Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Khairunnuur Fairuz Azman and Version 2 by Amina Yu.
Brain-derived neurotrophic factor (BDNF) has been extensively investigated, and it has emerged as an important regulator of synaptic plasticity, neuronal survival, and differentiation. In addition, BDNF serves as a crucial molecular target for the development of drugs to treat neurological diseases. The synthesis and maturation of BDNF is a multistep process, starting with the formation of several precursor isoforms.
- BDNF
- Molecular Mechanisms
- Signaling Cascades
1. Introduction
Neurodegenerative diseases are characterized by the gradual loss of neuronal structure or function, which can lead to neuronal death. Neurodegenerative disease often results in progressive cognitive, functional, and behavioral changes, manifested as dysfunctional motor and cognitive impairment. The most common types of neurodegenerative disease, such as Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, and amyotrophic lateral sclerosis (ALS), have all been extensively studied. Neurodegenerative diseases represent a major socioeconomic burden worldwide. The risk of acquiring a neurodegenerative disease increases dramatically with age [1]. Thus, with an aging population, the number of people affected is set to increase even further, necessitating the development of therapeutic strategies capable of reversing or stopping the degenerative process.
Neurotrophins are regulatory factors that mediate the differentiation and survival of neurons [2]. Neurotrophins include nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT3), and neurotrophin-4/5 (NT4/5), which are all derived from a common ancestral gene and have similar sequences and structures. Among neurotrophins, Brain-derived neurotrophic factor (BDNF)BDNF has been extensively investigated, and it has emerged as an important regulator of synaptic plasticity, neuronal survival, and differentiation. In addition, BDNF serves as a crucial molecular target for the development of drugs to treat neurological diseases. The first evidence of BDNF’s role in the etiology of neurodegenerative illnesses was discovered in the early 1990s. Since then, research on BDNF in neurodegenerative diseases has grown tremendously. In this review, we present recent updates on the role of BDNF and its downstream signaling pathways in neurodegenerative diseases, including Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, and ALS. In addition, the therapeutic potential of BDNF in the treatment of neurodegenerative diseases will be reviewed. Scopus database was searched for articles published between 2010 and 2022. The search terms included: “BDNF”, “neurodegenerative diseases”, “Alzheimer’s disease”, “Parkinson’s disease”, “Huntington’s disease”, and “amyotrophic lateral sclerosis”. Full articles were obtained and the references were reviewed for additional information when necessary.
2. BDNF Molecular Mechanisms and Signaling Cascades
The synthesis and maturation of BDNF is a multistep process, starting with the formation of several precursor isoforms. The precursor form of BDNF protein, preproBDNF, is synthesized in the endoplasmic reticulum (Figure 1) [3]. PreproBDNF is then converted into a precursor proneurotrophin isoform of BDNF (proBDNF) by the removal of the signal peptide. The proBDNF is made up of 129 amino acids with an N-terminal pro-domain and 118 amino acids, with a mature domain at the C-terminus [4]. The mature isoform of the BDNF (mBDNF) is produced by additional cleavage of the proBDNF. ProBDNF can be cleaved intracellularly by endoproteases from the subtilisin-kexin family, such as furin, or by convertases in intracellular vesicles [5]. Plasmin and matrix metalloproteases 2 and 9 (MMP2 and MMP9) are required for the extracellular cleavage of proBDNF [6]. The physiological activity of mBDNF and proBDNF is enabled by their secretion into the extracellular space. BDNF secretion can be either constitutive or activity-dependent, depending on the cell type [4]. In neuronal cells, both proBDNF and mBDNF are released upon cellular membrane depolarization [7]. The ratio of proBDNF to mBDNF varies, depending on the particular stages and regions of brain development. ProBDNF is higher in the early postnatal period, when it is important for the development of brain function, while mBDNF is higher in adulthood for brain function, such as neuroprotection and synaptic plasticity [8].
