神经性疼痛是由体感神经系统受累的组织损伤或疾病引起的慢性疼痛,严重影响患者的身体机能和生活质量。皂苷是一类结构多样的化合物,由皂苷元和糖基组成。构成皂苷的糖类常见的有Neuropathic pain is a chronic pain caused by tissue injury or disease involving the somatosensory nervous system, which seriously affects the patient’s body function and quality of life. Saponins are a class of compounds with diverse structures, consisting of sapogenin and glycosyl groups. The common ones of the saccharides that make up saponins are D-葡萄糖、D-半乳糖、D-木糖、L-阿拉伯糖、L-鼠李糖等。 glucose, D-galactose, D-xylose, L-arabinose, and L-rhamnose, etc.
- neuropathic pain
- saponins
- mechanism
1.人参皂甙
1. Ginsenosides
Ginsenosides are the major biologically active components of ,具有广泛的药理活性。根据其苷元的骨架,人参皂苷可分为两类,四环三萜达玛烷型皂苷(原人参二醇(Ginseng, which have a wide range of pharmacological activities. According to the skeleton of their aglycones, ginsenosides can be classified into two groups, tetracyclic triterpene dammarane-type saponins (protopanaxadiol (PPD)型,原人参三醇()-, protopanaxatriol (PPT)型)(图1)-type) (Figure 1A)和四环三萜齐墩果烷型皂苷[) and tetracyclic triterpene oleanane-type 48saponins ,[1][2][3]. 49So far, 50more than ]。迄今为止,已分离出100多种不同的人参皂苷单体,如人参皂苷Rb1、Rb2、Rc、Rd、Re、Rg1和Rf,它们的药理和药代动力学特性各不相同[ different ginsenoside monomers have been isolated, such as ginsenosides Rb1, Rb2, Rc, Rd, Re, Rg1, and Rf, the pharmacological and pharmacokinetic properties of which are 51、52different ]。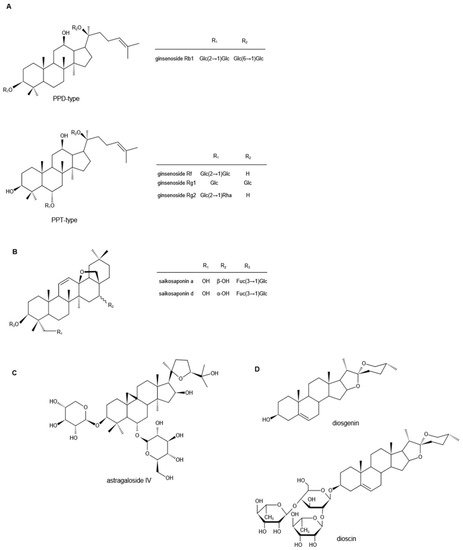
2. Saikosaponins
Saikosaponins are [derived 59 ]],尽管其潜在机制需要进一步探索。迄今为止,人参皂苷在神经性疼痛中的镇痛机制大多局限于对炎症因子的探索,缺乏对其靶向分子靶点的深入分析。此外,人参皂甙的调节作用是否与不同的神经性疼痛相关脑区有关,在很大程度上仍是未知数。围绕这些点开展进一步的研究可为药品的精准调控提供研究依据。2.柴胡皂苷
3. 黄芪甲苷
3. Astragalosides
Astragali Radix, the dried roots of Astragalus membranaceus ((Fisch.)Bge) Bge. 的干燥根 。变种。var. mongholicus (Bge.) Hsiao,或, or 黄芪Astragalus membranaceus (Fisch.) Bge.,被称为高档中药, is known as a high-grade traditional Chinese medicine [47]. There are three main types of compounds in astragalus: saponins, flavonoids, and polysaccharides, and triterpene saponins are the major constituents [48][49][50]. It is reported that more than 40 kinds of saponins have been isolated and identified from the dried [astragalus 81roots ]。黄芪中的化合物主要有三类:皂甙、黄酮和多糖,其中三萜皂甙是主要成分[via 82、83、84]。据报道,通过HPLC和 and GC-MS从干燥的黄芪根中分离鉴定了40多种皂苷,如黄芪甲苷I-VIII、乙酰黄芪甲苷、异黄芪甲苷I、III、黄芪甲素II、环黄芪甲苷、环黄芪苷B、短叶黄芪苷等。, such as astragalosides I–VIII, acetylastragaloside, isoastragaloside I, III, astramembrannin II, cycloastragenol, cycloascauloside B, brachyoside B, astrasieversianin X, 等etc. [[51][52][53][54]. 85Among these, 86astragaloside ,IV 87 , 88 ]。其中,黄芪甲苷((AS-IV)被称为主要活性成分和定性控制生物标志物。) is known as the major active ingredient and qualitative control biomarker. AS-IV是3 -O-β- is 3-O-beta-d-xylopyranosyl-6-O-β--xylopyranosyl-6-O-beta-d-吡喃葡萄糖基-glucopyranosyl-环黄芪醇(图1cycloastragenol (Figure 1C), the molecular formula ),分子式为is C 41 H 68O 14 [55]. It [is generally accepted that the transient receptor 89potential ]。4.薯蓣皂苷元
4. Diosgenin
Diosgenin is a naturally occurring steroidal sapogenin and is abundant in nature. Primary sources of diosgenin include the three 种薯蓣Dioscorea 属物种和一种species and one 异菝葜Heterosmilax 物种,即species, namely, D. zingiberensis、, D. septemloba、, D. collettii和, and H. yunnanensis [70]. [Diosgenin can also be obtained 100from ]。薯蓣皂苷元也可以从胡芦巴fenugreek ( T. foenum graecum Linn ) 和and Costus speciosus [71][72][73]. [It 101is ,a 102C27 ,spiroketal 103steroid ]sapogenin, 中获得。它是一种C27螺缩酮类固醇皂苷元,3β-羟基-5-螺甾烯(图13β-hydroxy-5-spirostene (Figure 1D),其分子式为), and its molecular formula is C 27 H42O3 [74]. OAs a representational phytosteroid, diosgenin is an important basic raw material for the production of steroid hormone drugs and has received increasing attention in the pharmaceutical industry for decades [75]. In addition, diosgenin itself has a wide range of biological effects. The following studies mainly describe its role in neuropathic pain. Neuropathic pain, one of the common complications of diabetes mellitus, manifests as increased sensitivity to noxious stimuli [76]. To evaluate the effects of diosgenin in the treatment of diabetes-induced neuropathic pain, an in 3vivo [study 104was ]。作为具有代表性的植物甾体,薯蓣皂苷元是生产甾体激素类药物的重要基础原料,几十年来越来越受到制药行业的关注[performed 105on ]。此外,薯蓣皂苷元本身具有广泛的生物学作用。以下研究主要描述了其在神经性疼痛中的作用。3.5. O. sanctum 富含皂苷的提取物
