flavonoids are widely used natural phenolic compounds. Their structure consists of a 2-phenyl-benzo-γ-pyran nucleus comprising two benzene rings. Depending on the degree of unsaturation and oxidation, they are classified into different subclasses [53][1]. In plants, they fulfil important functions, mainly defensive and regulatory. One of these functions is protection against bacterial and fungal pathogens. This ability has been confirmed by numerous in vitro studies. They have shown that flavonoids not only target the bacterial cell directly but also inhibit virulence factors and biofilm formation, reverse antibiotic resistance, or act synergistically with antibiotics [54][2]. Considering these properties, they have become patterns for semisynthetic or synthetic flavonoids combating microorganisms with MICs below 1 µg/mL. In addition to hydroxyl groups, they have been modified with halogens or other heteroatomic rings, such as pyridine, piperidine, or 1,3-dithiolium cations [55][3].
2. Flavonoids as Candidates for Therapy of Skin Lesions
Flavonoids have been shown to be excellent natural agents useful in the treatment of various skin lesions with minimal side effects [29,56,57][4][5][6]. Topical application is the best option for targeted use due to their lipophilic nature [57][6]. On the other hand, the polyhydroxyl structure determines their antibacterial, antifibrotic, antioxidant, and anti-inflammatory properties. Twenty-four structurally different flavonoids have shown the ability to accelerate healing, with quercetin, epigallocatechin gallate, and naringenin being the most studied [56][5]. The results showed that flavonoids decreased the levels of inflammatory mediators, such as prostaglandin E2 (PGE2), leukotriene B4 (LTB-4), interleukin 1β (IL-1β), tumour necrosis factor α (TNF-α), interleukin 6 (IL-6), interferon γ (IFN-γ); increased anti-inflammatory mediators, especially interleukin 10 (IL-10); downregulated the expression of nuclear factor kappa B (NF-κB); and inhibited the activity of cyclooxygenase (COX). Flavonoids affected cell proliferation, migration, differentiation, and angiogenesis by increasing the expression of matrix metalloproteinases 2, 8, 9, and 13. Vascular endothelial growth factor (VEGF), the main molecule regulating vascular growth, was also increased by various flavonoids. The formation of ROS significantly delays wound healing, but should be arrested by flavonoids, which increase the levels of common antioxidant enzymes. The limitation for clinical use is their low bioavailability, which is now solved by the formation of nanostructures that offer better stability, solubility, ability to cross the skin barrier, site-specific delivery, better pharmacokinetic parameters, and, in addition, a reduction in toxicity and side effects. These novel drug delivery systems in the form of nanoparticles, lipid nanocapsules, microparticles, microsponges, and so on, enable the uptake of both hydrophilic and lipophilic compounds and can be formulated into gels, creams, and other dosage forms [56,57][5][6]. Lipophilicity is a key factor of plant flavonoids against Gram-positive bacteria. The mechanism of action of flavonoids against Gram-positive bacteria likely involves the damage of phospholipid bilayers, the inhibition of the respiratory chain or adenosine triphosphate (ATP) synthesis, or some others [58][7].
3. Prenylated Flavonoids
They are a subclass of flavonoids modified with at least one lipophilic side chain of varying length. They attract the attention of scientists because of their promising biological activities, such as antibacterial, antifungal, estrogenic, immunosuppressive, anticancer, anti-inflammatory, antioxidant [59][8], antiviral, larvicidal, osteogenic, antiallergic, and cytotoxic [60][9]. They are found in roots, barks, seeds, and buds [61][10] of nontoxic or even medicinal and food plants [59][8]. Prenylated flavonoids have been found in the families Moraceae, Fabaceae, Cannabaceae, Guttiferae, Rutaceae, Paulowniaceae, Umbelliferae [62][11], Euphorbiaceae [59][8], Celastraceae [63][12], Asteraceae [64][13], and Thymelaeaceae [65][14]. Some species, such as Artocarpus heterophyllus, Broussonetia papyrifera, Epimedium brevicornum, Glycine max, Glycyrrhiza glabra, Humulus lupulus, and Morus alba, and propolis serve as fruits or vegetables, functional foods, or medicines in the daily diet [66][15]. Among the prenylated flavonoids, C-prenylated chalcones/dihydrochalcones, flavanones, flavones, flavonols, and isoflavones or, less frequently, O-prenylated forms occur. These structures are substituted with 3,3-dimethylallyl, 1,1-dimethylallyl, geranyl, lavandulyl, and farnesyl side chains, which can be modified by oxidation, reduction, dehydration, and/or cyclisation [61][10]. Several studies have shown that the prenyl component offers several advantages compared with parent flavonoids. In general, it causes a higher affinity to the cell membrane at the target site. Prenylated flavonoids are known to be potent P-glycoprotein inhibitors, and these abilities condition greater health-promoting properties [67][16]. In the case of antibacterial and enzyme inhibitory or enhancing functions, prenylation increases lipophilicity, leading to increased affinity for biological membranes and enhanced interaction with target proteins [60,68][9][17]. Cytotoxic tests showed that prenylated flavonoids have a higher binding energy in contrast to simple flavonoids [60][9]. Many prenylated flavonoids fulfil the assumption that they target certain diseases with an effective dose, but without having a toxic effect on their own cells. Therefore, these secondary metabolites are being intensively researched as candidates for novel dietary supplements or drugs [59][8]. The effect of prenylated flavonoids in the body and