Melanocytes produce melanin to protect the skin from 黑素细胞产生黑色素,以保护皮肤免受UV-B radiation. Notwithstanding, the spectrum of their functions extends far beyond their well-known role as melanin production factories. Melanocytes have been considered as sensory and computational cells. The neurotransmitters, neuropeptides, and other hormones produced by melanocytes make them part of the skin’s well-orchestrated and complex neuroendocrine network, counteracting environmental stressors. Melanocytes can also actively mediate the epidermal immune response. Melanocytes are equipped with ectopic sensory systems similar to the eye and nose and can sense light and odor.In addition, melanocytes have also been shown to be localized in internal sites such as the inner ear, brain, and heart, locations not stimulated by sunlight.辐射。尽管如此,它们的功能范围远远超出了它们作为黑色素生产工厂的众所周知的作用。黑素细胞被认为是感觉和计算细胞。黑素细胞产生的神经递质,神经肽和其他激素使它们成为皮肤精心策划和复杂的神经内分泌网络的一部分,抵消环境压力源。黑素细胞也可以主动介导表皮免疫反应。黑素细胞具有类似于眼睛和鼻子的异位感觉系统,可以感知光和气味。此外,黑素细胞也被证明位于内耳,大脑和心脏等内部部位,这些部位不受阳光刺激。
- melanocytes
- neuroendocrinology
- circadian rhythm
- photoentrainment
- homeostasis
- regulatory network
- sensory functions
- opsins
- extracutaneous pigment cell
- innate immunity
1. Introduction
2. Melanocytes and the Endocrine System
2.1. Hypothalamic-Pituitary-Adrenal (HPA) Axis Homolog
2.2. Hypothalamic-Pituitary-Thyroid Axis (HPT) Homolog
2.3. Serotoninergic/Melatoninergic System
2.4. Other Neuroendocrine Activities
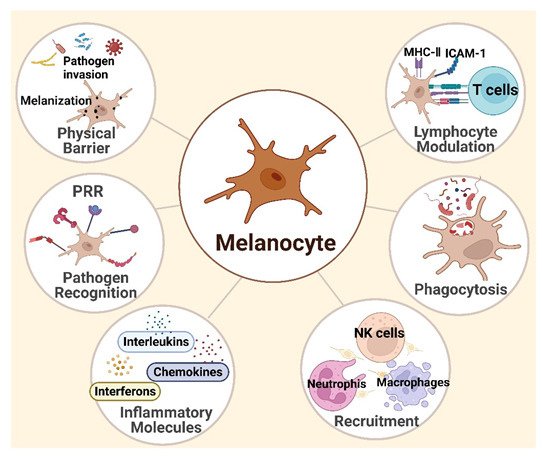
3. Melanocytes and Skin Immunity
3.1. Melanocytes Are Functional Immune Sentinel Cells in the Skin
3.2. Innate Immune Responses
3.3. Adaptive Immunity
4. Melanocytes and the Sensory System
4.1. The Melanocyte Photosensory System
4.1.1. Phototransduction
4.1.2. Circadian Photoentrainment
| Opsin | Species | Potential Effect | Reference |
|---|---|---|---|
| OPN1 | Mouse | Not shown | [146] |
| Human | Not shown | [127][129] | |
| OPN2 | Mouse | Mediate UVA-induced immediate pigment darkening | [126][135][147] |
| Regulate clock genes and melanogenesis responding to white light | |||
| Human | Mediate UVR-induced early melanin synthesis | [127][128][129] | |
| OPN3 | Human | Sense blue light and regulate long-lasting hyperpigmentation | [129][130][131][132][133] |
| Negatively regulate pigmentation through interaction with MC1R | |||
| Regulate the survival of melanocytes | |||
| OPN4 | Mouse | Mediate UVA-induced immediate pigment darkening | [146][147][148] |
| Regulate clock genes and melanogenesis responding to white light | |||
| Mediate UVA-related proliferation and apoptosis | |||
| Mediate thermal activation of clock genes | |||
| Human | Photoreceptor of blue light | [124] | |
| OPN5 | Mouse | Local circadian photoentrainment | [145] |
| Human | Regulate UVR-induced melanogenesis | [129] [130] |
4.2. The Melanocyte Olfactory System
Olfaction is the principal chemosensory system in many organisms and detects diverse smells [105][108]. Odor perception starts with odorant molecules binding to specific olfactory receptors (ORs), which are primarily expressed in olfactory sensory neurons, and then proper chemical signals are transmitted to the brain via electrical signals [105][108]. Numerous reports have described that ORs are also ectopically expressed in multiple extra-nasal tissues. These ORs have generally not been thought to have any significant impact on the pathology of any common disease and have limited potential as therapeutic strategies. Nevertheless, this viewpoint is drastically and rapidly changing as increasing ectopic ORs, responding to various ligands, have been reported to be involved in distinct biological processes including sperm chemotaxis, wound healing, hair growth, muscle regeneration, tumor suppression, and energy metabolism [109][154][155][156][157][158]. The skin is the outermost barrier of the human body and is regularly exposed to multiple environmental chemical stimuli. Several ORs have been identified in diverse cell types housed in the skin, including melanocytes and keratinocytes [154][155][159][160][161]. The expression of diverse cutaneous chemical-sensitive receptors contributes to decoding environmental odor information [116][159]. The developing olfactory system originates from both the embryonic ectodermal placode and the neural crest [162]. Intriguingly, ORs have also been found in keratinocytes and melanocytes that precisely originate from surface ectodermal cells and neural crest cells during the embryonic period, respectively [163]. In the 1980s, the first evidence of the effects of odor on pigment cells was presented in frog melanophores dispersing their melanosomes in response to the same chemical class and concentrations of odorants used in olfactory cilia studies, and this process was correlated to increasing intracellular cAMP [164]. At that time, the superfamily of odor receptors was still unknown. Corresponding studies found melanosome dispersion in fish melanophores responding to the stimulus of cinnamaldehyde and β-ionone [165]. Later, the expression of OR51E2, an olfactory receptor which is located in the olfactory epithelium, was also detected at the transcript or protein levels [161]. It has been further demonstrated that activation of OR51E2 by a specific agonist, β-ionone, ultimately inhibits melanocyte proliferation and stimulates melanogenesis [161]. It is possible that β-ionone, commonly found in beauty care products, may contribute to cosmetic-induced hyperpigmentation [161]. Soon afterward, OR2A4/7 was verified to be expressed and play an active role in primary human melanocytes. The odorant cyclohexyl salicylate, a specific ligand of OR2A4/7, has been shown to inhibit proliferation and promote cell differentiation in conjunction with enhanced pigmentation in melanocytes by elevating intracellular cAMP and Ca2+ levels [162]. Together, these findings imply that several ORs may be potential targets for pigmentation disorder treatment [161][162]. In addition, as OR expression is upregulated in several cancer tissues, the promising application of ORs in cancer diagnosis and therapeutics has also emerged [108]. In particular, OR51E2 serves as a potential biomarker for melanoma [108]. The previous study identified that human melanoma expresses the olfactory receptor 51E2 and further showed that activating endogenous OR51E2 in cultured cells isolated from metastatic and vertical growth phases can suppress the growth of melanoma via inducing apoptosis [166]. In addition, research evaluating the expression profile of olfactory receptors in a multitude of cancer cell lines has shown that OR2C3 is exclusively expressed in melanoma lines [167]. Remarkably, the expression of OR2C3 was confirmed in human melanoma cells but in not normal melanocytes [167]. Therefore, the pattern of OR2C3 expression implies a potential role in the initiation and/or development of melanoma [166][167]. Thus, human melanocytes can “smell” in the sense that they recruit the evolutionarily ancient and largest of receptor families to regulate skin pigmentation, cell proliferation, apoptosis, and tumorigenesis. Although informative, most of the insights into ORs’ role come from studies performed in vitro cell-based models, leaving several biologic questions unanswered. The expression levels of OR transcripts are low; therefore, further confirmation of the function of these ectopic ORs in vivo is urgently needed. Furthermore, it is critical to confirm the role of these receptors in humans. For verifying functions, especially in vitro, most experiments rely on ligand-induced activation of ORs, and few ligands have been identified. The molecular basis of the specificity and sensitivity of ORs is poorly understood. Therefore, the lack of knowledge of the ligands of ORs is an obstacle that needs to be overcome for future research. Furthermore, the identification of endogenous ligands is challenging, and humans may not always share the same ligands with mouse receptors. Moreover, odorant levels sufficient to sustain chronic excitation of the receptor are also worthy of consideration for future clinical applications. In addition, are melanocytes able to transmit signals to the brain like olfactory epithelial cells after sensing an odor? Although the current understanding of the biological function, signaling cascades, and pharmacology of ectopic ORs remains insufficient and problematic and may represent merely the tip of the iceberg, future in-depth exploration and recognition of the potential roles of ORs may lead to the use of potential odorants as a promising strategy for the treatment of various diseases based on the theory of ectopically expressed ORs.5. Melanocytes Outside the Skin