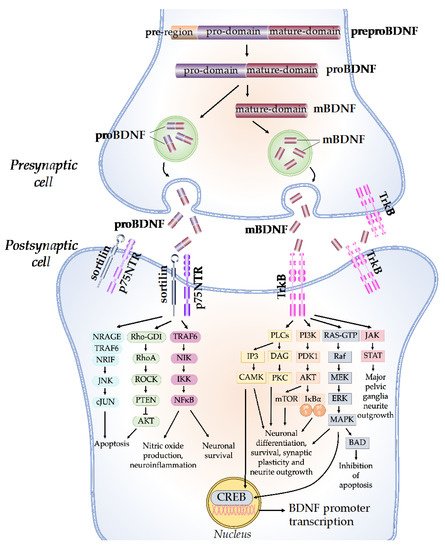
Figure 1. BDNF molecular mechanisms and signaling cascades. The BDNF protein is synthesized as a precursor form, preproBDNF. PreproBDNF is then converted into proBDNF, which consists of a pro-domain and a mature domain. The proBDNF is further cleaved to generate the mature isoform, mBDNF. ProBDNF and mBDNF are exocytosed into the extracellular space. The pro-domain of the proBDNF binds to the sortilin receptor, while the mature domain binds to the p75NTR receptor and activates the JNK/cJUN, PI3K/AKT, and TRAF6/NF-kB signaling pathways, which determine neuronal fate. The mBDNF binds to the TrkB receptor and activates PLCs, PI3K, MAPK, and JAK/STAT signaling cascades, which promote CREB translation, neuronal survival, and synaptic plasticity.
The network of BDNF/tyrosine receptor kinase B (TrkB) and BDNF/ p75 pan-neurotrophin receptor (p75NTR) signaling pathways was first graphically mapped by Sandhya et al. in 2013. Briefly, the mature domain of proBDNF interacts preferentially with the p75NTR, while the pro-domain interacts with the sortilin receptor or other vacuolar protein-sorting 10 protein (Vps10p) [9]. The binding of proBDNF to its specific receptor activates signaling pathways that determine the fate of neurons and synapses. The proBDNF/p75NTR/sortilin binding complex can cause the activation of the c-Jun amino-terminal kinase (JNK) pathway, leading to dendritic spine loss, caspase release, and neuronal apoptosis [10]. The activation of JNK requires neurotrophin receptor-interacting MAGE homolog (NRAGE), neurotrophin receptor-interacting factor (NRIF), and tumor necrosis factor receptor-associated factor 6 (TRAF6). The binding of proBDNF to p75NTR can also cause the activation of the RhoA/Rho-associated kinase (ROCK) signaling pathway [11]. ROCK then activates the phosphatase and tension homolog (PTEN), which blocks the phosphoinositide 3-kinases-protein kinase B (PI3K/AKT) signaling necessary for TrkB-induced potentiation, thus eliciting apoptosis [12]. In addition, the TRAF6 signaling pathway is initiated, which leads to nuclear factor kappa B (NF-kB) activation. The activation of NF-kB can either promotes neuronal survival or nitric oxide production and neuroinflammation through multiple reactions [13].
On the other hand, mBDNF binds with the TrkB receptor, which has a high affinity. Upon BDNF binding, TrkB dimerizes, intracellular tyrosine residues are autophosphorylated, and several enzymes are activated, including phospholipase C (PLC), PI3K, guanosine triphosphate hydrolases (GTP), and Janus kinase (JAK) [14]. Through the activation of calcium-calmodulin-dependent protein kinase (CAMK) and protein kinase C (PKC), the PLC-dependent pathway leads to calcium-dependent signaling steps and the release of calcium ions from intracellular calcium storage, resulting in increased synaptic plasticity [15]. The PI3K/AKT-related pathway modulates N-methyl-D-aspartate receptor (NMDAR)-dependent synaptic plasticity and exerts pro-survival and antiapoptotic activity [16]. The PI3K/Akt/mTOR cascade promotes dendritic growth and branching by regulating the synthesis of protein and the development of the cytoskeleton [17]. The mitogen-activated protein kinase (MAPK)/RAS-signaling cascade regulates the synthesis of protein during neuronal development and is also involved in the activation of extracellular-signal-regulated kinase 1/2 (ERK 1/2) and cAMP response element-binding protein (CREB) [18]. CREB activation leads to the initiation of transcription, the prolongation of synaptic potentiation, dendritic arborization, and neuroprotection [19]. The JAK/STAT pathway promotes major pelvic ganglia neurite outgrowths [20].
The specialized role of BDNF in the regulation of numerous physiological brain processes is determined by the interaction of BDNF isoforms with different types of receptors. For example, the activation of TrkB by BDNF is important in the late phase of long-term potentiation (L-LTP), which stimulates structural changes at the synapse [21]. On the other hand, P75NTR activation by proBDNF facilitates hippocampal long-term depression (LTD), which results in a decrease in synaptic strength [21]. The disruptions in BDNF production that result in signaling-cascade failure may be responsible for a range of neurological disorders. In addition, changes in BDNF levels and activities have been associated with a variety of neurodegenerative diseases, including Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, and ALS.