their pharmacokinetics after oral administration are well described. Prenylated modifications play a crucial role in absorption, tissue distribution, and metabolism. The lipophilic portion worsens the transport in the intestine into the internal circulation but, on the other hand, improves the incorporation of the prenylated flavonoids into the tissue-forming cells. Compared with the parent flavonoids, the prenylated forms are detected in tissues to a greater extent, and their accumulation lasts longer. This may indicate their difficult elimination from tissue-forming cells via efflux pumps or a low rate of glucuronidated forms. In summary, prenylation enhances various biological effects but also carries the risk of potential side effects, especially with long-term dietary intake [67][16]. There is less information on the topical use of prenylated flavonoids. Dong et al. [69][18] tested in vivo the anti-inflammatory activity of sophoraflavanone G, the prenylated flavanone presented in Sophora flavescens, with effects observed after oral and topical administration. Although the potencies of inhibition were far below those of the reference drug prednisolone, sophoraflavanone G showed higher anti-inflammatory activity when applied topically [69][18].
4. Mechanisms of Antibacterial Activity
In general, the structure of 2-phenyl-1,4-benzopyrone is crucial for the antibacterial activity of flavonoids and prenylated flavonoids. The available reports on the mechanisms of antibacterial action have led to different results. It seems possible that flavonoids may not only affect one specific target but also influence several cellular processes. Existing research has suggested that antibacterial activity may be caused by the following mechanisms [70][19].
4.1. Direct Interaction with Bacterial Cell
For apigenin and quercetin, the inhibition of cell wall synthesis has been observed via reversible inhibition of D-alanine–D-alanine (D-Ala–D-Ala) ligase, the essential enzyme important for the ligation of D-Ala–D-Ala in the completion of peptidoglycan precursors [71][20]. Alteration of cell membrane permeability and damage to membrane functions were found for several flavonoids. Direct damage to the bacterial cytoplasmic membrane using hydrogen peroxide were the first mechanisms of action attributed to various flavan-3-ols [72][21], flavolans [73[22][23][24],74,75], and green tea catechins [75,76][24][25]. Flavonoids generate hydrogen peroxide by releasing a hydrogen from their pyrogallol or catechol structure to oxygen via a superoxide anion radical [77][26]. On the other hand, catechins can cause membrane fusion, leading to leakage of intramembranous material and aggregation. Inhibition of membrane function has also been discovered for galangin [78][27] and quercetin [79][28]. Sophoraflavanone G showed that lipophilic flavonoids may be able to reduce the fluidity of the outer and inner layers of cell membranes [80][29]. Other mechanisms of antibacterial action have been described for retrochalcones isolated from Glycyrrhiza inflata as an effect on the biosynthesis of macromolecules. Retrochalcones inhibited the incorporation of thymidine, uracil, and leucine into macromolecules, such as deoxyribonucleic acid (DNA), ribonucleic acid (RNA), and proteins. They also inhibited the oxidation of nicotinamide adenine dinucleotide (NADH) and oxygen consumption in the bacterial membrane. The results suggest an inhibition of respiration between coenzyme Q and cytochrome c in the bacterial electron transport chain [81][30]. The mechanism of ATP synthase or hydrolysis blockade has been described. This enzyme is responsible for ATP generation through phosphorylation and photophosphorylation; therefore, the antibacterial effect is attributed to the inhibition of energy metabolism. This effect is caused by the interaction between the flavonoid and polyphenol binding pocket residues of the enzyme. The size, shape, geometry, and presence of functional groups of the compounds are crucial for the binding and inhibition of the enzyme [82][31]. This mechanism of action could also disrupt bacterial motility [79][28]. For example, seventeen flavonoids have been shown to block ATP synthase and subsequently inhibit energy metabolism in E. coli [82][31]. Another suspected mechanism has been suggested in nucleic acid synthesis. Flavonoids have been identified as promising topoisomerase I inhibitors due to their redox, structural, and steric properties. They must undergo oxidation to quinones and could then interact with the DNA topoisomerase complex [83][32]. The inhibition of DNA gyrase has been found for quercetin and apigenin, for example [84][33], and another flavonoid, rutin, could interact with topoisomerase, in particular, IV [85][34]. Flavones and flavonols were identified as inhibitors of helicases and thus interfere with the process of separation of two cross-linked nucleic acid strands [86][35]. Extensively studied catechins from green tea showed activity against Proteus vulgaris and S. aureus. The mechanism of their action was elucidated using radioactive precursors as flavonoid–DNA intercalation when DNA and protein synthesis RNA inhibition was shown [87][36]. In another study, EGCG was able to affect an important bacterial enzyme, dihydrofolate reductase. In addition, EGCG enhanced the effect of standard inhibitors of folic acid metabolism, such as sulfamethoxazole and ethambutol [88][37]. Many flavonoids can also inhibit bacterial metal enzymes due to their chelating ability [89][38]. Another target of action is to influence fatty acid biosynthesis. It has been published that some flavonoids (e.g., EGCG) are able to inhibit three successive enzymes: β-ketoacyl-ACP reductase (FabG), β-hydroxyacyl-ACP dehydratase (FabZ), and enoyl-ACP reductase (FabI) [90][39].