5. Saponin-Rich Extracts of O. sanctum
除了上述四种已鉴定出结构清晰的植物皂苷的镇痛作用外,还发现了富含皂苷的O. sanctum提取物具有相似的作用。O. sanctum是唇形科植物罗勒的地上部分。现代药理研究表明,O. sanctum化学成分复杂,种类多样,包括挥发油、黄酮类及其苷类、香豆素类、苯丙素类和脂肪酸类,主要是挥发油和黄酮类及其苷类[111]。此外,已从O. sanctum的酒精提取物中分离出多种皂苷。[112],其中最重要的是以熊果酸和齐墩果酸为主的五环三萜皂苷[113-115],具有广泛的药理作用。
In addition to the analgesic effects of the above four plant saponins that have been identified with clear structures, saponin-rich extracts of O. sanctum have also been found with similar effects. O. sanctum is the aerial part of Ocimum basilicum, a plant of the Labiatae family. Modern pharmacological studies have illustrated that the chemical composition of O. sanctum is complex and the types are diverse, including volatile oils, flavonoids and their glycosides, coumarins, phenylpropanoids, and fatty acids, mainly volatile oils and flavonoids and their glycosides [91]. In addition, a variety of saponins have been isolated from the alcoholic extract of O. sanctum [92], the most important of which are pentacyclic triterpenoid saponins that are dominated by ursolic and oleanolic acids [93][94][95], and have a wide range of pharmacological effects.氧化应激 [116] 和钙稳态的改变 [117] 被认为与神经性疼痛密切相关。在神经系统疾病期间,细胞内钙调节系统的功能障碍会产生氧化应激 [118],而自由基的增加会导致神经元变性和细胞凋亡。另一方面,由氧化应激引起的代谢异常[119]、蛋白质聚集体的形成[120]和膜通透性的变化[121]都会增加钙水平,它们共同作用促进神经性疼痛的恶化。O. sanctum具有良好的抗氧化作用 [122,123],可防止自由基损伤 [124],并且能够降低钙水平 [125]。O. sanctum在印度部分地区用作神经补品,用于缓解头痛、关节痛和肌肉痛。在 Muthuraman 等人进行的实验中,O. sanctum的给药减轻了坐骨神经横断引起的周围神经病变和运动协调,减弱了截肢引起的硫代巴比妥酸活性物质、总钙和谷胱甘肽水平的降低以剂量依赖的方式[125]。提示O. sanctum的镇痛作用可能与其抗氧化和降低钙水平有关。此外,在其他研究中,用O. sanctum治疗其富含皂苷的部分可减轻由慢性缩窄性损伤和化学治疗剂长春新碱引起的神经性疼痛,这与其对氧化应激和钙水平的影响有关[125,126]。基于上述发现,可以观察到,O. sanctum给药对钙水平的下调可能是由于直接影响或继发于氧化应激的降低。然后,它对神经元产生镇痛或抗凋亡作用。据报道,皂苷具有抗氧化作用 [127] 和降钙作用 [128]。因此,O. sanctum皂苷的镇痛作用可以通过直接或间接降低钙水平来构建。
Oxidative stress [96] and alterations in calcium homeostasis [97] are thought to be closely associated with neuropathic pain. During neurological disorders, dysfunction of the intracellular calcium regulatory system produces oxidative stress [98], and increases in free radicals lead to neuronal degeneration and apoptosis. On the other hand, metabolic abnormalities [99], formation of protein aggregates [100], and changes in membrane permeability [101] caused by oxidative stress all increase calcium levels, and they act together to promote the deterioration of neuropathic pain. O. sanctum has a good antioxidant effect [102][103], protects against free radical damage [104], and is able to reduce calcium levels [105]. O. sanctum is used as a neurotonic in parts of India for the relief of headache, joint pain, and muscle pain. In the experiments conducted by Muthuraman et al., the administration of O. sanctum attenuated sciatic nerve transection-induced peripheral neuropathy and motor in-co-ordination, attenuated the amputation-induced reduction in thiobarbituric acid reactive species, total calcium, and glutathione levels in a dose-dependent manner [105]. It suggested that the analgesic effect of O. sanctum might be related to its antioxidation and reduction of calcium levels. Additionally, in other studies, treatment with O. sanctum and its saponin-rich fraction reduced neuropathic pain caused by chronic constrictive injury and chemotherapeutic agent vincristine, associated with its effects on the oxidative stress and calcium levels [105][106]. Based on the above findings, it can be observed that the downregulation of calcium levels by O. sanctum administration may be due to a direct effect on or secondary to a decrease in oxidative stress. Then, it produces an antinociceptive or antiapoptotic effect on neurons. It has been reported that Saponins have antioxidant [107] and calcium lowering effects [108]. Thus, the antinociceptive effect of O. sanctum saponins may be constructed through direct or indirect reduction of calcium levels.还有证据表明,O. sanctum叶子和种子可降低兔子的尿酸水平 [129],而尿酸水平升高与痛风性关节炎和其他关节炎症有关 [130]。O. sanctum的乙醇提取物具有镇痛作用,并涉及神经递质系统(如阿片受体和去甲肾上腺素)的相互作用 [131]。这些研究支持传统使用O. sanctum治疗炎症和疼痛,但不排除其他活性成分如黄酮类和酚类的作用。