Melanocytes predominantly inhabit the skin epidermis, hair follicles, and the iris of the eye, where they have various functions including producing melanin for protection from ultra-violet radiation and are responsible for the coloration of the skin, hair, and eyes in animals, which is of crucial importance in various aspects of the animal’s life, including communication and visual camouflage 黑素细胞主要栖息在皮肤表皮,毛囊和眼睛的虹膜中,它们具有各种功能,包括产生黑色素以防止紫外线辐射,并负责动物皮肤,头发和眼睛的着色,这在动物生活的各个方面至关重要,包括交流和视觉伪装[1]. Meanwhile, melanocytes play vital roles in embryonic development and organ functions, as can be seen in patients with oculocutaneous albinism type 同时,黑素细胞在胚胎发育和器官功能中起着至关重要的作用,这可以在1 (OCA1), which is caused by mutations of the TYR and leads to hypopigmentation in the skin, hair, and eyes, consequently bringing actinic damage and a higher risk of skin cancer 型眼皮白化病(OCA1)患者中看到,这是由TYR突变引起的,并导致皮肤,头发和眼睛的色素减退,从而带来光化损伤和更高的皮肤癌风险[168]. Most melanocytes originate in neural crests, follow diverse and interesting developmental pathways, and finally migrate to specific areas of the developing embryo. Interestingly, melanocytes are also located in deeper parts of the body that are not directly exposed to 大多数黑素细胞起源于神经嵴,遵循多样化和有趣的发育途径,最后迁移到发育中的胚胎的特定区域。有趣的是,黑素细胞也位于身体更深处不直接暴露于UVR, including in the striatum vessels of the inner ear, in the substantia nigra, leptomeninges, and locus coeruleus of the brain 的部位,包括内耳纹状体血管,黑质,软脑膜和大脑的腔肠位点[163]. In addition, these extracutaneous melanocytes are present in other lesser-known regions, including cardiac valves, the septum, and major arteries and veins in the heart. Therefore, this begs the question of what key roles do these melanocytes in such a sun-protected habitat play in still producing melanin? Here, researchers briefly summarize the function of the extracutaneous melanocytes, primarily focusing on the heart and ears to observe the structural significance they contribute.此外,这些皮外黑素细胞存在于其他鲜为人知的区域,包括心脏瓣膜、鼻中隔以及心脏的主要动脉和静脉。因此,这就引出了一个问题,即在这样一个受阳光保护的栖息地中,这些黑素细胞在仍然产生黑色素方面起着什么关键作用?在这里,我们简要总结了皮外黑素细胞的功能,主要集中在心脏和耳朵上,以观察它们所贡献的结构意义。5.1. Heart
5.1. 心脏
The presence of pigment cells in the heart was first mentioned in a 在PET⁄MCV mouse strain, although it could not clearly indicate their origin or whether they were melanocytes /MCV小鼠品系中首次提到了心脏中色素细胞的存在,尽管它不能清楚地表明它们的起源或它们是否是黑素细胞。[169]. 最近,More recently, Mjaatvedt et al. discovered a group of pigment cells located in the atrium, mitral valve, tricuspid valve, and ventricular septum in fetal and adult C57BL/6J mouse hearts 等人在胎儿和成人C57BL / 6J小鼠心脏中发现了一组位于心房,二尖瓣,三尖瓣和心室间隔的色素细胞。[170]. These cells are positive for tyrosinase-related protein 这些细胞对酪氨酸酶相关蛋白1呈阳性,酪氨酸酶相关蛋白1, a specific marker of melanocytes and their precursors 是黑素细胞及其前体的特定标志物[170]. Subsequent studies have further des随后的研究进一步描述了心脏色素细胞。这些细胞还表达多巴色素互变异构酶(Dcribed cardiac pigment cells. These cells also express dopachrome tautomerase (Dct), which is a recognized marker for melanocyte stem cells and relies on the same signaling pathways involved in normal skin melanocyte development t),Dct是黑色素细胞干细胞的公认标志物,依赖于参与正常皮肤黑素细胞发育的相同信号通路。[171]. These facts indicate that pigmented cardiac cells are melanocytes or melanocyte-related 这些事实表明,色素沉着的心脏细胞是黑素细胞或黑素细胞相关的。[171]. In addition, the cardiac pigmentation levels may be relevant to coat color 此外,心脏色素沉着水平可能与毛色有关[172]. It has been repeatedly proven that melanocytes are located in the valves (mitral, tricuspid, and aortic) and septum (ventricular and atrium) 已经反复证明,黑素细胞位于瓣膜(二尖瓣、三尖瓣和主动脉)和鼻中隔(心室和心房)中[171][173][174]; however, a distinct perspective has recently been put forward 然而,最近提出了一个独特的观点[173], proving the existence of melanocytes in the pulmonary valve and suggesting that the incidence and degree of pigmentation in the heart did not differ statistically with the coat color of the animals.,证明肺动脉瓣中存在黑素细胞,并表明心脏色素沉着的发生率和程度与动物的毛色没有统计学上的差异。 Concentrating on the atrioventricular valve, Balani et al. found that the presence of melanocyte pigmentation may affect the stiffness of the tricuspid valve leaflet of mice 等人专注于房室瓣,发现黑素细胞色素沉着的存在可能会影响小鼠三尖瓣小叶的硬度[174]. The 房室瓣膜的粘弹性与色素沉着的程度相关。此外,支持这一观点的一项新发现表明,房室瓣的黑素细胞主要存在于高度表达viscoelastic property of atrioventricular valves is correlated with the degree of pigmentation. Furthermore, a novel finding supporting this view showed that the melanocytes of the atrioventricular valve mainly exist in the region that highly expresses versican B, a molecule responsible for the mechanical properties of murine atrioventricular valves ersican B的区域,versican B是一种负责小鼠房室瓣膜机械性能的分子。[175]. Additionally, melanocytes may influence other extracellular matrix molecules to contribute to the changes in the valve microenvironment 此外,黑素细胞可能会影响其他细胞外基质分子,以促进瓣膜微环境的变化。[175]. Intriguingly, cardiac melanocytes are considered to trigger atrial arrhythmias 有趣的是,心脏黑素细胞被认为是引发房性心律失常的。[176]. Both mouse and human cardiac melanocytes express the melanin synthase 小鼠和人心脏黑素细胞都表达黑色素合酶Dct, which is involved in buffering calcium and reactive oxygen species. Compared with wild-type mice, mice lacking functional Dct display increased susceptibility to atrial arrhythmias and are proven to produce frequent intracellular calcium oscillations. In contrast, mice lacking melanocytes in the heart fail to show atrial arrhythmia even if they lack DCT as well. Treating Dct-deficient mice with antioxidants can also significantly reduce arrhythmogenesis ,其参与缓冲钙和活性氧。与野生型小鼠相比,缺乏功能性Dct的小鼠表现出对房性心律失常的易感性增加,并被证明产生频繁的细胞内钙振荡。相比之下,心脏中缺乏黑素细胞的小鼠即使缺乏DCT也无法显示房性心律失常。用抗氧化剂治疗缺乏Dct的小鼠也可以显着减少心律失常的发生[176]. In addition, 此外,Hwang et al. showed cardiac melanocytes were located in the anatomical region, including pulmonary veins, atrioventricular valve annulus, left posterior atrium, and oval foramen, where atrial arrhythmia commonly occurs 等人显示心脏黑素细胞位于解剖区域,包括肺静脉,房室瓣环,左后心房和椭圆形孔,其中通常发生房性心律失常[177]. Then they confirmed that 然后他们证实,心脏黑素细胞表达的Dct expressed by cardiac melanocytes is involved with regulating whole atrial oxidative stress which leads to electrical and structural remodeling and triggers the occurrence of atrial arrhythmias in mice 参与调节整个心房氧化应激,导致电和结构重塑,并引发小鼠房性心律失常的发生。[178]. A further study uncovered the underlying mechanisms of the atrial arrhythmia hypothesis 进一步的研究揭示了房性心律失常假说的潜在机制。[179]. It is assumed that 据推测,Dct−deficient mice may have upregulated small conductance calcium-activated potassium current and therefore may be more prone to initiating atrial arrhythmias -缺陷小鼠可能具有上调的小电导钙活化钾电流,因此可能更容易引发房性心律失常[179].5.2. Ear
5.2. 耳朵
Although generally localized in visible anatomical sites, melanocytes also exist as intermediate cells in the stria vascularis of the cochlea in mammals and are essential for proper cochlear development 虽然黑素细胞通常局限于可见的解剖部位,但黑素细胞也作为中间细胞存在于哺乳动物耳蜗血管的纹状体中,并且对于耳蜗的正常发育至关重要。[180]. Melanocytes are also essential for normal auditory function by maintaining the endolymphatic potential at the scala media 黑素细胞对于正常的听觉功能也是必不可少的,因为它可以维持鳞片介质的内淋巴潜力[180]. Additionally, numerous pericytes and perivascular macrophage-like melanocytes (此外,许多周细胞和血管周围巨噬细胞样黑素细胞(PVM/Ms), as specialized melanocytes expressing several melanocyte markers, are known to play vital roles in stabilizing the intrastriatal fluid–blood barrier and sustaining the endocochlear potential, which is critical for normal hearing / Ms)作为表达几种黑素细胞标志物的特化黑素细胞,已知在稳定胸骨内液 - 血液屏障和维持眼内电位方面起着至关重要的作用,这对正常听力至关重要[181]. Moreover, these specialized melanocytes mediate disruption of the blood barrier responding to acoustic trauma, and finally result in the pathological deterioration of ear health after noise damage 此外,这些特化的黑素细胞介导对听觉创伤作出反应的血液屏障的破坏,最终导致噪声损伤后耳朵健康的病理恶化。[182]. Hearing impairment, in some cases, is associated with genetic pigment disorders, such as 在某些情况下,听力障碍与遗传性色素紊乱有关,例如Waardenburg syndrome type 2A (WS2A), a rare inherited disorder with autosomal dominant inheritance 综合征2A型(WS2A),这是一种罕见的常染色体显性遗传的遗传性疾病。[183]. WS2A is caused by the mutation of microphthalmia-associated transcription factor (MITF), an essential regulator for melanocyte lineage development, and is characterized by varying degrees of pigmentation abnormalities and sensorineural hearing impairment 是由小眼症相关转录因子(MITF)突变引起的,MITF是黑素细胞谱系发育的重要调节因子,其特征在于不同程度的色素沉着异常和感音神经性听力障碍[183]. There are other diseases exhibiting deafness or arrhythmias combined with hypopigmentation that further highlight the importance of the melanocytes, such as 还有其他疾病表现出耳聋或心律失常,并伴有色素减退,进一步突出了黑素细胞的重要性,例如Alezzandrini syndrome, Vogt–综合征,Vogt-Koyanagi–-Harada disease, and Tietz syndrome 病和Tietz综合征[184][185][186]. Moreover, observations that bilateral cochlear dysfunction is common in both segmental and nonsegmental vitiligo individuals have illustrated the important role of melanocytes in cochlear function 此外,双侧耳蜗功能障碍在节段性和非节段性白癜风个体中都很常见的观察表明,黑色素细胞在耳蜗功能中的重要作用。