4.2. Indirect Antimicrobial Activity
The inhibition of bacterial pathogenicity is considered one of the nonspecific mechanisms of antibacterial action. Flavonoids can inhibit the quorum sensing system, which is important for bacterial communication and regulation of virulence factors, including biofilm formation [91][40]. In a study by Vikram et al. [92][41], the citrus flavonoids apigenin, kaempferol, quercetin, and naringenin were highlighted as significant antagonists of cell–cell signalling. The apple flavonoid phloretin reduced the expression of genes involved in toxin production and fimbriae formation [93][42]. Other flavonoids, such as myricetin, quercetin, kaempferol, pinocembrin, catechins, and proanthocyanidins, also neutralise bacterial toxic virulence factors (e.g., hyaluronidase and α-hemolysin [54][2]. The unexpected discovery was the nonspecific aggregating effect of flavonoids on whole cells of bacteria. It has been postulated that the antibacterial effect of flavonoids does not target specific enzymes and may not affect enzymes at all [70,91][19][40]. Nevertheless, this bacterial cell aggregation affects membrane integrity and causes biofilm disruption [94][43]. In summary, studies have led to the realisation that a compound can have multiple mechanisms (see Figure 1).
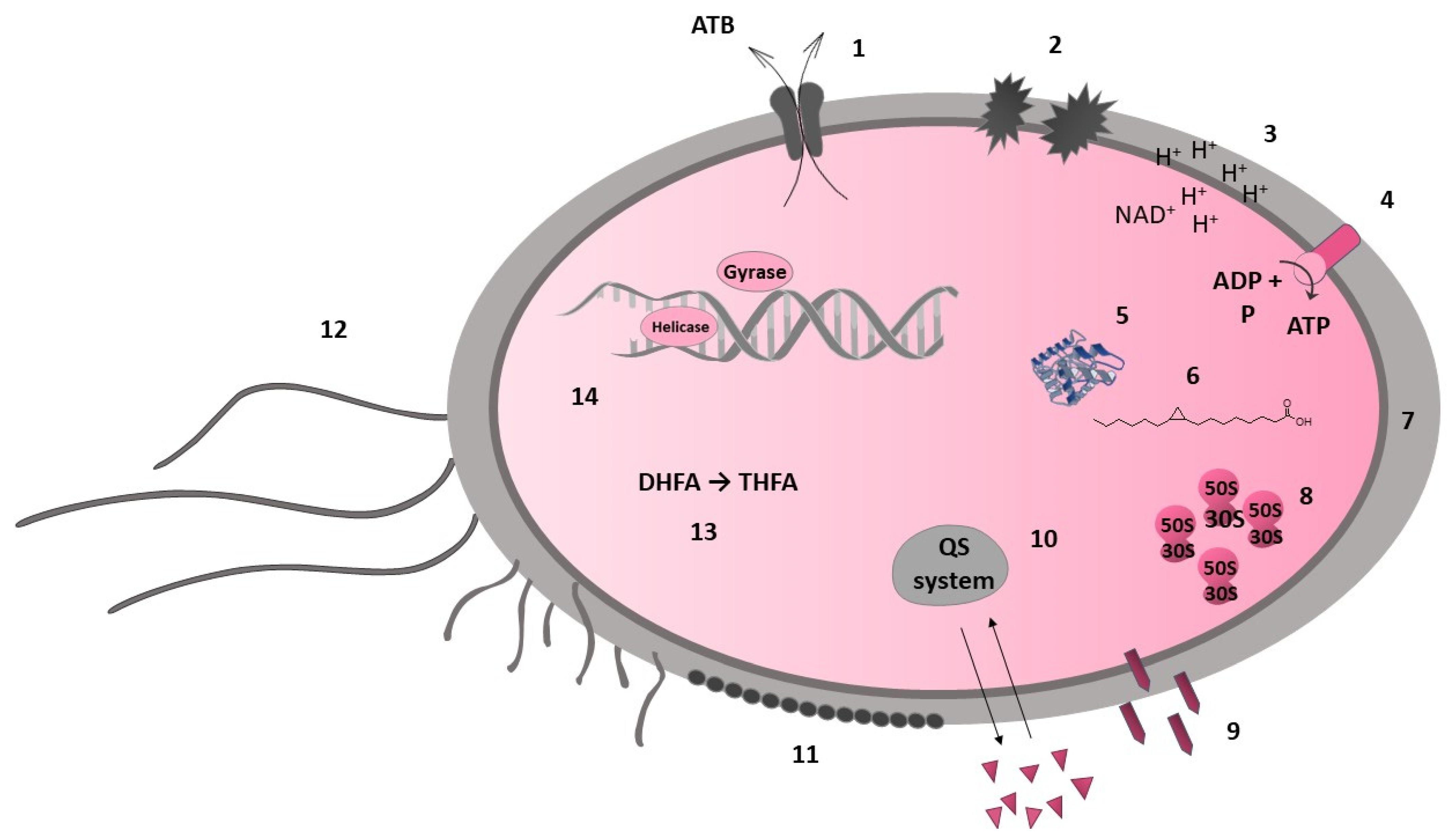
Figure 1. Bacterial targets of flavonoids. Cell membrane: efflux pump (1), membrane disruption (2), electron transport chain (3), ATP synthesis (4); bacterial metalloenzymes (5); fatty acid synthesis (FabG, FabZ, FabI) (6); cell wall synthesis (7): peptidoglycan, D-alanine–D-alanine ligase; protein synthesis (8): (cell envelope); nonspecific mechanism: bacterial toxic virulence factors (9), quorum sensing system (10), biofilm formation (disruption) (11), motility (12); folic acid metabolism: dihydrofolate reductase (13); nucleic acid synthesis (14): DNA gyrase, topoisomerases I and IV, helicase, DNA intercalation.
5. Structure–Activity Relationship
The presence of hydrophobic and hydrophilic moieties is crucial for the antibacterial activity of flavonoids [95][44]. The prenyl group on the condensed pyran ring system normally controls this activity. In contrast, the presence of the prenyl group was not required for aryl substitutions [96][45]. The hydrophobic substituents such as alkylamino and/or alkyl chains and nitrogen- or oxygen-containing heterocyclic moieties usually enhance the antibacterial activity for all flavonoids [97][46]. On the other hand, modifications of the lipophilic side chain with carbonyl, hydroxyl, and methoxyl moieties and/or cyclisation of the prenyl and/or geranyl substituent reduce the activity [97,98][46][47]. The most structurally active compounds include chalcones, flavanones, and flavan-3-ols [99][48]. Antibacterial properties depend on several structural features typical for each class of flavonoids. In general, the hydroxyl groups are able to trigger and enhance the anti-MRSA