There is also evidence that O. sanctum leaves and seeds reduce uric acid levels in rabbits [109], and elevated uric acid levels are associated with gouty arthritis and other joint inflammation [110]. The ethanolic extract of O. sanctum can be antinociceptive, and involves the interaction of neurotransmitter systems such as opioid receptors and norepinephrine [111]. These studies support the traditional use of O. sanctum for the treatment of inflammation and pain, without excluding the effects of other active ingredients such as flavonoids and phenols.References
- Christensen, L.P. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 2009, 55, 1–99.
- Zheng, M.; Xin, Y.; Li, Y.; Xu, F.; Xi, X.; Guo, H.; Cui, X.; Cao, H.; Zhang, X.; Han, C. Ginsenosides: A Potential Neuroprotective Agent. BioMed Res. Int. 2018, 2018, 8174345.
- Hou, M.; Wang, R.; Zhao, S.; Wang, Z. Ginsenosides in Panax genus and their biosynthesis. Acta Pharm. Sin. B 2021, 11, 1813–1834.
- Lee, H.S.; Lee, H.J.; Yu, H.J.; Ju do, W.; Kim, Y.; Kim, C.T.; Kim, C.J.; Cho, Y.J.; Kim, N.; Choi, S.Y.; et al. A comparison between high hydrostatic pressure extraction and heat extraction of ginsenosides from ginseng (Panax ginseng CA Meyer). J. Sci. Food Agric. 2011, 91, 1466–1473.
- Nag, S.A.; Qin, J.J.; Wang, W.; Wang, M.H.; Wang, H.; Zhang, R. Ginsenosides as Anticancer Agents: In vitro and in vivo Activities, Structure-Activity Relationships, and Molecular Mechanisms of Action. Front. Pharmacol. 2012, 3, 25.
- Lee, J.Y.; Choi, H.Y.; Park, C.S.; Kim, D.H.; Yune, T.Y. Total saponin extract, ginsenoside Rb1, and compound K alleviate peripheral and central neuropathic pain through estrogen receptors on rats. Phytother. Res. 2021, 35, 2119–2132.
- Gao, C.; Guo, X.; Weng, L.; Zhou, W.; Shen, Y.; Yu, Y.; Han, Y. Effect of ginsenoside Rg1 on activity of spinal microglia and expression of p38 mitogen-activated protein kinase/nuclear transcription factor-kappa B in neuropathic pain rats. Int. J. Anesth. Resus 2017, 38, 1084.
- Li, Y.; Chen, C.; Li, S.; Jiang, C. Ginsenoside Rf relieves mechanical hypersensitivity, depression-like behavior, and inflammatory reactions in chronic constriction injury rats. Phytother. Res. 2019, 33, 1095–1103.
- Wolf, G.; Yirmiya, R.; Goshen, I.; Iverfeldt, K.; Holmlund, L.; Takeda, K.; Shavit, Y. Impairment of interleukin-1 (IL-1) signaling reduces basal pain sensitivity in mice: Genetic, pharmacological and developmental aspects. Pain 2003, 104, 471–480.
- Miyoshi, K.; Obata, K.; Kondo, T.; Okamura, H.; Noguchi, K. Interleukin-18-mediated microglia/astrocyte interaction in the spinal cord enhances neuropathic pain processing after nerve injury. J. Neurosci. 2008, 28, 12775–12787.
- Ji, R.R.; Chamessian, A.; Zhang, Y.Q. Pain regulation by non-neuronal cells and inflammation. Science 2016, 354, 572–577.
- Sommer, C.; Leinders, M.; Üçeyler, N. Inflammation in the pathophysiology of neuropathic pain. Pain 2018, 159, 595–602.
- Huang, F.; Li, Y.N.; Yin, F.; Wu, Y.T.; Zhao, D.X.; Li, Y.; Zhang, Y.F.; Zhu, Q.S. Ginsenoside Rb1 inhibits neuronal apoptosis and damage, enhances spinal aquaporin 4 expression and improves neurological deficits in rats with spinal cord ischemiareperfusion injury. Mol. Med. Rep. 2015, 11, 3565–3572.
- Gao, X.-Q.; Yang, C.-X.; Chen, G.-J.; Wang, G.-Y.; Chen, B.; Tan, S.-K.; Liu, J.; Yuan, Q.-L. Ginsenoside Rb1 regulates the expressions of brain-derived neurotrophic factor and caspase-3 and induces neurogenesis in rats with experimental cerebral ischemia. J. Ethnopharmacol. 2010, 132, 393–399.