[187][188]. Additionally, it is suggested that vitiligo patients should take extra precautions to prevent acoustic trauma and avoid ototoxic drugs, especially during the period of disease activity, as their auditory system may be more vulnerable due to impaired melanocytes 此外,建议白癜风患者应采取额外的预防措施,以防止听觉创伤并避免耳毒性药物,特别是在疾病活动期间,因为他们的听觉系统可能由于黑素细胞受损而更容易受到伤害。[187][188]. Cochlear dysfunction may increase with a longer duration of the disease耳蜗功能障碍可能随着疾病持续时间的延长而增加; therefore, related audiometry tests should be performed for early detection of outer hair cells injury 因此,应进行相关的测听测试,以早期发现外毛细胞损伤[189]. Interestingly, pigmentation is not essential for the hearing ability of the inner ear, as hearing ability is indeed largely unaffected in most albino mice and albino guinea pigs 有趣的是,色素沉着对于内耳的听力能力不是必需的,因为在大多数白化小鼠和白化豚鼠中,听力能力确实在很大程度上不受影响。[180]. Nevertheless, it has been suggested that melanogenesis seems to exert protective effects in response to stressful conditions, including intense noise exposure and ototoxic injury 然而,有人认为黑色素生成似乎对压力条件(包括强烈的噪音暴露和耳毒性损伤)具有保护作用。[180]. The pigmented cochlea shows lower susceptibility to noise, which can be explained by the role of melanin as a scavenger of reactive oxygen species. In addition, the melanin of melanocytes in the inner ear is capable of incorporating drugs, such as aminoglycoside antibiotics and cisplatin, which also explains the potential toxicity to the auditory function of the cochlea 色素沉着的耳蜗对噪声的易感性较低,这可以通过黑色素作为活性氧的清除剂的作用来解释。此外,内耳黑素细胞的黑色素能够掺入药物,如氨基糖苷类抗生素和顺铂,这也解释了对耳蜗听觉功能的潜在毒性。[190][191][192][193].5.3. Other Organs
其他器官
In addition, other extracutaneous pigment cells, such as choroidal melanocytes, have been proven to contribute to visual function by supporting normal morphogenesis and maintaining functional vasculature structures and may not depend on their melanin-producing role 此外,其他皮外色素细胞,如脉络膜黑素细胞,已被证明通过支持正常的形态发生和维持功能性脉管系统结构来促进视觉功能,并且可能不依赖于它们的黑色素产生作用。[194]. There is evidence that the melanocytes in the brain might have neuroendocrine functions 有证据表明,大脑中的黑素细胞可能具有神经内分泌功能。[195]. There is also evidence that the ectopic synthesis of melanin in human adipose tissue has been speculated as a compensatory mechanism for abating oxidative stress and responding to inflammation 还有证据表明,人类脂肪组织中黑色素的异位合成已被推测为减轻氧化应激和应对炎症的代偿机制。[196][197]. The functions of extracutaneous melanocytes located in various regions are summarized in Table 位于不同区域的皮外黑素细胞的功能总结在表2.中。| Location位置 | Function功能 | Reference参考 |
|---|---|---|
| Heart心 | Support the stiffness and mechanical properties of the cardiac valves支持心脏瓣膜的刚度和机械性能 | [172][173][174][175][176] |
| Reduce降低 ROS | ||
| Regulate electrical and structural remodel调节电气和结构改造 | ||
| Maintain the endolymphatic potential维持内淋巴潜力 | ||
| Regulate cochlear development调节人工耳蜗发育 | ||
| Stabilize the intrastriatal fluid–blood barrier稳定纹状体内液体-血液屏障 | ||
| Protect from noise and ototoxic防止噪音和耳毒性 | ||
| Inner ear内耳 | Reduce降低 ROS | [180][181][182][190][191][193] |
| Eye眼睛 | Eye pigmentation and protection against UV眼睛色素沉着和紫外线防护 | [189][194][198][199] |
| Support the normal vasculature of the choroid支持脉络膜的正常脉管系统 | ||
| Induce chemokine secretion and monocyte Migration诱导趋化因子分泌和单核细胞迁移 | ||
| Sebaceous glands皮脂腺 | May be a source of melanocyte stem cells可能是黑素细胞干细胞的来源 | [200] |
| Brain脑 | Neuroendocrine and detoxification神经内分泌和解毒 | [195] |
| Adipose脂肪的 | Abate oxidative stress and inflammation减轻氧化应激和炎症 | [196][197] |
Recent evidence has led to a renewed appreciation for the embryonic origin of cutaneous melanocytes in that these pigment cells not only originate from the migratory neural crest cells but also arise from the nerve-derived multipotent 最近的证据表明,人们对皮肤黑素细胞的胚胎起源有了新的认识,因为这些色素细胞不仅起源于迁移神经嵴细胞,还来自神经来源的多能施旺细胞前体(Schwann cell precursors (SCPs) CP)。[201][202]. Furthermore, this 此外,这种皮肤黑素细胞的SCP-dependent origin of skin melanocytes has been conserved among fish, birds, and mammals during evolution 依赖性起源在进化过程中在鱼类,鸟类和哺乳动物中得到了保护。[201][202][203][204]. Still, the origin of extracutaneous melanocytes located in the inner ear, brain meninges, heart, and other locations has been an 尽管如此,位于内耳,脑膜,心脏和其他部位的皮外黑素细胞的起源一直是一个“enigmatic” problem, fascinating scientists. Recently, this long-standing problem has been gradually addressed by a lineage-tracing strategy combined with 3D visualization, revealing that peripheral nerve-derived SCPs are an essential cellular origin of extracutaneous melanocytes 神秘”的问题,迷人的科学家。最近,通过谱系追踪策略与3D可视化相结合,逐渐解决了这一长期存在的问题,揭示了周围神经衍生的SCP是皮外黑素细胞的基本细胞起源。[204][205]. The origin of such extracutaneous melanocytes can be associated with their localized specialization and their unconventional roles. But this complicates researchers' understanding of what melanocytes really do 这种皮外黑素细胞的起源可能与它们的局部特化及其非常规作用有关。但这使我们对黑素细胞真正作用的理解复杂化。[204]. Therefore, melanocytes outside the skin also exhibit fascinating functions other than melanin synthesis, and further exploration of the embryonic development of melanocytes will lead to a better understanding of the specific skin disorders and envisioning future prevention and treatment strategies 因此,皮肤外的黑素细胞也表现出黑色素合成以外的迷人功能,进一步探索黑色素细胞的胚胎发育将导致对特定皮肤疾病的更好理解,并设想未来的预防和治疗策略。[204].
6. Conclusions
The last several decades have seen tremendous advances in ouresearchers' understanding of the biology of melanocytes. ResearchersWe now adequately recognize that melanocytes are amazing cells and should not be solely considered as a “melanin factory.” They can carry out multiple functions, acting as epidermal photoreceptors and an active partner of the skin’s immune and neuroendocrine systems and responding quickly and effectively to environmental information to regulate the local and systemic homeostasis, as shown in Figure 2. However, critical questions as to why melanocytes are equipped with so many functional devices and to what extent these physiological functions can be exerted and even whether there is coordination between these functions, such as neuroendocrine activities and circadian rhythms, should be further integrated. Despite how these activities affect local physiology, how they transmit and amplify signals and even integrate with other remote tissues remains relatively unknown. Undoubtedly, the numerous activities of melanocytes are complex and highly controlled processes. Therefore, deciphering the mechanisms of the communication process can potentially contribute to gaining insight into the pleiotropic roles of melanocytes within the organism and can open a completely new horizon for revolutionizing the way diseases will be diagnosed and treated in the future.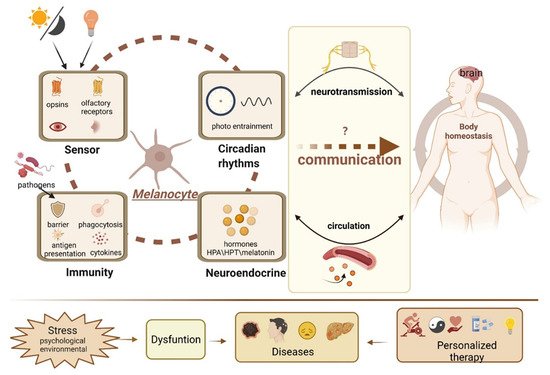
References
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850.
- Sulaimon, S.S.; Kitchell, B.E. The biology of melanocytes. Vet. Dermatol. 2003, 14, 57–65.
- Cordero, R.J.B.; Casadevall, A. Melanin. Curr. Biol. 2020, 30, R142–R143.
- ElObeid, A.S.; Kamal-Eldin, A.; Abdelhalim, M.A.K.; Haseeb, A.M. Pharmacological Properties of Melanin and its Function in Health. Basic Clin. Pharmacol. Toxicol. 2017, 120, 515–522.
- Plonka, P.M.; Passeron, T.; Brenner, M.; Tobin, D.J.; Shibahara, S.; Thomas, A.; Slominski, A.; Kadekaro, A.L.; Hershkovitz, D.; Peters, E.; et al. What are melanocytes really doing all day long…? Exp. Dermatol. 2009, 18, 799–819.
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115.
- Slominski, A.; Paus, R.; Schadendorf, D. Melanocytes as “sensory” and regulatory cells in the epidermis. J. Theor. Biol. 1993, 164, 103–120.
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tyminska, A. Skin melanocytes: Biology and development. Postepy Dermatol. Alergol. 2013, 30, 30–41.
- Slominski, A. Neuroendocrine activity of the melanocyte. Exp. Dermatol. 2009, 18, 760–763.
- Alexopoulos, A.; Chrousos, G.P. Stress-related skin disorders. Rev. Endocr. Metab. Disord. 2016, 17, 295–304.
- Phan, T.S.; Schink, L.; Mann, J.; Merk, V.M.; Zwicky, P.; Mundt, S.; Simon, D.; Kulms, D.; Abraham, S.; Legler, D.F.; et al. Keratinocytes control skin immune homeostasis through de novo-synthesized glucocorticoids. Sci. Adv. 2021, 7, eabe0337.
- Paus, R.; Arck, P. Neuroendocrine perspectives in alopecia areata: Does stress play a role? J. Investig. Dermatol. 2009, 129, 1324–1326.