activity of flavonoids, while the presence of methoxy units drastically decreases the antibacterial activity. Hydroxyl groups in positions 2′ of the chalcones and 5 of the flavanones (or flavones) increase activity against MRSA, while the methoxy groups have the opposite effect. Very promising anti-MRSA activity was measured for 2′(OH)-chalcone, 2′,4′(OH)2-chalcone, and 2′,4(OH)2-chalcone [100][49]. Considering their antibiofilm activity against MRSA, hydroxylations in positions 2′ or 4′ in the A ring and 4 in the B ring also seem to be relevant structural features. Some heterocyclic chalcone analogues have been synthesised with the result that replacing the aromatic ring B with a heterocyclic ring containing nitrogen, oxygen, or sulphur atoms does not significantly increase antibacterial activity against MSSA and MRSA [101][50]. On the other hand, a lipophilic substitution of ring A increases the activity [97][46]. Xie et al. [97][46] summarised that 5,7,4′-hydroxyl substitutions indicate the antibacterial activity of flavones, and their methylation decreases the activity, while flavonols seem to be better antibacterial agents than flavones. Substitutions in the A ring at positions 7 (-O-acyl or -O-alkylamino) and 5-hydroxyl have been reported to be crucial for the antibacterial activity of flavones. Favourable interactions are found in the B ring, where positions 3′ or 4′ are hydroxylated or O-acylated. Hydroxyl or methoxy substituents at position 6′ cause moderate antibiofilm activity [102][51]. Flavanones containing a saturated C3–C4 bond are considered the more promising compounds than flavones [97][46]. Results by Oh et al. (2011) showed that lavandulyl or isoprenyl groups at C-8 contribute to the antibacterial activity of prenylflavanone derivatives [103][52]. The number and position of prenyl/geranyl and hydroxyl groups determine the anti-MRSA activity of flavanones and flavanonols [99][48], with at least C-5, C-7, and C-4′ hydroxylations being basic requirements [98,104][47][53]. Structures with a 2′,4′- or 2′,6′-dihydroxylation of the B ring and substituted with a long-chain aliphatic group such as lavandulyl or geranyl at the 6- or 8-position showed strong activity [97][46]. In contrast, Tsuchiya et al. [104][53] investigated that the lavandulyl group was a more efficient moiety for enhancing antibacterial activity than geranyl. Dihydroflavonols generally show better activity than flavonols, and the compounds with double prenyl substituents are more active than the corresponding monosubstituted ones. Among the flavanols, especially catechins and theaflavins show antibacterial properties against common pathogenic bacteria, such as S. aureus, MRSA, E. coli, and H. pylori [97][46]. In general, oligomeric flavanols show higher activity than monomeric ones, and this rule also applies to flavan trimers compared with dimers [105,106][54][55]. The structure–antimicrobial activity relationship of flavonoids is explained in Figure 2.