- Adamo, D.; Calabria, E.; Coppola, N.; Pecoraro, G.; Mignogna, M.D. Vortioxetine as a new frontier in the treatment of chronic neuropathic pain: A review and update. Ther. Adv. Psychopharmacol. 2021, 11, 1–19.
- Zhang, Q.L.; Li, S.Y.; Li, P. Effects of ginsenoside-Rg2 on mechanical allodynia, heat hyperalgeia, depressive state of rats with chronic sciatic nerve constriction injury. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2019, 35, 228–231.
- Ruizhen, C.; Chunyan, Z. Research progress on pharmacological activities of Saikosaponins from Radix Bupleurum. Occup. Health 2021, 37, 568–576.
- He, Y.; Hu, Z.; Li, A.; Zhu, Z.; Yang, N.; Ying, Z.; He, J.; Wang, C.; Yin, S.; Cheng, S. Recent Advances in Biotransformation of Saponins. Molecules 2019, 24, 2365.
- Li, X.; Li, X.; Huang, N.; Liu, R.; Sun, R. A comprehensive review and perspectives on pharmacology and toxicology of saikosaponins. Phytomedicine 2018, 50, 73–87.
- Ashour, M.L.; Wink, M. Genus Bupleurum: A review of its phytochemistry, pharmacology and modes of action. J. Pharm. Pharm. 2011, 63, 305–321.
- Ebata, N.; Nakajima, K.; Hayashi, K.; Okada, M.; Maruno, M. Saponins from the root of Bupleurum falcatum. Phytochemistry 1996, 41, 895–901.
- Li, X.Q.; Song, Y.N.; Wang, S.J.; Rahman, K.; Zhu, J.Y.; Zhang, H. Saikosaponins: A review of pharmacological effects. J. Asian Nat. Prod. Res. 2018, 20, 399–411.
- Wong, V.K.; Zhou, H.; Cheung, S.S.; Li, T.; Liu, L. Mechanistic study of saikosaponin-d (Ssd) on suppression of murine T lymphocyte activation. J Cell Biochem. 2009, 107, 303–315.
- Wang, Y.L.; He, S.X.; Luo, J.Y. Progress in research on antitumor activity of saikosaponin and its mechanism. J. Chin. Integr. Med. 2006, 4, 98–100.
- Lee, G.; Choi, J.; Nam, Y.J.; Song, M.J.; Kim, J.K.; Kim, W.J.; Kim, P.; Lee, J.S.; Kim, S.; No, K.T.; et al. Identification and characterization of saikosaponins as antagonists of transient receptor potential A1 channel. Phytother. Res. 2020, 34, 788–795.
- Gupta, R.; Saito, S.; Mori, Y.; Itoh, S.G.; Okumura, H.; Tominaga, M. Structural basis of TRPA1 inhibition by HC-030031 utilizing species-specific differences. Sci. Rep. 2016, 6, 37460.
- Wang, S.; Kobayashi, K.; Kogure, Y.; Yamanaka, H.; Yamamoto, S.; Yagi, H.; Noguchi, K.; Dai, Y. Negative Regulation of TRPA1 by AMPK in Primary Sensory Neurons as a Potential Mechanism of Painful Diabetic Neuropathy. Diabetes 2018, 67, 98–109.
- Nativi, C.; Gualdani, R.; Dragoni, E.; Di Cesare Mannelli, L.; Sostegni, S.; Norcini, M.; Gabrielli, G.; la Marca, G.; Richichi, B.; Francesconi, O.; et al. A TRPA1 antagonist reverts oxaliplatin-induced neuropathic pain. Sci. Rep. 2013, 3, 2005.
- Nassini, R.; Gees, M.; Harrison, S.; De Siena, G.; Materazzi, S.; Moretto, N.; Failli, P.; Preti, D.; Marchetti, N.; Cavazzini, A.; et al. Oxaliplatin elicits mechanical and cold allodynia in rodents via TRPA1 receptor stimulation. Pain 2011, 152, 1621–1631.
- Descoeur, J.; Pereira, V.; Pizzoccaro, A.; Francois, A.; Ling, B.; Maffre, V.; Couette, B.; Busserolles, J.; Courteix, C.; Noel, J.; et al. Oxaliplatin-induced cold hypersensitivity is due to remodelling of ion channel expression in nociceptors. EMBO Mol. Med. 2011, 3, 266–278.
- Anderson, D.W.; Bradbury, K.A.; Schneider, J.S. Broad neuroprotective profile of nicotinamide in different mouse models of MPTP-induced parkinsonism. Eur. J. Neurosci. 2008, 28, 610–617.
- Lee, G.; Nam, Y.-J.; Kim, W.J.; Shin, B.H.; Lee, J.S.; Park, H.T.; Kim, P.; Lee, J.H.; Choi, Y. Saikosaponin D Ameliorates Mechanical Hypersensitivity in Animal Models of Neuropathic Pain. Planta Med. Int. Open 2020, 7, e145–e149.
- Li, Z.Y.; Jiang, Y.M.; Liu, Y.M.; Guo, Z.; Shen, S.N.; Liu, X.M.; Pan, R.L. Saikosaponin D acts against corticosterone-induced apoptosis via regulation of mitochondrial GR translocation and a GR-dependent pathway. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 53, 80–89.