- Kotb El-Sayed, M.I.; Abd El-Ghany, A.A.; Mohamed, R.R. Neural and Endocrinal Pathobiochemistry of Vitiligo: Comparative Study for a Hypothesized Mechanism. Front. Endocrinol. 2018, 9, 197.
- Zhang, B.; Ma, S.; Rachmin, I.; He, M.; Baral, P.; Choi, S.; Goncalves, W.A.; Shwartz, Y.; Fast, E.M.; Su, Y.; et al. Hyperactivation of sympathetic nerves drives depletion of melanocyte stem cells. Nature 2020, 577, 676–681.
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine Aspects of Skin Aging. Int. J. Mol. Sci. 2019, 20, 2798.
- Theoharides, T.C.; Stewart, J.M.; Taracanova, A.; Conti, P.; Zouboulis, C.C. Neuroendocrinology of the skin. Rev. Endocr. Metab. Disord. 2016, 17, 287–294.
- Leis, K.; Mazur, E.; Jablonska, M.J.; Kolan, M.; Galazka, P. Endocrine systems of the skin. Postepy Dermatol. Alergol. 2019, 36, 519–523.
- Takeda, K.; Takahashi, N.H.; Shibahara, S. Neuroendocrine functions of melanocytes: Beyond the skin-deep melanin maker. Tohoku J. Exp. Med. 2007, 211, 201–221.
- Rousseau, K.; Kauser, S.; Pritchard, L.E.; Warhurst, A.; Oliver, R.L.; Slominski, A.; Wei, E.T.; Thody, A.J.; Tobin, D.J.; White, A. Proopiomelanocortin (POMC), the ACTH/melanocortin precursor, is secreted by human epidermal keratinocytes and melanocytes and stimulates melanogenesis. FASEB J. 2007, 21, 1844–1856.
- Spencer, J.D.; Schallreuter, K.U. Regulation of pigmentation in human epidermal melanocytes by functional high-affinity beta-melanocyte-stimulating hormone/melanocortin-4 receptor signaling. Endocrinology 2009, 150, 1250–1258.
- Slominski, A. Identification of beta-endorphin, alpha-MSH and ACTH peptides in cultured human melanocytes, melanoma and squamous cell carcinoma cells by RP-HPLC. Exp. Dermatol. 1998, 7, 213–216.
- Bhm, M.; Metze, D.; Schulte, U.; Becher, E.; Luger, T.A.; Brzoska, T. Detection of melanocortin-1 receptor antigenicity on human skin cells in culture and in situ. Exp. Dermatol. 2010, 8, 453–461.
- Farooqui, J.Z.; Medrano, E.E.; Abdel-Malek, Z.; Nordlund, J. The expression of proopiomelanocortin and various POMC-derived peptides in mouse and human skin. Ann. N. Y. Acad. Sci. 1993, 680, 508–510.
- Slominski, A.; Ermak, G.; Hwang, J.; Chakraborty, A.; Mazurkiewicz, J.E.; Mihm, M. Proopiomelanocortin, corticotropin releasing hormone and corticotropin releasing hormone receptor genes are expressed in human skin. FEBS Lett. 1995, 374, 113–116.
- Slominski, A.; Baker, J.; Ermak, G.; Chakraborty, A.; Pawelek, J. Ultraviolet B stimulates production of corticotropin releasing factor (CRF) by human melanocytes. FEBS Lett. 1996, 399, 175–176.
- Kono, M.; Nagata, H.; Umemura, S.; Kawana, S.; Osamura, R.Y. In situ expression of corticotropin-releasing hormone (CRH) and proopiomelanocortin (POMC) genes in human skin. FASEB J. 2001, 15, 2297–2299.
- Slominski, A.; Zbytek, B.; Szczesniewski, A.; Semak, I.; Kaminski, J.; Sweatman, T.; Wortsman, J. CRH stimulation of corticosteroids production in melanocytes is mediated by ACTH. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E701–E706.
- Ramot, Y.; Bohm, M.; Paus, R. Translational Neuroendocrinology of Human Skin: Concepts and Perspectives. Trends Mol. Med. 2021, 27, 60–74.
- Skobowiat, C.; Dowdy, J.C.; Sayre, R.M.; Tuckey, R.C.; Slominski, A. Cutaneous hypothalamic-pituitary-adrenal axis homolog: Regulation by ultraviolet radiation. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E484–E493.
- Skobowiat, C.; Nejati, R.; Lu, L.; Williams, R.W.; Slominski, A.T. Genetic variation of the cutaneous HPA axis: An analysis of UVB-induced differential responses. Gene 2013, 530, 1–7.
- Slominski, A.T.; Slominski, R.M.; Raman, C. UVB stimulates production of enkephalins and other neuropeptides by skin-resident cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2020425118.
- Jozic, I.; Stojadinovic, O.; Kirsner, R.S.F.; Tomic-Canic, M. Skin under the (Spot)-Light: Cross-Talk with the Central Hypothalamic-Pituitary-Adrenal (HPA) Axis. J. Investig. Dermatol. 2015, 135, 1469–1471.
- Sato, H.; Nagashima, Y.; Chrousos, G.P.; Ichihashi, M.; Funasak, Y. The expression of corticotropin-releasing hormone in melanoma. Pigment. Cell Res. 2002, 15, 98–103.
- Kim, M.H.; Cho, D.; Kim, H.J.; Chong, S.J.; Lee, K.H.; Yu, D.S.; Park, C.J.; Lee, J.Y.; Cho, B.K.; Park, H.J. Investigation of the corticotropin-releasing hormone-proopiomelanocortin axis in various skin tumours. Br. J. Dermatol. 2006, 155, 910–915.
- Eberle, A.N.; Rout, B.; Qi, M.B.; Bigliardi, P.L. Synthetic Peptide Drugs for Targeting Skin Cancer: Malignant Melanoma and Melanotic Lesions. Curr. Med. Chem. 2017, 24, 1797–1826.
- Kingo, K.; Aunin, E.; Karelson, M.; Philips, M.A.; Ratsep, R.; Silm, H.; Vasar, E.; Soomets, U.; Koks, S. Gene expression analysis of melanocortin system in vitiligo. J. Dermatol. Sci. 2007, 48, 113–122.
- Shaker, O.G.; Eltahlawi, S.M.; Tawfic, S.O.; Eltawdy, A.M.; Bedair, N.I. Corticotropin-releasing hormone (CRH) and CRH receptor 1 gene expression in vitiligo. Clin. Exp. Dermatol. 2016, 41, 734–740.
- Spencer, J.; Gibbons, N.; Rokos, H.; Peters, E.; Wood, J.; Schallreuter, K. Oxidative stress via hydrogen peroxide affects proopiomelanocortin peptides directly in the epidermis of patients with vitiligo. J. Investig. Dermatol. 2007, 127, 411–420.
- Mancino, G.; Miro, C.; Di Cicco, E.; Dentice, M. Thyroid hormone action in epidermal development and homeostasis and its implications in the pathophysiology of the skin. J. Endocrinol. Investig. 2021, 44, 1571–1579.
- Bodo, E.; Kany, B.; Gaspar, E.; Knuver, J.; Kromminga, A.; Ramot, Y.; Biro, T.; Tiede, S.; van Beek, N.; Poeggeler, B.; et al. Thyroid-stimulating hormone, a novel, locally produced modulator of human epidermal functions, is regulated by thyrotropin-releasing hormone and thyroid hormones. Endocrinology 2010, 151, 1633–1642.
- Baldini, E.; Odorisio, T.; Sorrenti, S.; Catania, A.; Tartaglia, F.; Carbotta, G.; Pironi, D.; Rendina, R.; D’Armiento, E.; Persechino, S.; et al. Vitiligo and Autoimmune Thyroid Disorders. Front. Endocrinol. 2017, 8, 290.
- Slominski, A.; Wortsman, J.; Kohn, L.; Ain, K.B.; Venkataraman, G.M.; Pisarchik, A.; Chung, J.H.; Giuliani, C.; Thornton, M.; Slugocki, G.; et al. Expression of hypothalamic-pituitary-thyroid axis related genes in the human skin. J. Investig. Dermatol. 2002, 119, 1449–1455.
- Gaspar, E.; Nguyen-Thi, K.T.; Hardenbicker, C.; Tiede, S.; Plate, C.; Bodo, E.; Knuever, J.; Funk, W.; Biro, T.; Paus, R. Thyrotropin-releasing hormone selectively stimulates human hair follicle pigmentation. J. Investig. Dermatol. 2011, 131, 2368–2377.
- Sandru, F.; Carsote, M.; Albu, S.E.; Dumitrascu, M.C.; Valea, A. Vitiligo and chronic autoimmune thyroiditis. J. Med. Life 2021, 14, 127–130.
- Liu, M.; Murphy, E.; Amerson, E.H. Rethinking screening for thyroid autoimmunity in vitiligo. J. Am. Acad. Dermatol. 2016, 75, 1278–1280.
- Li, D.; Liang, G.; Calderone, R.; Bellanti, J.A. Vitiligo and Hashimoto’s thyroiditis: Autoimmune diseases linked by clinical presentation, biochemical commonality, and autoimmune/oxidative stress-mediated toxicity pathogenesis. Med. Hypotheses 2019, 128, 69–75.
- Ellerhorst, J.A.; Naderi, A.A.; Johnson, M.K.; Pelletier, P.; Prieto, V.G.; Diwan, A.H.; Johnson, M.M.; Gunn, D.C.; Yekell, S.; Grimm, E.A. Expression of thyrotropin-releasing hormone by human melanoma and nevi. Clin. Cancer Res. 2004, 10, 5531–5536.
- Ellerhorst, J.A.; Sendi-Naderi, A.; Johnson, M.K.; Cooke, C.P.; Dang, S.M.; Diwan, A.H. Human melanoma cells express functional receptors for thyroid-stimulating hormone. Endocr. Relat. Cancer 2006, 13, 1269–1277.
- Ursu, H. Functional TSH Receptors, Malignant Melanomas and Subclinical Hypothyroidism. Eur. Thyroid. J. 2012, 1, 208.