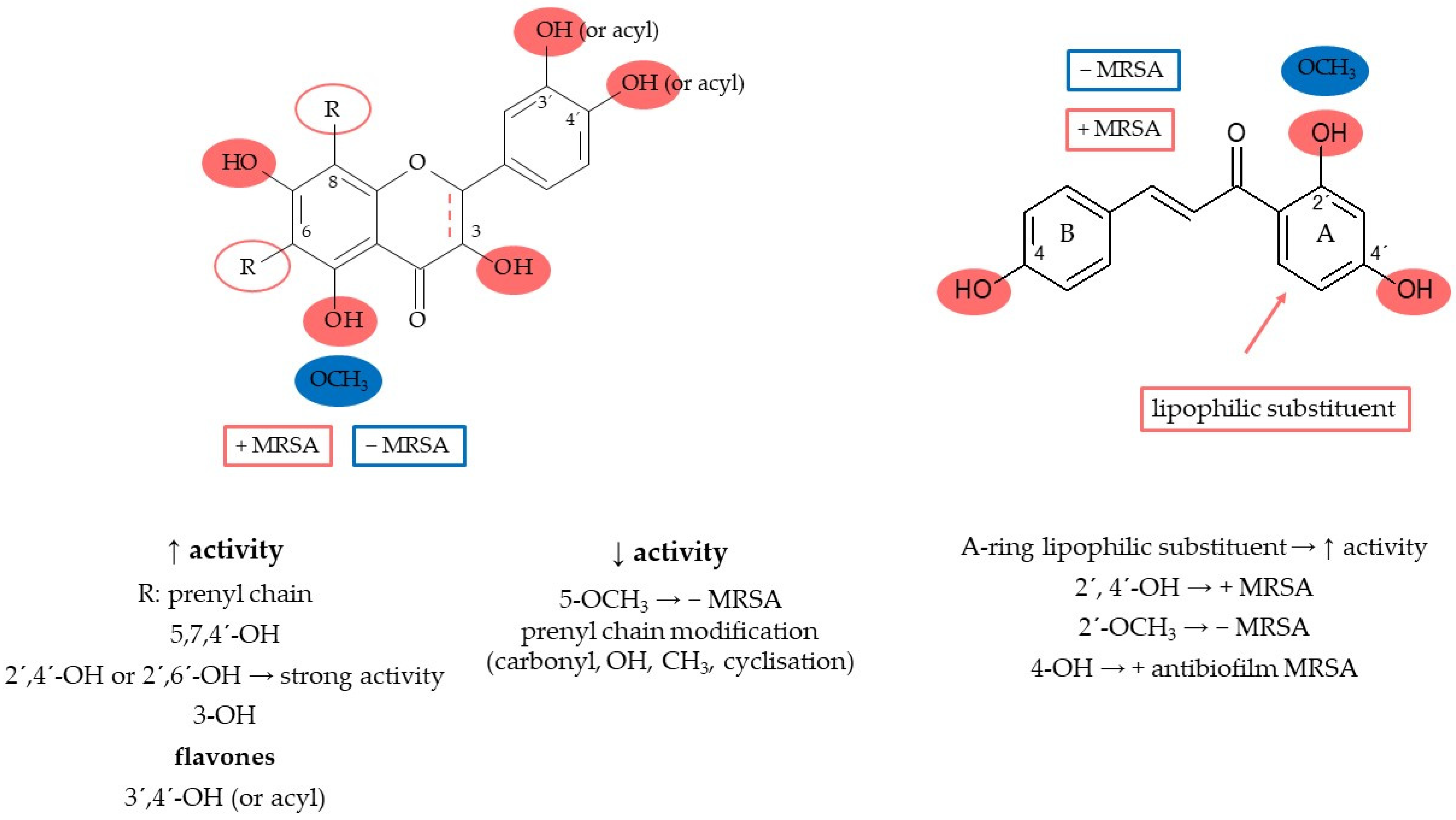
Figure 2.