- Yuan, B.; Yang, R.; Ma, Y.; Zhou, S.; Zhang, X.; Liu, Y. A systematic review of the active saikosaponins and extracts isolated from Radix Bupleuri and their applications. Pharm. Biol. 2017, 55, 620–635.
- Lin, X.; Wu, S.; Wang, Q.; Shi, Y.; Liu, G.; Zhi, J.; Wang, F. Saikosaponin-D Reduces H2O2-Induced PC12 Cell Apoptosis by Removing ROS and Blocking MAPK-Dependent Oxidative Damage. Cell Mol. Neurobiol. 2016, 36, 1365–1375.
- Sun, T.; Song, W.G.; Fu, Z.J.; Liu, Z.H.; Liu, Y.M.; Yao, S.L. Alleviation of neuropathic pain by intrathecal injection of antisense oligonucleotides to p65 subunit of NF-κB. Br. J. Anaesth. 2006, 97, 553–558.
- Zhou, X.; Cheng, H.; Xu, D.; Yin, Q.; Cheng, L.; Wang, L.; Song, S.; Zhang, M. Attenuation of neuropathic pain by saikosaponin a in a rat model of chronic constriction injury. Neurochem. Res. 2014, 39, 2136–2142.
- Ramesh, G.; MacLean, A.G.; Philipp, M.T. Cytokines and chemokines at the crossroads of neuroinflammation, neurodegeneration, and neuropathic pain. Mediat. Inflamm. 2013, 2013, 480739.
- Lu, C.N.; Yuan, Z.G.; Zhang, X.L.; Yan, R.; Zhao, Y.Q.; Liao, M.; Chen, J.X. Saikosaponin a and its epimer saikosaponin d exhibit anti-inflammatory activity by suppressing activation of NF-kappaB signaling pathway. Int. Immunopharmacol. 2012, 14, 121–126.
- Zhu, J.; Luo, C.; Wang, P.; He, Q.; Zhou, J.; Peng, H. Saikosaponin A mediates the inflammatory response by inhibiting the MAPK and NF-kappaB pathways in LPS-stimulated RAW 264.7 cells. Exp. Ther. Med. 2013, 5, 1345–1350.
- Mao, X.; Miao, G.; Tao, X.; Hao, S.; Zhang, H.; Li, H.; Hou, Z.; Tian, R.; Lu, T.; Ma, J.; et al. Saikosaponin a protects TBI rats after controlled cortical impact and the underlying mechanism. Am. J. Transl. Res. 2016, 8, 133–141.
- Yoon, S.S.; Seo, J.W.; Ann, S.H.; Kim, H.Y.; Kim, H.S.; Cho, H.Y.; Yun, J.; Chung, E.Y.; Koo, J.S.; Yang, C.H. Effects of saikosaponin A on cocaine self-administration in rats. Neurosci. Lett. 2013, 555, 198–202.
- Yoon, S.S.; Kim, H.S.; Cho, H.Y.; Yun, J.; Chung, E.Y.; Jang, C.G.; Kim, K.J.; Yang, C.H. Effect of saikosaponin A on maintenance of intravenous morphine self-administration. Neurosci. Lett. 2012, 529, 97–101.
- Ahmadimoghaddam, D.; Zarei, M.; Mohammadi, S.; Izadidastenaei, Z.; Salehi, I. Bupleurum falcatum L. alleviates nociceptive and neuropathic pain: Potential mechanisms of action. J. Ethnopharmacol. 2021, 273, 113990.
- Minett, M.S.; Nassar, M.A.; Clark, A.K.; Passmore, G.; Dickenson, A.H.; Wang, F.; Malcangio, M.; Wood, J.N. Distinct Nav1.7-dependent pain sensations require different sets of sensory and sympathetic neurons. Nat. Commun. 2012, 3, 791.
- Xu, Y.; Yu, Y.; Wang, Q.; Li, W.; Zhang, S.; Liao, X.; Liu, Y.; Su, Y.; Zhao, M.; Zhang, J. Active components of Bupleurum chinense and Angelica biserrata showed analgesic effects in formalin induced pain by acting on Nav1.7. J. Ethnopharmacol. 2021, 269, 113736.
- Chen, Z.; Liu, L.; Gao, C.; Chen, W.; Vong, C.T.; Yao, P.; Yang, Y.; Li, X.; Tang, X.; Wang, S.; et al. Astragali Radix (Huangqi): A promising edible immunomodulatory herbal medicine. J. Ethnopharmacol. 2020, 258, 112895.
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S.; Zhang, H.; Yang, S. Review of the botanical characteristics, phytochemistry, and pharmacology of Astragalus membranaceus (Huangqi). Phytother. Res. 2014, 28, 1275–1283.
- Song, J.Z.; Mo, S.F.; Yip, Y.K.; Qiao, C.F.; Han, Q.B.; Xu, H.X. Development of microwave assisted extraction for the simultaneous determination of isoflavonoids and saponins in radix astragali by high performance liquid chromatography. J. Sep. Sci. 2007, 30, 819–824.
- Ma, X.Q.; Shi, Q.; Duan, J.A.; Dong, T.T.; Tsim, K.W. Chemical analysis of Radix Astragali (Huangqi) in China: A comparison with its adulterants and seasonal variations. J. Agric. Food Chem. 2002, 50, 4861–4866.
- Wang, Y.; Liu, L.; Ma, Y.; Guo, L.; Sun, Y.; Liu, Q.; Liu, J. Chemical Discrimination of Astragalus mongholicus and Astragalus membranaceus Based on Metabolomics Using UHPLC-ESI-Q-TOF-MS/MS Approach. Molecules 2019, 24, 4064.