- Hou, P.; Liu, D.; Ji, M.; Liu, Z.; Engles, J.M.; Wahl, R.L.; Xing, M. Induction of thyroid gene expression and radioiodine uptake in melanoma cells: Novel therapeutic implications. PLoS ONE 2009, 4, e6200.
- Scheau, C.; Draghici, C.; Ilie, M.A.; Lupu, M.; Caruntu, C. Neuroendocrine Factors in Melanoma Pathogenesis. Cancers 2021, 13, 2277.
- Sevilla, A.; Chéret, J.; Slominski, R.M.; Slominski, A.T.; Paus, R. Revisiting the role of melatonin in human melanocyte physiology: A skin context perspective. J. Pineal Res. 2022, 72, e12790.
- Rusanova, I.; Martinez-Ruiz, L.; Florido, J.; Rodriguez-Santana, C.; Guerra-Librero, A.; Acuna-Castroviejo, D.; Escames, G. Protective Effects of Melatonin on the Skin: Future Perspectives. Int. J. Mol. Sci. 2019, 20, 4948.
- Kim, T.K.; Kleszczynski, K.; Janjetovic, Z.; Sweatman, T.; Lin, Z.; Li, W.; Reiter, R.J.; Fischer, T.W.; Slominski, A.T. Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells. FASEB J. 2013, 27, 2742–2755.
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Wortsman, J.; Szczesniewski, A.; Slugocki, G.; McNulty, J.; Kauser, S.; Tobin, D.J.; et al. Serotoninergic and melatoninergic systems are fully expressed in human skin. FASEB J. 2002, 16, 896–898.
- Slominski, A.T.; Kim, T.K.; Kleszczynski, K.; Semak, I.; Janjetovic, Z.; Sweatman, T.; Skobowiat, C.; Steketee, J.D.; Lin, Z.; Postlethwaite, A.; et al. Characterization of serotonin and N-acetylserotonin systems in the human epidermis and skin cells. J. Pineal Res. 2020, 68, e12626.
- Slominski, A.; Pisarchik, A.; Johansson, O.; Jing, C.; Semak, I.; Slugocki, G.; Wortsman, J. Tryptophan hydroxylase expression in human skin cells. Biochim. Biophys. Acta—Mol. Basis Dis. 2003, 1639, 80–86.
- Kim, T.K.; Lin, Z.; Tidwell, W.J.; Li, W.; Slominski, A.T. Melatonin and its metabolites accumulate in the human epidermis in vivo and inhibit proliferation and tyrosinase activity in epidermal melanocytes in vitro. Mol. Cell Endocrinol. 2015, 404, 1–8.
- Slominski, A.; Semak, I.; Pisarchik, A.; Sweatman, T.; Szczesniewski, A.; Wortsman, J. Conversion ofL-tryptophan to serotonin and melatonin in human melanoma cells. FEBS Lett. 2002, 511, 102–106.
- Skobowiat, C.; Brozyna, A.A.; Janjetovic, Z.; Jeayeng, S.; Oak, A.S.W.; Kim, T.K.; Panich, U.; Reiter, R.J.; Slominski, A.T. Melatonin and its derivatives counteract the ultraviolet B radiation-induced damage in human and porcine skin ex vivo. J. Pineal Res. 2018, 65, e12501.
- Kleszczynski, K.; Kim, T.K.; Bilska, B.; Sarna, M.; Mokrzynski, K.; Stegemann, A.; Pyza, E.; Reiter, R.J.; Steinbrink, K.; Bohm, M.; et al. Melatonin exerts oncostatic capacity and decreases melanogenesis in human MNT-1 melanoma cells. J. Pineal Res. 2019, 67, e12610.
- Perdomo, J.; Quintana, C.; Gonzalez, I.; Hernandez, I.; Rubio, S.; Loro, J.F.; Reiter, R.J.; Estevez, F.; Quintana, J. Melatonin Induces Melanogenesis in Human SK-MEL-1 Melanoma Cells Involving Glycogen Synthase Kinase-3 and Reactive Oxygen Species. Int. J. Mol. Sci. 2020, 21, 4970.
- Janjetovic, Z.; Nahmias, Z.P.; Hanna, S.; Jarrett, S.G.; Kim, T.K.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites ameliorate ultraviolet B-induced damage in human epidermal keratinocytes. J. Pineal Res. 2014, 57, 90–102.
- Janjetovic, Z.; Jarrett, S.G.; Lee, E.F.; Duprey, C.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites protect human melanocytes against UVB-induced damage: Involvement of NRF2-mediated pathways. Sci Rep. 2017, 7, 1274.
- Dong, K.; Goyarts, E.; Rella, A.; Pelle, E.; Wong, Y.H.; Pernodet, N. Age Associated Decrease of MT-1 Melatonin Receptor in Human Dermal Skin Fibroblasts Impairs Protection Against UV-Induced DNA Damage. Int. J. Mol. Sci. 2020, 21, 326.
- Gillbro, J.M.; Marles, L.K.; Hibberts, N.A.; Schallreuter, K.U. Autocrine catecholamine biosynthesis and the beta-adrenoceptor signal promote pigmentation in human epidermal melanocytes. J. Investig. Dermatol. 2004, 123, 346–353.
- Sivamani, R.K.; Porter, S.M.; Isseroff, R.R. An epinephrine-dependent mechanism for the control of UV-induced pigmentation. J. Investig. Dermatol. 2009, 129, 784–787.
- Choi, M.E.; Yoo, H.; Lee, H.R.; Moon, I.J.; Lee, W.J.; Song, Y.; Chang, S.E. Carvedilol, an Adrenergic Blocker, Suppresses Melanin Synthesis by Inhibiting the cAMP/CREB Signaling Pathway in Human Melanocytes and Ex Vivo Human Skin Culture. Int. J. Mol. Sci. 2020, 21, 8796.
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators of melanocyte functions. Pigment. Cell Melanoma Res. 2012, 25, 14–27.
- Ono, K.; Viet, C.T.; Ye, Y.; Dang, D.; Hitomi, S.; Toyono, T.; Inenaga, K.; Dolan, J.C.; Schmidt, B.L. Cutaneous pigmentation modulates skin sensitivity via tyrosinase-dependent dopaminergic signalling. Sci. Rep. 2017, 7, 9181.
- Mackintosh, J.A. The antimicrobial properties of melanocytes, melanosomes and melanin and the evolution of black skin. J. Theor. Biol. 2001, 211, 101–113.
- Gasque, P.; Jaffar-Bandjee, M.C. The immunology and inflammatory responses of human melanocytes in infectious diseases. J. Infect. 2015, 71, 413–421.
- Quaresma, J. Organization of the Skin Immune System and Compartmentalized Immune Responses in Infectious Diseases. Clin. Microbiol. Rev. 2019, 32, e00034-18.
- Harder, J.; Schroder, J.M.; Glaser, R. The skin surface as antimicrobial barrier: Present concepts and future outlooks. Exp. Dermatol. 2013, 22, 1–5.
- Wang, S.; Liu, D.; Ning, W.; Xu, A. Cytosolic dsDNA triggers apoptosis and pro-inflammatory cytokine production in normal human melanocytes. Exp. Dermatol. 2015, 24, 298–300.
- Iverson, M.V. Hypothesis: Vitiligo virus. Pigment Cell Res. 2000, 13, 281–282.
- Erf, G.F.; Bersi, T.K.; Wang, X.; Sreekumar, G.P.; Smyth, J.R., Jr. Herpesvirus connection in the expression of autoimmune vitiligo in Smyth line chickens. Pigment Cell Res. 2001, 14, 40–46.
- Kawamura, T.; Ogawa, Y.; Aoki, R.; Shimada, S. Innate and intrinsic antiviral immunity in skin. J. Dermatol. Sci. 2014, 75, 159–166.
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30.
- Coates, M.; Blanchard, S.; MacLeod, A.S. Innate antimicrobial immunity in the skin: A protective barrier against bacteria, viruses, and fungi. PLoS Pathog. 2018, 14, e1007353.
- Ahn, J.H.; Park, T.J.; Jin, S.H.; Kang, H.Y. Human melanocytes express functional Toll-like receptor 4. Exp. Dermatol. 2008, 17, 412–417.
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384.
- Tapia, C.; Falconer, M.; Tempio, F.; Falcón, F.; López, M.; Fuentes, M.; Alburquenque, C.; Amaro, J.; Bucarey, S.; Di Nardo, A. Melanocytes and melanin represent a first line of innate immunity against Candida albicans. Med. Mycol. 2014, 52, 445–454.
- Tam, I.; Dzierzega-Lecznar, A.; Stepien, K. Differential expression of inflammatory cytokines and chemokines in lipopolysaccharide-stimulated melanocytes from lightly and darkly pigmented skin. Exp. Dermatol. 2019, 28, 551–560.
- Tam, I.; Stępień, K. Secretion of proinflammatory cytokines by normal human melanocytes in response to lipopolysaccharide. Acta Biochim. Pol. 2011, 58, 507–511.
- Song, H.; Lee, S.; Choi, G.; Shin, J. Repeated ultraviolet irradiation induces the expression of Toll-like receptor 4, IL-6, and IL-10 in neonatal human melanocytes. Photodermatol. Photoimmunol. Photomed. 2018, 34, 145–151.
- Koike, S.; Yamasaki, K. Melanogenesis Connection with Innate Immunity and Toll-Like Receptors. Int. J. Mol. Sci. 2020, 21, 9769.
- Vavricka, C.J.; Christensen, B.M.; Li, J. Melanization in living organisms: A perspective of species evolution. Protein Cell 2010, 1, 830–841.
- D’Alba, L.; Shawkey, M.D. Melanosomes: Biogenesis, Properties, and Evolution of an Ancient Organelle. Physiol. Rev. 2019, 99, 1–19.
- Tolleson, W.H. Human melanocyte biology, toxicology, and pathology. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2005, 23, 105–161.
- Montefiori, D.C.; Zhou, J. Selective antiviral activity of synthetic soluble l-tyrosine and l-dopa melanins against human immunodeficiency virus in vitro. Antivir. Res. 1991, 15, 11–25.