Flavonoids’ structure–activity relationship.
References
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, Biological Functions, and Biotechnological Applications. Front. Plant Sci. 2012, 3, 222.
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 2019, 18, 241–272.
- Sarbu, L.G.; Bahrin, L.G.; Babii, C.; Stefan, M.; Birsa, M.L. Synthetic Flavonoids with Antimicrobial Activity: A Review. J. Appl. Microbiol. 2019, 127, 1282–1290.
- Tsala, D.E.; Amadou, D.; Habtemariam, S. Natural Wound Healing and Bioactive Natural Products. Phytopharmacology 2013, 4, 532–560.
- Carvalho, M.T.B.; Araújo-Filho, H.G.; Barreto, A.S.; Quintans-Júnior, L.J.; Quintans, J.S.S.; Barreto, R.S.S. Wound Healing Properties of Flavonoids: A Systematic Review Highlighting the Mechanisms of Action. Phytomedicine 2021, 90, 153636.
- Nagula, R.L.; Wairkar, S. Recent Advances in Topical Delivery of Flavonoids: A Review. J. Control. Release 2019, 296, 190–201.
- Yuan, G.; Guan, Y.; Yi, H.; Lai, S.; Sun, Y.; Cao, S. Antibacterial Activity and Mechanism of Plant Flavonoids to Gram-Positive Bacteria Predicted from Their Lipophilicities. Sci. Rep. 2021, 11, 10471.
- Yang, X.; Jiang, Y.; Yang, J.; He, J.; Sun, J.; Chen, F.; Zhang, M.; Yang, B. Prenylated Flavonoids, Promising Nutraceuticals with Impressive Biological Activities. Trends Food Sci. Technol. 2015, 44, 93–104.
- Chen, X.; Mukwaya, E.; Wong, M.S.; Zhang, Y. A Systematic Review on Biological Activities of Prenylated Flavonoids. Pharm. Biol. 2014, 52, 655–660.
- Barron, D.; Ibrahim, R.K. Isoprenylated Flavonoids—A Survey. Phytochemistry 1996, 43, 921–982.
- Šmejkal, K. Cytotoxic potential of C-prenylated flavonoids. Phytochem. Rev. 2014, 13, 245–275.
- Chen, Y.; Zhao, J.; Qiu, Y.; Yuan, H.; Khan, S.I.; Hussain, N.; Iqbal Choudhary, M.; Zeng, F.; Guo, D.A.; Khan, I.A.; et al. Prenylated Flavonoids from the Stems and Roots of Tripterygium wilfordii. Fitoterapia 2017, 119, 64–68.
- Al-Rehaily, A.J.; Albishi, O.A.; El-Olemy, M.M.; Mossa, J.S. Flavonoids and Terpenoids from Helichrysum forskahlii. Phytochemistry 2008, 69, 1910–1914.
- Sun, Q.; Wang, D.; Li, F.F.; Yao, G.D.; Li, X.; Li, L.Z.; Huang, X.X.; Song, S.J. Cytotoxic Prenylated Flavones from the Stem and Root Bark of Daphne giraldii. Bioorganic Med. Chem. Lett. 2016, 26, 3968–3972.
- Chang, S.K.; Jiang, Y.; Yang, B. An Update of Prenylated Phenolics: Food Sources, Chemistry and Health Benefits. Trends Food Sci. Technol. 2021, 108, 197–213.
- Mukai, R. Prenylation Enhances the Biological Activity of Dietary Flavonoids by Altering Their Bioavailability. Biosci. Biotechnol. Biochem. 2018, 82, 207–215.
- Hatano, T.; Shintani, Y.; Aga, Y.; Shiota, S.; Tsuchiya, T.; Yoshida, T. Phenolic Constituents of Licorice. VIII. Structures of Glicophenone and Glicoisoflavanone, and Effects of Licorice Phenolics on Methicillin-Resistant Staphylococcus aureus. Chem. Pharm. Bull. 2000, 48, 1286–1292.
- Dong, W.K.; Yeon, S.C.; Son, K.H.; Hyeun, W.C.; Ju, S.K.; Sam, S.K.; Hyun, P.K. Effects of Sophoraflavanone G, a Prenylated Flavonoid from Sophora flavescens, on Cyclooxygenase-2 and in Vivo Inflammatory Response. Arch. Pharm. Res. 2002, 25, 329–335.
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial Activity of Flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356.
- Wu, D.; Kong, Y.; Han, C.; Chen, J.; Hu, L.; Jiang, H.; Shen, X. D-Alanine:D-Alanine Ligase as a New Target for the Flavonoids Quercetin and Apigenin. Int. J. Antimicrob. Agents 2008, 32, 421–426.