- Liu, D.-l.; Bao, H.-Y.; Liu, Y. Progress on Chemical Constituents and Pharmacological Effects of Astragali Radix in Recent Five Years. Food Drug 2014, 16, 68–70.
- Polat, E.; Bedir, E.; Perrone, A.; Piacente, S.; Alankus-Caliskan, O. Triterpenoid saponins from Astragalus wiedemannianus Fischer. Phytochemistry 2010, 71, 658–662.
- Verotta, L.; Guerrini, M.; El-Sebakhy, N.A.; Assad, A.M.; Toaima, S.M.; Radwan, M.M.; Luo, Y.D.; Pezzuto, J.M. Cycloartane and oleanane saponins from egyptian astragalus spp. as modulators of lymphocyte proliferation. Planta Med. 2002, 68, 986–994.
- Zhang, J.; Wu, C.; Gao, L.; Du, G.; Qin, X. Astragaloside IV derived from Astragalus membranaceus: A research review on the pharmacological effects. Adv. Pharm. 2020, 87, 89–112.
- Yang, F.; Xiao, X.; Lee, B.H.; Vu, S.; Yang, W.; Yarov-Yarovoy, V.; Zheng, J. The conformational wave in capsaicin activation of transient receptor potential vanilloid 1 ion channel. Nat. Commun. 2018, 9, 2879.
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543.
- Mitchell, K.; Bates, B.D.; Keller, J.M.; Lopez, M.; Scholl, L.; Navarro, J.; Madian, N.; Haspel, G.; Nemenov, M.I.; Iadarola, M.J. Ablation of rat TRPV1-expressing Adelta/C-fibers with resiniferatoxin: Analysis of withdrawal behaviors, recovery of function and molecular correlates. Mol. Pain 2010, 6, 94.
- Li, X.; Kang, L.; Li, G.; Zeng, H.; Zhang, L.; Ling, X.; Dong, H.; Liang, S.; Chen, H. Intrathecal leptin inhibits expression of the P2X2/3 receptors and alleviates neuropathic pain induced by chronic constriction sciatic nerve injury. Mol. Pain 2013, 9, 65.
- Lin, J.; Li, G.; Den, X.; Xu, C.; Liu, S.; Gao, Y.; Liu, H.; Zhang, J.; Li, X.; Liang, S. VEGF and its receptor-2 involved in neuropathic pain transmission mediated by P2X2/3 receptor of primary sensory neurons. Brain Res. Bull. 2010, 83, 284–291.
- Shi, G.B.; Fan, R.; Zhang, W.; Yang, C.; Wang, Q.; Song, J.; Gao, Y.; Hou, M.X.; Chen, Y.F.; Wang, T.C.; et al. Antinociceptive activity of astragaloside IV in the animal model of chronic constriction injury. Behav. Pharm. 2015, 26, 436–446.
- Dong, Z.Q.; Ma, F.; Xie, H.; Wang, Y.Q.; Wu, G.C. Down-regulation of GFRalpha-1 expression by antisense oligodeoxynucleotide attenuates electroacupuncture analgesia on heat hyperalgesia in a rat model of neuropathic pain. Brain Res. Bull. 2006, 69, 30–36.
- Dong, Z.Q.; Ma, F.; Xie, H.; Wang, Y.Q.; Wu, G.C. Changes of expression of glial cell line-derived neurotrophic factor and its receptor in dorsal root ganglions and spinal dorsal horn during electroacupuncture treatment in neuropathic pain rats. Neurosci. Lett. 2005, 376, 143–148.
- Cheng, C.Y.; Yao, C.H.; Liu, B.S.; Liu, C.J.; Chen, G.W.; Chen, Y.S. The role of astragaloside in regeneration of the peripheral nerve system. J. Biomed. Mater. Res. A 2006, 76, 463–469.
- Zhang, X.; Chen, J. The mechanism of astragaloside IV promoting sciatic nerve regeneration. Neural Regen. Res. 2013, 8, 2256–2265.
- Shen, Y.; Meiri, K. GAP-43 dependency defines distinct effects of netrin-1 on cortical and spinal neurite outgrowth and directional guidance. Int. J. Dev. Neurosci. 2012, 31, 11–20.
- Mendonca, H.R.; Araujo, S.E.; Gomes, A.L.; Sholl-Franco, A.; da Cunha Faria Melibeu, A.; Serfaty, C.A.; Campello-Costa, P. Expression of GAP-43 during development and after monocular enucleation in the rat superior colliculus. Neurosci. Lett. 2010, 477, 23–27.
- Yu, J.; Zhang, Y.; Sun, S.; Shen, J.; Qiu, J.; Yin, X.; Yin, H.; Jiang, S. Inhibitory effects of astragaloside IV on diabetic peripheral neuropathy in rats. Can. J. Physiol. Pharm. 2006, 84, 579–587.
- Davidson, J.A. Treatment of the patient with diabetes: Importance of maintaining target HbA(1c) levels. Curr. Med. Res. Opin. 2004, 20, 1919–1927.
- Yi, T.; Fan, L.L.; Chen, H.L.; Zhu, G.Y.; Suen, H.M.; Tang, Y.N.; Zhu, L.; Chu, C.; Zhao, Z.Z.; Chen, H.B. Comparative analysis of diosgenin in Dioscorea species and related medicinal plants by UPLC-DAD-MS. BMC Biochem. 2014, 15, 19.
- Arya, P.; Kumar, P. Diosgenin a steroidal compound: An emerging way to cancer management. J. Food Biochem. 2021, 45, e14005.