- Ali, S.M.; Yosipovitch, G. Skin pH: From basic science to basic skin care. Acta Derm. Venereol. 2013, 93, 261–267.
- Gunathilake, R.; Schurer, N.Y.; Shoo, B.A.; Celli, A.; Hachem, J.P.; Crumrine, D.; Sirimanna, G.; Feingold, K.R.; Mauro, T.M.; Elias, P.M. pH-regulated mechanisms account for pigment-type differences in epidermal barrier function. J. Investig. Dermatol. 2009, 129, 1719–1729.
- Elias, P.M.; Menon, G.; Wetzel, B.K.; Williams, J. Barrier requirements as the evolutionary “driver” of epidermal pigmentation in humans. Am. J. Hum. Biol. 2010, 22, 526–537.
- Lin, T.K.; Man, M.Q.; Abuabara, K.; Wakefield, J.S.; Sheu, H.M.; Tsai, J.C.; Lee, C.H.; Elias, P.M. By protecting against cutaneous inflammation, epidermal pigmentation provided an additional advantage for ancestral humans. Evol. Appl. 2019, 12, 1960–1970.
- Jablonski, N.G.; Chaplin, G. Colloquium paper: Human skin pigmentation as an adaptation to UV radiation. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. S2), 8962–8968.
- Man, M.Q.; Lin, T.K.; Santiago, J.L.; Celli, A.; Zhong, L.; Huang, Z.M.; Roelandt, T.; Hupe, M.; Sundberg, J.P.; Silva, K.A.; et al. Basis for enhanced barrier function of pigmented skin. J. Investig. Dermatol. 2014, 134, 2399–2407.
- Liu, J.; Man, W.Y.; Lv, C.Z.; Song, S.P.; Shi, Y.J.; Elias, P.M.; Man, M.Q. Epidermal permeability barrier recovery is delayed in vitiligo-involved sites. Skin Pharmacol. Physiol. 2010, 23, 193–200.
- Koike, S.; Yamasaki, K.; Yamauchi, T.; Inoue, M.; Shimada-Ohmori, R.; Tsuchiyama, K.; Aiba, S. Toll-like receptors 2 and 3 enhance melanogenesis and melanosome transport in human melanocytes. Pigment Cell Melanoma Res. 2018, 31, 570–584.
- Sun, L.; Pan, S.; Yang, Y.; Sun, J.; Liang, D.; Wang, X.; Xie, X.; Hu, J. Toll-like receptor 9 regulates melanogenesis through NF-κB activation. Exp. Biol. Med. 2016, 241, 1497–1504.
- Le Poole, I.C.; van den Wijngaard, R.M.; Westerhof, W.; Verkruisen, R.P.; Dutrieux, R.P.; Dingemans, K.P.; Das, P.K. Phagocytosis by normal human melanocytes in vitro. Exp. Cell Res. 1993, 205, 388–395.
- Smit, N.; Le Poole, I.; van den Wijngaard, R.; Tigges, A.; Westerhof, W.; Das, P. Expression of different immunological markers by cultured human melanocytes. Arch. Dermatol. Res. 1993, 285, 356–365.
- Le Poole, I.; Mutis, T.; van den Wijngaard, R.; Westerhof, W.; Ottenhoff, T.; de Vries, R.; Das, P. A novel, antigen-presenting function of melanocytes and its possible relationship to hypopigmentary disorders. J. Immunol. 1993, 151, 7284–7292.
- Lu, Y.; Zhu, W.Y.; Tan, C.; Yu, G.H.; Gu, J.X. Melanocytes are potential immunocompetent cells: Evidence from recognition of immunological characteristics of cultured human melanocytes. Pigment Cell Res. 2002, 15, 454–460.
- Dalesio, N.M.; Barreto Ortiz, S.F.; Pluznick, J.L.; Berkowitz, D.E. Olfactory, Taste, and Photo Sensory Receptors in Non-sensory Organs: It Just Makes Sense. Front. Physiol. 2018, 9, 1673.
- Moraes, M.N.; de Assis, L.V.M.; Provencio, I.; Castrucci, A.M.L. Opsins outside the eye and the skin: A more complex scenario than originally thought for a classical light sensor. Cell Tissue Res. 2021, 385, 519–538.
- Do, M.T.H. Melanopsin and the Intrinsically Photosensitive Retinal Ganglion Cells: Biophysics to Behavior. Neuron 2019, 104, 205–226.
- Lee, S.J.; Depoortere, I.; Hatt, H. Therapeutic potential of ectopic olfactory and taste receptors. Nat. Rev. Drug Discov. 2019, 18, 116–138.
- Cheret, J.; Bertolini, M.; Ponce, L.; Lehmann, J.; Tsai, T.; Alam, M.; Hatt, H.; Paus, R. Olfactory receptor OR2AT4 regulates human hair growth. Nat. Commun. 2018, 9, 3624.
- Buscone, S.; Mardaryev, A.N.; Raafs, B.; Bikker, J.W.; Sticht, C.; Gretz, N.; Farjo, N.; Uzunbajakava, N.E.; Botchkareva, N.V. A new path in defining light parameters for hair growth: Discovery and modulation of photoreceptors in human hair follicle. Lasers Surg. Med. 2017, 49, 705–718.
- Mignon, C.; Botchkareva, N.V.; Uzunbajakava, N.E.; Tobin, D.J. Photobiomodulation devices for hair regrowth and wound healing: A therapy full of promise but a literature full of confusion. Exp. Dermatol. 2016, 25, 745–749.
- Castellano-Pellicena, I.; Uzunbajakava, N.E.; Mignon, C.; Raafs, B.; Botchkarev, V.A.; Thornton, M.J. Does blue light restore human epidermal barrier function via activation of Opsin during cutaneous wound healing? Lasers Surg. Med. 2019, 51, 370–382.
- Portillo, M.; Mataix, M.; Alonso-Juarranz, M.; Lorrio, S.; Villalba, M.; Rodriguez-Luna, A.; Gonzalez, S. The Aqueous Extract of Polypodium leucotomos (Fernblock((R))) Regulates Opsin 3 and Prevents Photooxidation of Melanin Precursors on Skin Cells Exposed to Blue Light Emitted from Digital Devices. Antioxidants 2021, 10, 400.
- Lan, Y.; Wang, Y.; Lu, H. Opsin 3 is a key regulator of ultraviolet A-induced photoageing in human dermal fibroblast cells. Br. J. Dermatol. 2020, 182, 1228–1244.
- Lan, Y.; Zeng, W.; Dong, X.; Lu, H. Opsin 5 is a key regulator of ultraviolet radiation-induced melanogenesis in human epidermal melanocytes. Br. J. Dermatol. 2021, 185, 391–404.
- Massberg, D.; Hatt, H. Human Olfactory Receptors: Novel Cellular Functions Outside of the Nose. Physiol. Rev. 2018, 98, 1739–1763.
- Suh, S.; Choi, E.H.; Atanaskova Mesinkovska, N. The expression of opsins in the human skin and its implications for photobiomodulation: A Systematic Review. Photodermatol. Photoimmunol. Photomed. 2020, 36, 329–338.
- Leung, N.Y.; Montell, C. Unconventional Roles of Opsins. Annu. Rev. Cell Dev. Biol. 2017, 33, 241–264.
- Buhr, E.D.; Yue, W.W.; Ren, X.; Jiang, Z.; Liao, H.W.; Mei, X.; Vemaraju, S.; Nguyen, M.T.; Reed, R.R.; Lang, R.A.; et al. Neuropsin (OPN5)-mediated photoentrainment of local circadian oscillators in mammalian retina and cornea. Proc. Natl. Acad. Sci. USA 2015, 112, 13093–13098.
- Olinski, L.E.; Lin, E.M.; Oancea, E. Illuminating insights into opsin 3 function in the skin. Adv. Biol. Regul. 2020, 75, 100668.
- Provencio, I.; Jiang, G.; De Grip, W.J.; Hayes, W.P.; Rollag, M.D. Melanopsin: An opsin in melanophores, brain, and eye. Proc. Natl. Acad. Sci. USA 1998, 95, 340–345.
- Isoldi, M.C.; Rollag, M.D.; Castrucci, A.M.; Provencio, I. Rhabdomeric phototransduction initiated by the vertebrate photopigment melanopsin. Proc. Natl. Acad. Sci. USA 2005, 102, 1217–1221.
- Bertolesi, G.E.; McFarlane, S. Seeing the light to change colour: An evolutionary perspective on the role of melanopsin in neuroendocrine circuits regulating light-mediated skin pigmentation. Pigment Cell Melanoma Res. 2018, 31, 354–373.
- Kusumoto, J.; Takeo, M.; Hashikawa, K.; Komori, T.; Tsuji, T.; Terashi, H.; Sakakibara, S. OPN4 belongs to the photosensitive system of the human skin. Genes Cells 2020, 25, 215–225.
- Miyashita, Y.; Moriya, T.; Kubota, T.; Yamada, K.; Asami, K. Expression of opsin molecule in cultured murine melanocyte. J. Investig. Dermatol. Symp. Proc. 2001, 6, 54–57.
- Denda, M.; Fuziwara, S. Visible radiation affects epidermal permeability barrier recovery: Selective effects of red and blue light. J. Investig. Dermatol. 2008, 128, 1335–1336.
- Tsutsumi, M.; Ikeyama, K.; Denda, S.; Nakanishi, J.; Fuziwara, S.; Aoki, H.; Denda, M. Expressions of rod and cone photoreceptor-like proteins in human epidermis. Exp. Dermatol. 2009, 18, 567–570.
- Wicks, N.L.; Chan, J.W.; Najera, J.A.; Ciriello, J.M.; Oancea, E. UVA phototransduction drives early melanin synthesis in human melanocytes. Curr. Biol. 2011, 21, 1906–1911.
- Haltaufderhyde, K.; Ozdeslik, R.N.; Wicks, N.L.; Najera, J.A.; Oancea, E. Opsin expression in human epidermal skin. Photochem. Photobiol. 2015, 91, 117–123.