- Sirk, T.W.; Brown, E.F.; Sum, A.K.; Friedman, M. Molecular Dynamics Study on the Biophysical Interactions of Seven Green Tea Catechins with Lipid Bilayers of Cell Membranes. J. Agric. Food Chem. 2008, 56, 7750–7758.
- Kusuda, M.; Inada, K.; Ogawa, T.O.; Yoshida, T.; Shiota, S.; Tsuchiya, T.; Hatano, T. Polyphenolic Constituent Structures of Zanthoxylum piperitum Fruit and the Antibacterial Effects of Its Polymeric Procyanidin on Methicillin-Resistant Staphylococcus aureus. Biosci. Biotechnol. Biochem. 2006, 70, 1423–1431.
- Arakawa, H.; Kanemitsu, M.; Tajima, N.; Maeda, M. Chemiluminescence Assay for Catechin Based on Generation of Hydrogen Peroxide in Basic Solution. Anal. Chim. Acta 2002, 472, 75–82.
- Arakawa, H.; Maeda, M.; Okubo, S.; Shimamura, T. Role of Hydrogen Peroxide in Bactericidal Action of Catechin. Biol. Pharm. Bull. 2004, 27, 277–281.
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal Catechins Damage the Lipid Bilayer. BBA-Biomembr. 1993, 1147, 132–136.
- Miura, Y.H.; Tomita, I.; Watanabe, T.; Hirayama, T.; Fukui, S. Active Oxygens Generation by Flavonoids. Biol. Pharm. Bull. 1998, 21, 93–96.
- Cushnie, T.P.T.; Lamb, A.J. Detection of Galangin-Induced Cytoplasmic Membrane Damage in Staphylococcus aureus by Measuring Potassium Loss. J. Ethnopharmacol. 2005, 101, 243–248.
- Mirzoeva, O.K.; Grishanin, R.N.; Calder, P.C. Antimicrobial Action of Propolis and Some of Its Components: The Effects on Growth, Membrane Potential and Motility of Bacteria. Microbiol. Res. 1997, 152, 239–246.
- Tsuchiya, H.; Iinuma, M. Reduction of Membrane Fluidity by Antibacterial Sophoraflavanone G Isolated from Sophora exigua. Phytomedicine 2000, 7, 161–165.
- Haraguchi, H.; Tanimoto, K.; Tamura, Y.; Mizutani, K.; Kinoshita, T. Mode of Antibacterial Action of Retrochalcones from Glycyrrhiza inflata. Phytochemistry 1998, 48, 125–129.
- Chinnam, N.; Dadi, P.K.; Sabri, S.A.; Ahmad, M.; Kabir, M.A.; Ahmad, Z. Dietary Bioflavonoids Inhibit Escherichia coli ATP Synthase in a Differential Manner. Int. J. Biol. Macromol. 2010, 46, 478–486.
- Bensasson, R.V.; Zoete, V.; Jossang, A.; Bodo, B.; Arimondo, P.B.; Land, E.J. Potency of Inhibition of Human DNA Topoisomerase i by Flavones Assessed through Physicochemical Parameters. Free Radic. Biol. Med. 2011, 51, 1406–1410.
- Ohemeng, K.A.; Schwender, C.F.; Fu, K.P.; Barrett, J.F. DNA Gyrase Inhibitory and Antibacterial Activity of Some Flavones(1). Bioorganic Med. Chem. Lett. 1993, 3, 225–230.
- Bernard, F.X.; Sablé, S.; Cameron, B.; Provost, J.; Desnottes, J.F.; Crouzet, J.; Blanche, F. Glycosylated Flavones as Selective Inhibitors of Topoisomerase IV. Antimicrob. Agents Chemother. 1997, 41, 992–998.
- Xu, H.; Ziegelin, G.; Schröder, W.; Frank, J.; Ayora, S.; Alonso, J.C.; Lanka, E.; Saenger, W. Flavones Inhibit the Hexameric Replicative Helicase RepA. Nucleic Acids Res. 2001, 29, 5058–5066.
- Mori, A.; Nishino, C.; Enoki, N.; Tawata, S. Antibacterial Activity and Mode of Action of Plant Flavonoids against Proteus vulgaris and Staphylococcus aureus. Phytochemistry 1987, 26, 2231–2234.
- Navarro-Martínez, M.D.; Navarro-Perán, E.; Cabezas-Herrera, J.; Ruiz-Gómez, J.; García-Cánovas, F.; Rodríguez-López, J.N. Antifolate Activity of Epigallocatechin Gallate against Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2005, 49, 2914–2920.
- Havsteen, B.H. The Biochemistry and Medical Significance of the Flavonoids. Pharmacol. Ther. 2002, 96, 67–202.
- Tasdemir, D.; Lack, G.; Brun, R.; Rüedi, P.; Scapozza, L.; Perozzo, R. Inhibition of Plasmodium falciparum Fatty Acid Biosynthesis: Evaluation of FabG, FabZ, and FabI as Drug Targets for Flavonoids. J. Med. Chem. 2006, 49, 3345–3353.