- Chen, Y.; Tang, Y.-M.; Yu, S.-L.; Han, Y.-W.; Kou, J.-P.; Liu, B.-L.; Yu, B.-Y. Advances in the pharmacological activities and mechanisms of diosgenin. Chin. J. Nat. Med. 2015, 13, 578–587.
- Al-Habori, M.; Raman, A.; Lawrence, M.J.; Skett, P. In vitro effect of fenugreek extracts on intestinal sodium-dependent glucose uptake and hepatic glycogen phosphorylase A. Int. J. Exp. Diabetes Res. 2001, 2, 91–99.
- Fan, R.; He, W.; Fan, Y.; Xu, W.; Xu, W.; Yan, G.; Xu, S. Recent advances in chemical synthesis, biocatalysis, and biological evaluation of diosgenin derivatives—A review. Steroids 2022, 180, 108991.
- Fernandes, P.; Cruz, A.; Angelova, B.; Pinheiro, H.M.; Cabral, J.M.S. Microbial conversion of steroid compounds: Recent developments. Enzym. Microb. Tech. 2003, 32, 688–705.
- Obrosova, I.G. Update on the pathogenesis of diabetic neuropathy. Curr. Diabetes Rep. 2003, 3, 439–445.
- Kiasalari, Z.; Rahmani, T.; Mahmoudi, N.; Baluchnejadmojarad, T.; Roghani, M. Diosgenin ameliorates development of neuropathic pain in diabetic rats: Involvement of oxidative stress and inflammation. Biomed. Pharm. 2017, 86, 654–661.
- Kasznicki, J.; Kosmalski, M.; Sliwinska, A.; Mrowicka, M.; Stanczyk, M.; Majsterek, I.; Drzewoski, J. Evaluation of oxidative stress markers in pathogenesis of diabetic neuropathy. Mol. Biol. Rep. 2012, 39, 8669–8678.
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocr. Rev. 2004, 25, 612–628.
- Wang, C.; Ning, L.P.; Wang, Y.H.; Zhang, Y.; Ding, X.L.; Ge, H.Y.; Arendt-Nielsen, L.; Yue, S.W. Nuclear factor-kappa B mediates TRPV4-NO pathway involved in thermal hyperalgesia following chronic compression of the dorsal root ganglion in rats. Behav. Brain Res. 2011, 221, 19–24.
- Zhang, Y.P.; Song, C.Y.; Yuan, Y.; Eber, A.; Rodriguez, Y.; Levitt, R.C.; Takacs, P.; Yang, Z.; Goldberg, R.; Candiotti, K.A. Diabetic neuropathic pain development in type 2 diabetic mouse model and the prophylactic and therapeutic effects of coenzyme Q10. Neurobiol. Dis. 2013, 58, 169–178.
- Kumar, A.; Negi, G.; Sharma, S.S. Suppression of NF-κB and NF-κB regulated oxidative stress and neuroinflammation by BAY 11-7082 (IκB phosphorylation inhibitor) in experimental diabetic neuropathy. Biochimie 2012, 94, 1158–1165.
- Kawamura, N.; Dyck, P.J.; Schmeichel, A.M.; Engelstad, J.K.; Low, P.A.; Dyck, P.J. Inflammatory mediators in diabetic and non-diabetic lumbosacral radiculoplexus neuropathy. Acta Neuropathol. 2008, 115, 231–239.
- Gao, M.; Chen, L.; Yu, H.; Sun, Q.; Kou, J.; Yu, B. Diosgenin down-regulates NF-kappaB p65/p50 and p38MAPK pathways and attenuates acute lung injury induced by lipopolysaccharide in mice. Int. Immunopharmacol. 2013, 15, 240–245.
- Gao, Z.; Feng, Y.; Ju, H. The Different Dynamic Changes of Nerve Growth Factor in the Dorsal Horn and Dorsal Root Ganglion Leads to Hyperalgesia and Allodynia in Diabetic Neuropathic Pain. Pain Physician 2017, 20, E551–E561.
- Calissano, P.; Amadoro, G.; Matrone, C.; Ciafrè, S.; Marolda, R.; Corsetti, V.; Ciotti, M.T.; Mercanti, D.; Di Luzio, A.; Severini, C.; et al. Does the term ‘trophic’ actually mean anti-amyloidogenic? The case of NGF. Cell Death Differ. 2010, 17, 1126–1133.
- Kang, T.H.; Moon, E.; Hong, B.N.; Choi, S.Z.; Son, M.; Park, J.H.; Kim, S.Y. Diosgenin from Dioscorea nipponica ameliorates diabetic neuropathy by inducing nerve growth factor. Biol. Pharm. Bull. 2011, 34, 1493–1498.
- Zhao, W.X.; Wang, P.F.; Song, H.G.; Sun, N. Diosgenin attenuates neuropathic pain in a rat model of chronic constriction injury. Mol. Med. Rep. 2017, 16, 1559–1564.
- Lee, B.K.; Kim, C.J.; Shin, M.S.; Cho, Y.S. Diosgenin improves functional recovery from sciatic crushed nerve injury in rats. J. Exerc. Rehabil. 2018, 14, 566–572.
- Byun, Y.H.; Lee, M.H.; Kim, S.S.; Kim, H.; Chang, H.K.; Lee, T.H.; Jang, M.H.; Shin, M.C.; Shin, M.S.; Kim, C.J. Treadmill running promotes functional recovery and decreases brain-derived neurotrophic factor mRNA expression following sciatic crushed nerve injury in rats. J. Sports Med. Phys. Fit. 2005, 45, 222–228.