- Hu, Q.M.; Yi, W.J.; Su, M.Y.; Jiang, S.; Xu, S.Z.; Lei, T.C. Induction of retinal-dependent calcium influx in human melanocytes by UVA or UVB radiation contributes to the stimulation of melanosome transfer. Cell Prolif. 2017, 50, e12372.
- Regazzetti, C.; Sormani, L.; Debayle, D.; Bernerd, F.; Tulic, M.K.; De Donatis, G.M.; Chignon-Sicard, B.; Rocchi, S.; Passeron, T. Melanocytes Sense Blue Light and Regulate Pigmentation through Opsin-3. J. Investig. Dermatol. 2018, 138, 171–178.
- Ozdeslik, R.N.; Olinski, L.E.; Trieu, M.M.; Oprian, D.D.; Oancea, E. Human nonvisual opsin 3 regulates pigmentation of epidermal melanocytes through functional interaction with melanocortin 1 receptor. Proc. Natl. Acad. Sci. USA 2019, 116, 11508–11517.
- Wang, Y.; Lan, Y.; Lu, H. Opsin3 Downregulation Induces Apoptosis of Human Epidermal Melanocytes via Mitochondrial Pathway. Photochem. Photobiol. 2020, 96, 83–93.
- de Assis, L.V.M.; Moraes, M.N.; Magalhaes-Marques, K.K.; Castrucci, A.M.L. Melanopsin and rhodopsin mediate UVA-induced immediate pigment darkening: Unravelling the photosensitive system of the skin. Eur. J. Cell Biol. 2018, 97, 150–162.
- Jin, H.; Zou, Z.; Chang, H.; Shen, Q.; Liu, L.; Xing, D. Photobiomodulation therapy for hair regeneration: A synergetic activation of β-CATENIN in hair follicle stem cells by ROS and paracrine WNTs. Stem Cell Rep. 2021, 16, 1568–1583.
- Le Duff, F.; Fontas, E.; Guardoli, D.; Lacour, J.; Passeron, T. HeaLED: Assessment of skin healing under light-emitting diode (LED) exposure-A randomized controlled study versus placebo. Lasers Surg. Med. 2022, 54, 342–347.
- Diogo, M.; Campos, T.; Fonseca, E.; Pavani, C.; Horliana, A.; Fernandes, K.; Bussadori, S.; Fantin, F.; Leite, D.; Yamamoto, Â.; et al. Effect of Blue Light on Acne Vulgaris: A Systematic Review. Sensors 2021, 21, 6943.
- Spinella, A.; de Pinto, M.; Galluzzo, C.; Testoni, S.; Macripò, P.; Lumetti, F.; Parenti, L.; Magnani, L.; Sandri, G.; Bajocchi, G.; et al. Photobiomodulation Therapy: A New Light in the Treatment of Systemic Sclerosis Skin Ulcers. Rheumatol. Ther. 2022, 9, 891–905.
- Kemény, L.; Varga, E.; Novak, Z. Advances in phototherapy for psoriasis and atopic dermatitis. Expert Rev. Clin. Immunol. 2019, 15, 1205–1214.
- Allada, R.; Bass, J. Circadian Mechanisms in Medicine. N. Engl. J. Med. 2021, 384, 550–561.
- Tanioka, M.; Yamada, H.; Doi, M.; Bando, H.; Yamaguchi, Y.; Nishigori, C.; Okamura, H. Molecular clocks in mouse skin. J. Investig. Dermatol. 2009, 129, 1225–1231.
- Sandu, C.; Dumas, M.; Malan, A.; Sambakhe, D.; Marteau, C.; Nizard, C.; Schnebert, S.; Perrier, E.; Challet, E.; Pevet, P.; et al. Human skin keratinocytes, melanocytes, and fibroblasts contain distinct circadian clock machineries. Cell Mol. Life Sci. 2012, 69, 3329–3339.
- Plikus, M.V.; Andersen, B. Skin as a window to body-clock time. Proc. Natl. Acad. Sci. USA 2018, 115, 12095–12097.
- Upton, B.A.; Diaz, N.M.; Gordon, S.A.; Van Gelder, R.N.; Buhr, E.D.; Lang, R.A. Evolutionary Constraint on Visual and Nonvisual Mammalian Opsins. J. Biol. Rhythm. 2021, 36, 109–126.
- Buhr, E.D.; Vemaraju, S.; Diaz, N.; Lang, R.A.; Van Gelder, R.N. Neuropsin (OPN5) Mediates Local Light-Dependent Induction of Circadian Clock Genes and Circadian Photoentrainment in Exposed Murine Skin. Curr. Biol. 2019, 29, 3478–3487.e4.
- de Assis, L.V.; Moraes, M.N.; da Silveira Cruz-Machado, S.; Castrucci, A.M. The effect of white light on normal and malignant murine melanocytes: A link between opsins, clock genes, and melanogenesis. Biochim. Biophys. Acta 2016, 1863, 1119–1133.
- de Assis, L.V.M.; Mendes, D.; Silva, M.M.; Kinker, G.S.; Pereira-Lima, I.; Moraes, M.N.; Menck, C.F.M.; Castrucci, A.M.L. Melanopsin mediates UVA-dependent modulation of proliferation, pigmentation, apoptosis, and molecular clock in normal and malignant melanocytes. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118789.
- Moraes, M.N.; de Assis, L.V.M.; Magalhaes-Marques, K.K.; Poletini, M.O.; de Lima, L.; Castrucci, A.M.L. Melanopsin, a Canonical Light Receptor, Mediates Thermal Activation of Clock Genes. Sci. Rep. 2017, 7, 13977.
- Solessio, E.; Engbretson, G.A. Antagonistic chromatic mechanisms in photoreceptors of the parietal eye of lizards. Nature 1993, 364, 442–445.
- Okano, T.; Yoshizawa, T.; Fukada, Y. Pinopsin is a chicken pineal photoreceptive molecule. Nature 1994, 372, 94–97.
- Zhao, X.; Haeseleer, F.; Fariss, R.N.; Huang, J.; Baehr, W.; Milam, A.H.; Palczewski, K. Molecular cloning and localization of rhodopsin kinase in the mammalian pineal. Vis. Neurosci. 1997, 14, 225–232.
- Czeisler, C.A.; Shanahan, T.L.; Klerman, E.B.; Martens, H.; Brotman, D.J.; Emens, J.S.; Klein, T.; Rizzo, J.F., 3rd. Suppression of melatonin secretion in some blind patients by exposure to bright light. N. Engl. J. Med. 1995, 332, 6–11.
- Campbell, S.S.; Murphy, P.J. Extraocular circadian phototransduction in humans. Science 1998, 279, 396–399.
- Busse, D.; Kudella, P.; Gruning, N.M.; Gisselmann, G.; Stander, S.; Luger, T.; Jacobsen, F.; Steinstrasser, L.; Paus, R.; Gkogkolou, P.; et al. A synthetic sandalwood odorant induces wound-healing processes in human keratinocytes via the olfactory receptor OR2AT4. J. Investig. Dermatol. 2014, 134, 2823–2832.
- Pavan, B.; Dalpiaz, A. Odorants could elicit repair processes in melanized neuronal and skin cells. Neural Regen Res. 2017, 12, 1401–1404.
- Spehr, M.; Gisselmann, G.; Poplawski, A.; Riffell, J.A.; Wetzel, C.H.; Zimmer, R.K.; Hatt, H. Identification of a testicular odorant receptor mediating human sperm chemotaxis. Science 2003, 299, 2054–2058.
- Griffin, C.A.; Kafadar, K.A.; Pavlath, G.K. MOR23 promotes muscle regeneration and regulates cell adhesion and migration. Dev. Cell 2009, 17, 649–661.
- Weber, L.; Al-Refae, K.; Ebbert, J.; Jagers, P.; Altmuller, J.; Becker, C.; Hahn, S.; Gisselmann, G.; Hatt, H. Activation of odorant receptor in colorectal cancer cells leads to inhibition of cell proliferation and apoptosis. PLoS ONE 2017, 12, e0172491.
- Sondersorg, A.C.; Busse, D.; Kyereme, J.; Rothermel, M.; Neufang, G.; Gisselmann, G.; Hatt, H.; Conrad, H. Chemosensory information processing between keratinocytes and trigeminal neurons. J. Biol. Chem. 2014, 289, 17529–17540.
- Tsai, T.; Veitinger, S.; Peek, I.; Busse, D.; Eckardt, J.; Vladimirova, D.; Jovancevic, N.; Wojcik, S.; Gisselmann, G.; Altmuller, J.; et al. Two olfactory receptors-OR2A4/7 and OR51B5-differentially affect epidermal proliferation and differentiation. Exp. Dermatol. 2017, 26, 58–65.
- Gelis, L.; Jovancevic, N.; Veitinger, S.; Mandal, B.; Arndt, H.D.; Neuhaus, E.M.; Hatt, H. Functional Characterization of the Odorant Receptor 51E2 in Human Melanocytes. J. Biol. Chem. 2016, 291, 17772–17786.
- Wojcik, S.; Weidinger, D.; Stander, S.; Luger, T.; Hatt, H.; Jovancevic, N. Functional characterization of the extranasal OR2A4/7 expressed in human melanocytes. Exp. Dermatol. 2018, 27, 1216–1223.
- Adameyko, I.; Lallemend, F. Glial versus melanocyte cell fate choice: Schwann cell precursors as a cellular origin of melanocytes. Cell Mol. Life Sci. 2010, 67, 3037–3055.
- Lerner, M.R.; Reagan, J.; Gyorgyi, T.; Roby, A. Olfaction by melanophores: What does it mean? Proc. Natl. Acad. Sci. USA 1988, 85, 261–264.
- Karlsson, J.O.; Svensson, S.P.; Martensson, L.G.; Odman, S.; Elwing, H.; Lundstrom, K.I. Effects of odorants on pigment aggregation and cAMP in fish melanophores. Pigment. Cell Res. 1994, 7, 61–64.
- Gelis, L.; Jovancevic, N.; Bechara, F.G.; Neuhaus, E.M.; Hatt, H. Functional expression of olfactory receptors in human primary melanoma and melanoma metastasis. Exp. Dermatol. 2017, 26, 569–576.