- Cushnie, T.P.T.; Lamb, A.J. Recent Advances in Understanding the Antibacterial Properties of Flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107.
- Vikram, A.; Jayaprakasha, G.K.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Suppression of Bacterial Cell-Cell Signalling, Biofilm Formation and Type III Secretion System by Citrus Flavonoids. J. Appl. Microbiol. 2010, 109, 515–527.
- Lee, J.H.; Regmi, S.C.; Kim, J.A.; Cho, M.H.; Yun, H.; Lee, C.S.; Lee, J. Apple Flavonoid Phloretin Inhibits Escherichia coli O157:H7 Biofilm Formation and Ameliorates Colon Inflammation in Rats. Infect. Immun. 2011, 79, 4819–4827.
- Budzyńska, A.; Rózalski, M.; Karolczak, W.; Wieckowska-Szakiel, M.; Sadowska, B.; Rózalska, B. Synthetic 3-Arylidenefl Avanones as Inhibitors of the Initial Stages of biofilm formation by Staphylococcus aureus and Enterococcus faecalis. Z. Naturforschung. C J. Biosci. 2011, 66, 104–114.
- Echeverría, J.; Opazo, J.; Mendoza, L.; Urzúa, A.; Wilkens, M. Structure-Activity and Lipophilicity Relationships of Selected Antibacterial Natural Flavones and Flavanones of Chilean Flora. Molecules 2017, 22, 608.
- Madan, S.; Singh, G.N.; Kohli, K.; Ali, M.; Kumar, Y.; Singh, R.M.; Prakash, O. Isoflavonoids from Flemingia strobilifera (L.) R. Br. Roots. Acta Pol. Pharm.-Drug Res. 2009, 66, 297–303.
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial Activities of Flavonoids: Structure-Activity Relationship and Mechanism. Curr. Med. Chem. 2014, 22, 132–149.
- Navrátilová, A.; Schneiderová, K.; Veselá, D.; Hanáková, Z.; Fontana, A.; Dall’acqua, S.; Cvačka, J.; Innocenti, G.; Novotná, J.; Urbanová, M.; et al. Minor C-Geranylated Flavanones from Paulownia tomentosa Fruits with MRSA Antibacterial Activity. Phytochemistry 2013, 89, 104–113.
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial Activity of Flavonoids and Their Structure–Activity Relationship: An Update Review. Phyther. Res. 2019, 33, 13–40.
- Alcaráz, L.E.; Blanco, S.E.; Puig, O.N.; Tomás, F.; Ferretti, F.H. Antibacterial Activity of Flavonoids against Methicillin-Resistant Staphylococcus aureus Strains. J. Theor. Biol. 2000, 205, 231–240.
- Tran, T.D.; Nguyen, T.T.N.; Do, T.H.; Huynh, T.N.P.; Tran, C.D.; Thai, K.M. Synthesis and Antibacterial Activity of Some Heterocyclic Chalcone Analogues Alone and in Combination with Antibiotics. Molecules 2012, 17, 6684–6696.
- Manner, S.; Skogman, M.; Goeres, D.; Vuorela, P.; Fallarero, A. Systematic Exploration of Natural and Synthetic Flavonoids for the Inhibition of Staphylococcus aureus Biofilms. Int. J. Mol. Sci. 2013, 14, 19434–19451.
- Oh, I.; Yang, W.Y.; Chung, S.C.; Kim, T.Y.; Oh, K.B.; Shin, J. In Vitro Sortase A Inhibitory and Antimicrobial Activity of Flavonoids Isolated from the Roots of Sophora flavescens. Arch. Pharm. Res. 2011, 34, 217–222.
- Tsuchiya, H.; Sato, M.; Miyazaki, T.; Fujiwara, S.; Tanigaki, S.; Ohyama, M.; Tanaka, T.; Iinuma, M. Comparative Study on the Antibacterial Activity of Phytochemical Flavanones against Methicillin-Resistant Staphylococcus aureus. J. Ethnopharmacol. 1996, 50, 27–34.
- Idowu, T.O.; Ogundaini, A.O.; Salau, A.O.; Obuotor, E.M.; Bezabih, M.; Abegaz, B.M. Doubly Linked, A-Type Proanthocyanidin Trimer and Other Constituents of Ixora coccinea Leaves and Their Antioxidant and Antibacterial Properties. Phytochemistry 2010, 71, 2092–2098.
- Mayer, R.; Stecher, G.; Wuerzner, R.; Silva, R.C.; Sultana, T.; Trojer, L.; Feuerstein, I.; Krieg, C.; Abel, G.; Popp, M.; et al. Proanthocyanidins: Target Compounds as Antibacterial Agents. J. Agric. Food Chem. 2008, 56, 6959–6966.
More