- Liu, M.; Luo, F.; Qing, Z.; Yang, H.; Liu, X.; Yang, Z.; Zeng, J. Chemical Composition and Bioactivity of Essential Oil of Ten Labiatae Species. Molecules 2020, 25, 4862.
- Palida, A.; Mi, R.; Cong, Y.; Yi, B.; Wang, X. Isolation and identification of chemical constituents in Ocimum bacilicum. West China J. Pharm. Sci. 2007, 22, 489–490.
- Kaur, G.; Bali, A.; Singh, N.; Jaggi, A.S. Ameliorative potential of Ocimum sanctum in chronic constriction injury-induced neuropathic pain in rats. An. Acad. Bras. Ciênc. 2015, 87, 417–429.
- Anandjiwala, S.; Kalola, J.; Rajani, M. Quantification of eugenol, luteolin, ursolic acid, and oleanolic acid in black (Krishna Tulasi) and green (Sri Tulasi) varieties of Ocimum sanctum Linn. using high-performance thin-layer chromatography. J. AOAC Int. 2006, 89, 1467–1474.
- Rao, A.R.; Veeresham, C.; Asres, K. In vitro and in vivo inhibitory activities of four Indian medicinal plant extracts and their major components on rat aldose reductase and generation of advanced glycation endproducts. Phytother. Res. 2013, 27, 753–760.
- Carrasco, C.; Naziroǧlu, M.; Rodríguez, A.B.; Pariente, J.A. Neuropathic Pain: Delving into the Oxidative Origin and the Possible Implication of Transient Receptor Potential Channels. Front. Physiol. 2018, 9, 95.
- Bourinet, E.; Altier, C.; Hildebrand, M.E.; Trang, T.; Salter, M.W.; Zamponi, G.W. Calcium-permeable ion channels in pain signaling. Physiol. Rev. 2014, 94, 81–140.
- Katsuyama, Y.; Sato, Y.; Okano, Y.; Masaki, H. Intracellular oxidative stress induced by calcium influx initiates the activation of phagocytosis in keratinocytes accumulating at S-phase of the cell cycle after UVB irradiation. J. Dermatol. Sci. 2021, 103, 41–48.
- Gibson, G.E. Interactions of oxidative stress with cellular calcium dynamics and glucose metabolism in Alzheimer’s disease. Free Radic. Biol. Med. 2002, 32, 1061–1070.
- Goodwin, J.; Nath, S.; Engelborghs, Y.; Pountney, D.L. Raised calcium and oxidative stress cooperatively promote alpha-synuclein aggregate formation. Neurochem. Int. 2013, 62, 703–711.
- Carbonera, D.; Azzone, G.F. Permeability of inner mitochondrial membrane and oxidative stress. Biochim. Biophys. Acta 1988, 943, 245–255.
- George, S.; Chaturvedi, P. Protective role of Ocimum canum plant extract in alcohol-induced oxidative stress in albino rats. Br. J. Biomed. Sci. 2008, 65, 80–85.
- Kelm, M.A.; Nair, M.G.; Strasburg, G.M.; DeWitt, D.L. Antioxidant and cyclooxygenase inhibitory phenolic compounds from Ocimum sanctum Linn. Phytomedicine 2000, 7, 7–13.
- Balanehru, S.; Nagarajan, B. Protective effect of oleanolic acid and ursolic acid against lipid peroxidation. Biochem. Int. 1991, 24, 981–990.
- Muthuraman, A.; Diwan, V.; Jaggi, A.S.; Singh, N.; Singh, D. Ameliorative effects of Ocimum sanctum in sciatic nerve transection-induced neuropathy in rats. J. Ethnopharmacol. 2008, 120, 56–62.
- Kaur, G.; Jaggi, A.S.; Singh, N. Exploring the potential effect of Ocimum sanctum in vincristine-induced neuropathic pain in rats. J. Brachial Plex. Peripher. Nerve Inj. 2010, 5, 3.
- Hu, S.; Wu, Y.; Zhao, B.; Hu, H.; Zhu, B.; Sun, Z.; Li, P.; Du, S. Panax notoginseng Saponins Protect Cerebral Microvascular Endothelial Cells against Oxygen-Glucose Deprivation/Reperfusion-Induced Barrier Dysfunction via Activation of PI3K/Akt/Nrf2 Antioxidant Signaling Pathway. Molecules 2018, 23, 2781.
- Neco, P.; Rose, B.; Huynh, N.; Zhang, R.; Bridge, J.H.; Philipson, K.D.; Goldhaber, J.I. Sodium-calcium exchange is essential for effective triggering of calcium release in mouse heart. Biophys. J. 2010, 99, 755–764.
- Sarkar, A.; Pandey, D.N.; Pant, M.C. A report on the effects of Ocimum sanctum (Tulsi) leaves and seeds on blood and urinary uric acid, urea and urine volume in normal albino rabbits. Indian J. Physiol. Pharmacol. 1990, 34, 61–62.
- Dalbeth, N.; Choi, H.K.; Joosten, L.A.B.; Khanna, P.P.; Matsuo, H.; Perez-Ruiz, F.; Stamp, L.K. Gout. Nat. Rev. Dis. Prim. 2019, 5, 69.
- Khanna, N.; Bhatia, J. Antinociceptive action of Ocimum sanctum (Tulsi) in mice: Possible mechanisms involved. J. Ethnopharmacol. 2003, 88, 293–296.