- Ranzani, M.; Iyer, V.; Ibarra-Soria, X.; Del Castillo Velasco-Herrera, M.; Garnett, M.; Logan, D.; Adams, D.J. Revisiting olfactory receptors as putative drivers of cancer. Wellcome Open Res. 2017, 2, 9.
- Suzuki, T.; Tomita, Y. Recent advances in genetic analyses of oculocutaneous albinism types 2 and 4. J. Dermatol. Sci. 2008, 51, 1–9.
- Nichols, S.E., Jr.; Reams, W.M., Jr. The occurrence and morphogenesis of melanocytes in the connective tissues of the PET/MCV mouse strain. J. Embryol. Exp. Morphol. 1960, 8, 24–32.
- Mjaatvedt, C.H.; Kern, C.B.; Norris, R.A.; Fairey, S.; Cave, C.L. Normal distribution of melanocytes in the mouse heart. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2005, 285, 748–757.
- Brito, F.C.; Kos, L. Timeline and distribution of melanocyte precursors in the mouse heart. Pigment. Cell Melanoma Res. 2008, 21, 464–470.
- Yajima, I.; Larue, L. The location of heart melanocytes is specified and the level of pigmentation in the heart may correlate with coat color. Pigment. Cell Melanoma Res. 2008, 21, 471–476.
- Sanchez-Pina, J.; Lorenzale, M.; Fernandez, M.C.; Duran, A.C.; Sans-Coma, V.; Fernandez, B. Pigmentation of the aortic and pulmonary valves in C57BL/6J x Balb/cByJ hybrid mice of different coat colours. Anat. Histol. Embryol. 2019, 48, 429–436.
- Balani, K.; Brito, F.C.; Kos, L.; Agarwal, A. Melanocyte pigmentation stiffens murine cardiac tricuspid valve leaflet. J. R. Soc. Interface 2009, 6, 1097–1102.
- Carneiro, F.; Kruithof, B.P.; Balani, K.; Agarwal, A.; Gaussin, V.; Kos, L. Relationships between melanocytes, mechanical properties and extracellular matrix composition in mouse heart valves. J. Long Term Eff. Med. Implant. 2015, 25, 17–26.
- Levin, M.D.; Lu, M.M.; Petrenko, N.B.; Hawkins, B.J.; Gupta, T.H.; Lang, D.; Buckley, P.T.; Jochems, J.; Liu, F.; Spurney, C.F.; et al. Melanocyte-like cells in the heart and pulmonary veins contribute to atrial arrhythmia triggers. J. Clin. Investig. 2009, 119, 3420–3436.
- Hwang, H.; Liu, F.; Levin, M.D.; Patel, V.V. Isolating primary melanocyte-like cells from the mouse heart. J. Vis. Exp. 2014, 91, 4357.
- Hwang, H.; Liu, F.; Petrenko, N.B.; Huang, J.; Schillinger, K.J.; Patel, V.V. Cardiac melanocytes influence atrial reactive oxygen species involved with electrical and structural remodeling in mice. Physiol. Rep. 2015, 3, e12559.
- Tsai, W.C.; Chan, Y.H.; Hsueh, C.H.; Everett, T.H., IV; Chang, P.C.; Choi, E.K.; Olaopa, M.A.; Lin, S.F.; Shen, C.; Kudela, M.A.; et al. Small conductance calcium-activated potassium current and the mechanism of atrial arrhythmia in mice with dysfunctional melanocyte-like cells. Heart Rhythm. 2016, 13, 1527–1535.
- Tachibana, M. Sound needs sound melanocytes to be heard. Pigment. Cell Res. 1999, 12, 344–354.
- Zhang, W.; Dai, M.; Fridberger, A.; Hassan, A.; Degagne, J.; Neng, L.; Zhang, F.; He, W.; Ren, T.; Trune, D.; et al. Perivascular-resident macrophage-like melanocytes in the inner ear are essential for the integrity of the intrastrial fluid-blood barrier. Proc. Natl. Acad. Sci. USA 2012, 109, 10388–10393.
- Zhang, F.; Dai, M.; Neng, L.; Zhang, J.H.; Zhi, Z.; Fridberger, A.; Shi, X. Perivascular macrophage-like melanocyte responsiveness to acoustic trauma--a salient feature of strial barrier associated hearing loss. FASEB J. 2013, 27, 3730–3740.
- Huang, S.; Song, J.; He, C.; Cai, X.; Yuan, K.; Mei, L.; Feng, Y. Genetic insights, disease mechanisms, and biological therapeutics for Waardenburg syndrome. Gene Ther. 2021.
- Andrade, A.; Pithon, M. Alezzandrini syndrome: Report of a sixth clinical case. Dermatology 2011, 222, 8–9.
- O’Keefe, G.A.; Rao, N.A. Vogt-Koyanagi-Harada disease. Surv. Ophthalmol. 2017, 62, 1–25.
- Lee, T.L.; Lin, P.H.; Chen, P.L.; Hong, J.B.; Wu, C.C. Hereditary Hearing Impairment with Cutaneous Abnormalities. Genes 2020, 12, 43.
- Anbar, T.S.; El-Badry, M.M.; McGrath, J.A.; Abdel-Azim, E.S. Most individuals with either segmental or non-segmental vitiligo display evidence of bilateral cochlear dysfunction. Br. J. Dermatol. 2015, 172, 406–411.
- Ertugrul, G.; Ertugrul, S.; Soylemez, E. Investigation of hearing and outer hair cell function of the cochlea in patients with vitiligo. Dermatol. Ther. 2020, 33, e13724.
- Genedy, R.; Assal, S.; Gomaa, A.; Almakkawy, B.; Elariny, A. Ocular and auditory abnormalities in patients with vitiligo: A case-control study. Clin. Exp. Dermatol. 2021, 46, 1058–1066.
- Mujica-Mota, M.A.; Schermbrucker, J.; Daniel, S.J. Eye color as a risk factor for acquired sensorineural hearing loss: A review. Hear. Res. 2015, 320, 1–10.
- Wrzesniok, D.; Beberok, A.; Otreba, M.; Buszman, E. Gentamicin affects melanogenesis in normal human melanocytes. Cutan. Ocul. Toxicol. 2015, 34, 107–111.
- Wrzesniok, D.; Rok, J.; Beberok, A.; Rzepka, Z.; Respondek, M.; Pilawa, B.; Zdybel, M.; Delijewski, M.; Buszman, E. Kanamycin induces free radicals formation in melanocytes: An important factor for aminoglycosides ototoxicity. J. Cell Biochem. 2019, 120, 1165–1173.
- Murillo-Cuesta, S.; Contreras, J.; Zurita, E.; Cediel, R.; Cantero, M.; Varela-Nieto, I.; Montoliu, L. Melanin precursors prevent premature age-related and noise-induced hearing loss in albino mice. Pigment Cell Melanoma Res. 2010, 23, 72–83.
- Shibuya, H.; Watanabe, R.; Maeno, A.; Ichimura, K.; Tamura, M.; Wakana, S.; Shiroishi, T.; Ohba, K.; Takeda, K.; Tomita, H.; et al. Melanocytes contribute to the vasculature of the choroid. Genes Genet. Syst. 2018, 93, 51–58.
- Gudjohnsen, S.; Atacho, D.; Gesbert, F.; Raposo, G.; Hurbain, I.; Larue, L.; Steingrimsson, E.; Petersen, P. Meningeal Melanocytes in the Mouse: Distribution and Dependence on Mitf. Front. Neuroanat. 2015, 9, 149.
- Randhawa, M.; Huff, T.; Valencia, J.C.; Younossi, Z.; Chandhoke, V.; Hearing, V.J.; Baranova, A. Evidence for the ectopic synthesis of melanin in human adipose tissue. FASEB J. 2009, 23, 835–843.
- Page, S.; Chandhoke, V.; Baranova, A. Melanin and melanogenesis in adipose tissue: Possible mechanisms for abating oxidative stress and inflammation? Obes. Rev. 2011, 12, e21–e31.
- Jehs, T.; Faber, C.; Udsen, M.S.; Jager, M.J.; Clark, S.J.; Nissen, M.H. Induction of Chemokine Secretion and Monocyte Migration by Human Choroidal Melanocytes in Response to Proinflammatory Cytokines. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6568–6579.
- Nag, T.C. Ultrastructural changes in the melanocytes of aging human choroid. Micron 2015, 79, 16–23.
- Jang, Y.H.; Kim, S.L.; Lee, J.S.; Kwon, K.Y.; Lee, S.J.; Kim, D.W.; Lee, W.J. Possible existence of melanocytes or melanoblasts in human sebaceous glands. Ann. Dermatol. 2014, 26, 469–473.
- Vandamme, N.; Berx, G. From neural crest cells to melanocytes: Cellular plasticity during development and beyond. Cell Mol. Life Sci. 2019, 76, 1919–1934.
- Adameyko, I.; Lallemend, F.; Aquino, J.B.; Pereira, J.A.; Topilko, P.; Muller, T.; Fritz, N.; Beljajeva, A.; Mochii, M.; Liste, I.; et al. Schwann cell precursors from nerve innervation are a cellular origin of melanocytes in skin. Cell 2009, 139, 366–379.
- Tatarakis, D.; Cang, Z.; Wu, X.; Sharma, P.P.; Karikomi, M.; MacLean, A.L.; Nie, Q.; Schilling, T.F. Single-cell transcriptomic analysis of zebrafish cranial neural crest reveals spatiotemporal regulation of lineage decisions during development. Cell Rep. 2021, 37, 110140.
- Kaucka, M.; Szarowska, B.; Kavkova, M.; Kastriti, M.E.; Kameneva, P.; Schmidt, I.; Peskova, L.; Joven Araus, A.; Simon, A.; Kaiser, J.; et al. Nerve-associated Schwann cell precursors contribute extracutaneous melanocytes to the heart, inner ear, supraorbital locations and brain meninges. Cell Mol. Life Sci. 2021, 78, 6033–6049.
- Bonnamour, G.; Soret, R.; Pilon, N. Dhh-expressing Schwann cell precursors contribute to skin and cochlear melanocytes, but not to vestibular melanocytes. Pigment Cell Melanoma Res. 2021, 34, 648–654.